
草履蚧不同寄主和地理种群遗传分化的RAPD分析
2016-05-07 09:00刘军侠刘全超姜文虎宋晓英高宝嘉
生态学报 2016年2期
关键词:寄主植物
刘军侠,刘全超,姜文虎,宋晓英,高宝嘉,*
1河北农业大学林学院,保定 0710002河北省林木种质资源与森林保护重点实验室,保定 071000
草履蚧不同寄主和地理种群遗传分化的RAPD分析
刘军侠1,2,刘全超1,姜文虎1,宋晓英1,高宝嘉1,2,*
1河北农业大学林学院,保定071000
2河北省林木种质资源与森林保护重点实验室,保定071000
摘要:应用随机扩增多态性DNA(RAPD)技术对草履蚧保定、石家庄、邯郸16个不同寄主地理种群遗传多样性和种群分化进行研究,结果显示4个RAPD引物共扩增出41个多态性位点,多态位点比率为100%。遗传距离指数在0.701—0.4360,平均为0.2395。其中以邯郸枫杨和邯郸垂丝海棠为寄主的草履蚧种群遗传距离最小(0.0701);以石家庄紫叶李和邯郸木槿为寄主的种群遗传距离最大(0.4360)。遗传一致度系数在0.6466—0.9290。说明草履蚧不同种群遗传多样性丰富并存在遗传差异。聚类分析结果表明草履蚧种群遗传多样性同时受到寄主和地理因素的双重影响,且不同寄主草履蚧种群已产生明显的遗传分化。
关键词:草履蚧;寄主植物;种群分化; RAPD
刘军侠,刘全超,姜文虎,宋晓英,高宝嘉.草履蚧不同寄主和地理种群遗传分化的RAPD分析.生态学报,2016,36(2): 298-305.
Liu J X,Liu Q C,Jiang W H,Song X Y,Gao B J.RAPD analysis of genetic differentiation in the different hosts and geographic populations of Drosicha corpulenta.Acta Ecologica Sinica,2016,36(2): 298-305.
随着分子生物学发展,应用分子系统分析的方法研究昆虫种群分化和遗传多样性,从分子水平探讨寄主、环境因子等对昆虫适应进化的影响,以明确昆虫适应进化的分子机制。蚧虫作为一类营相对固定生活的刺吸类害虫,呈聚集型空间分布,扩散能力弱,其发生与寄主种群以及地理环境有着密切的关系[1-4],在长期的系统发育中形成了相对稳定的区系组成以适应不同环境,不同的寄主种群、生态地理种群间差异明显。高宝嘉等对皱大球蚧(Eulecanium kuwana i)和瘤坚大球蚧(E.gigan tean)地理种群的RAPD遗传分化的研究发现,已发生种内遗传变异,但尚未达到亚种水平[5]。Burban等通过对松干蚧的研究指出松干蚧种群变异与寄主植物的遗传结构有关[6]。本研究选取河北省不同草履蚧Drosicha corpulenta(kawana)种群,对其遗传分化进行RAPD分析,揭示寄主植物、地理环境等生态条件对蚧虫遗传分化的影响,为深入探讨植物昆虫协同进化关系和昆虫适应潜力提供依据。
1材料和方法
1.1供试昆虫
2011年4—5月于保定、石家庄、邯郸不同寄主上采集雌成虫(表1)。不同寄主相距50m以上,每种寄主东、南、西、北方向的上、中、下部枝条分别取样。将采集的蚧虫饥饿24h以上,使其消化道内的物质充分消化,以防外源DNA干扰实验结果;-20℃冰箱中保存。
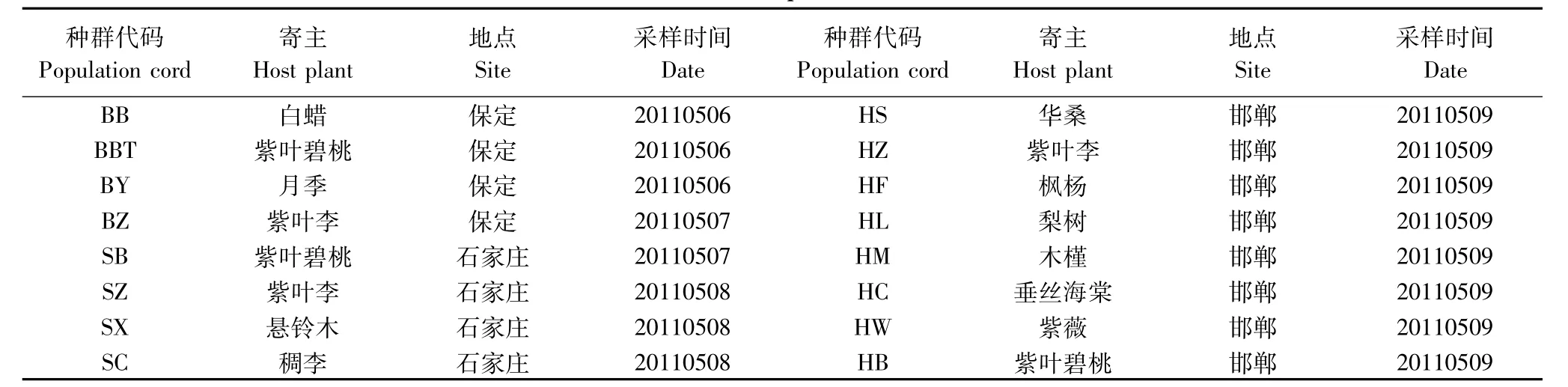
表1 供试草履蚧材料来源Table 1 Sources of D.corpulenta materials for trial
1.2主要试剂与仪器
本实验所用的TaqDNA聚合酶、dNTP、蛋白酶K、RAPD引物、DNA标记(DL2000)均由上海生物工程有限公司生产。
1.3研究方法
1.3.1提取DNA
参照Boyce等的方法[7],并做适当修改。取1只蚧虫于灭菌的研钵(每个种群取28个样本),加入液氮迅速研磨成粉末,放入1.5mL的离心管中,加入700μL预冷的2×CTAB(0.1mol/L Tris2-HCl,1.4mol/L NaCl,0.2mol/L EDTA,0.2%β巯基乙醇,0.05mol/L CT AB,pH8.3),加入蛋白酶K至终浓度为100μg/mL,65℃下水浴1h,用12000r/min离心5min,取上清液,加入等体积得酚∶氯仿∶异戊醇(25∶24∶1)轻轻混匀,反复抽提两次。再经等体积的异丙醇沉淀1h,4℃下10000r/min离心10min,去上清,70%的乙醇清洗1—2次,干燥后加入适量TE溶液溶解DNA,加入RNaesA至终浓度为2μg/L后,37℃下水浴1h,-20℃保存待用。
1.3.2 RAPD扩增及产物检测
经预试验筛选出4条可用引物,其引物名称及序列见表2。参照Williams的方法[8],反应体系25μL,其中模板50ng,10×Buffer2.5μL,Mg2+114—210 mmol/L(因座位而异),dNTP200μmol/L,引物(0.8μmol/L),Taq酶1.5U反应条件为: 94℃预变性1min; 92℃变性1 min,37℃退火1 min(因座位而异),72℃延伸2 min,45个循环; 72℃再延伸10min; 4℃终止反应。
扩增产物用1.5%琼脂糖凝胶(含1%核酸显色剂)分离,电泳缓冲液为0.5×TBE,上样量为10μL。电泳结束后,用凝胶成像仪拍照。
1.3.3数据分析
RAPD为显性标记,同一引物的扩增产物在琼脂糖凝胶电泳中迁移律相同的条带被认为是同源性的,属于同一位点的条带按清晰可见的强带或反复出现的弱带记为1,无带记为0,形成二元数据。应用Popstream32 和NTSYSpc2.1软件,计算种群遗传多样性参数并根据遗传相似系数进行UPGMA种群聚类分析。

表2 随机引物编号及序列Table 2 RAPD primer codes and their sequences
2结果与分析
2.1遗传多态位点分析
引物扩增结果见图1,不同种群遗传多样性统计见表3。供试引物对16个种群448个个体进行扩增,共产生41条清晰、稳定的片段,平均每条引物扩增10.25条片段,各片断分子量大小在200—2000bp之间,种群内和种群间并未产生各自特征性带谱。实验共检测出41个多态位点,多态位点比率为100%,说明草履蚧种群遗传多样性丰富。但不同种群的扩增产物多态性结果不同,各种群多态位点比率在31.71%—63.41%之间,其中邯郸紫薇寄主种群多态位点比率最低(31.71%),邯郸紫叶李寄主种群(63.41%)和邯郸桑树寄主种群(60.98%)多态位点最高,遗传多样性较其它种群丰富。
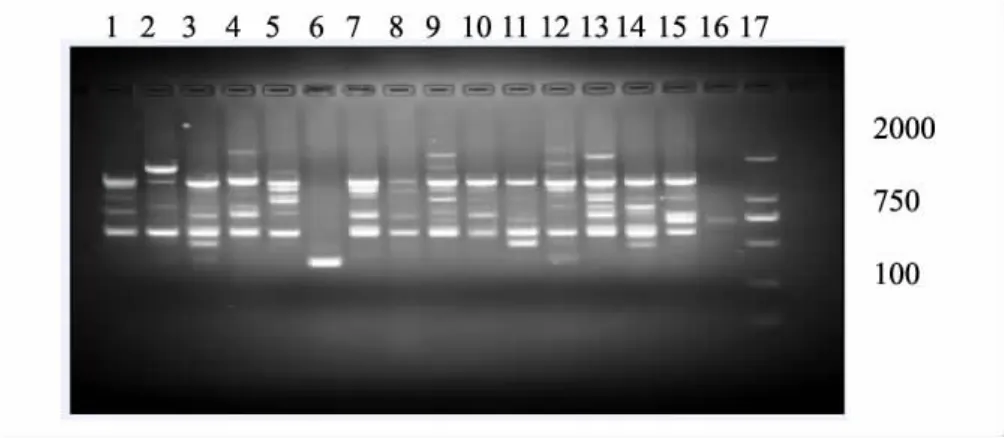
图1 引物H9对16个草履蚧种群的RAPD-PCR结果Fig.1 RAPD-PCR results of primer H9 for 16 populations of D.corpulenta1—4: HL、HF、HZ、HS; 5—8: HB、HW、HC、HM; 9—12: SC、SX、SZ、SB; 13—16: BY; BB; BZ; BBT; 17: DL2000
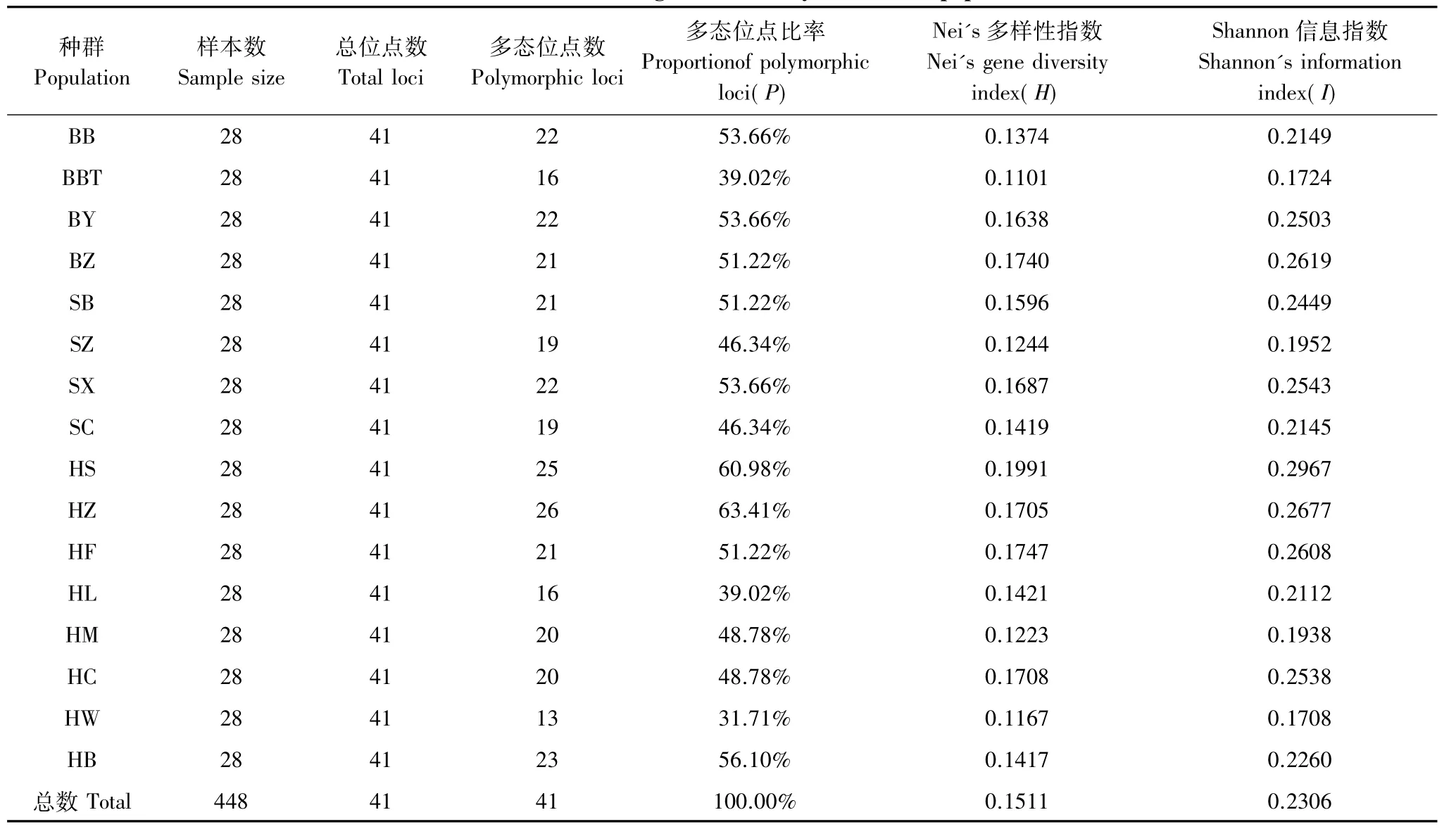
表3 不同种群遗传多样性统计Table 3 Statistics on the genetic diversity of different populations
2.2遗传多样性分析
Nei指数估计的不同种群间遗传距离和遗传一致度结果见表4。由表4可知,16个种群Shannon信息指数的变化范围在0.1708—0.2967之间,平均值为0.2306。邯郸桑树种群的Shannon信息指数最高,邯郸紫薇种群最低。16个种群Nei's基因多样性指数变化范围在0.1101—0.1991之间,平均值为0.1511。邯郸桑树种群的基因多样性最高,保定紫叶碧桃种群最低,这与Shannon信息指数估计的遗传多样性稍有差异。说明同一物种不同种群遗传多样性存在差异。
2.3遗传相似度与遗传距离分析
由表4可知,16个草履蚧种群的遗传相似度在0.6466—0.9290之间,平均值为0.7873;遗传距离在0.0701—0.4360之间,平均值为0.2395,说明不同种群间遗传差异较大。其中以邯郸枫杨和以邯郸垂丝海棠为寄主的草履蚧种群最相似,遗传距离最小(0.0701);以石家庄紫叶李和以邯郸木槿为寄主的种群遗传距离最大(0.4360),说明二者的遗传差异最大。
紫叶李邯郸种群与石家庄种群遗传距离为0.3330,与保定种群遗传距离为0.3426;紫叶碧桃邯郸种群与石家庄种群遗传距离为0.3170,与保定种群遗传距离为0.3816;即同种寄主不同地理种群随地理距离的增加遗传距离增加。
2.4种群遗传相似度聚类分析
用NTSYSpc2.1对草履蚧种群的遗传相似系数进行UPGMA聚类分析,进一步分析种群间遗传关系(图2)。结果显示:同一地理寄主种群绝大多数个体都能聚为一类,说明同一种群中个体间的遗传特性相似程度高,遗传稳定性;草履蚧相同寄主种群首先聚类,遗传相似系数在0.67水平,说明寄主因素对草履蚧的种群分化产生明显的影响;保定、石家庄、邯郸的紫叶碧桃种群各自聚类,且遗传距离较远;同样以上3地的紫叶李种群也各自聚类,说明地理因素对种群的分化产生影响;草履蚧不同寄主的三大地理种群并未产生明显地理聚类,说明相对于地理因素,寄主因素对草履蚧种群分化的影响更加明显。同一地区不同寄主种群聚类见图3、图4、图5,可见同一地区同一寄主的个体聚为一类,而同一地区不同寄主间聚类规律不明显。
3结论与讨论
本文通过以上对草履蚧不同寄主和地理种群遗传分化的RAPD分析,表明: Shannon信息指数和Nei's基因多样性指数显示草履蚧不同地理种群以及不同寄主种群遗传多样性丰富程度不同,草履蚧不同寄主种群之间存在不同程度的遗传差异,并已产生种群分化。草履蚧相同寄主不同地理种群之间存在遗传差异,地理因素对草履蚧种群分化产生影响。聚类分析结果显示,草履蚧种群分化同时受到寄主因素和地理因素的双重影响,相对于地理因素,寄主对草履蚧种群分化的影响更加明显。
草履蚧寄主植物广泛,主动扩散距离有限,因此其发生与寄主植物关系十分密切。本研究表明草履蚧不同寄主种群之间已经产生了明显的种群分化,这与草履蚧寄主生态条件、环境胁迫有密切关系。寄主植物自身的物理、化学等因素可影响植食性昆虫的生物学习性和遗传特性,促进昆虫种群分化,而昆虫对寄主植物具有选择性,对于不同寄主植物喜好程度也不尽相同[9-10]。昆虫学家认为植食性昆虫可因嗜食的寄主不同而造成遗传隔离,由此形成不同的物种。Takada、Blackman、Weber等研究表明烟蚜在不同寄主植物上存在着种群分化[11-13]。本实验中寄主植物分类科属不尽相同,其生物学特性和化学成分各异,我们推测寄主植物种类不同是造成草履蚧种群分化的重要原因。
此外,大气污染以及昆虫群落结构[14-15]等环境胁迫均能影响寄主植物与昆虫的关系,本实验寄主植物除受到草履蚧危害外,同时受到其他不同种昆虫取食。不同寄主植物上昆虫的种类、数量以及植物所处环境的不同可能都对草履蚧种群分化期作用,下一步有待从多因素分析探讨草履蚧种群分化原因。
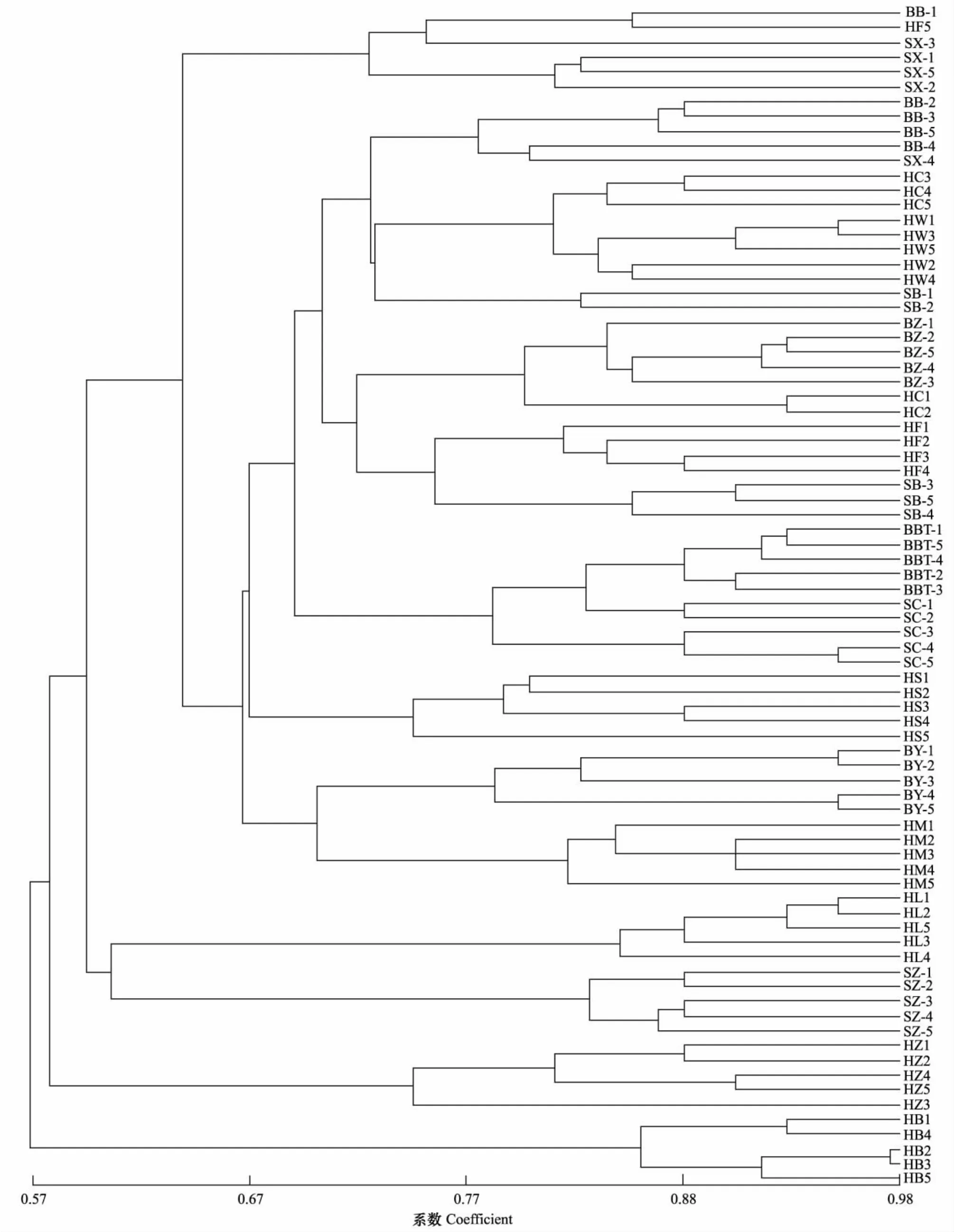
图2 16个草履蚧种群聚类分析图Fig.2 The UPGMA dendrogram cluster of 16 Drosicha corpul populations种群代号同表1(See table 1 for explanation of population code);为了方便比较查看,本图中每个种群采用5个样本的聚类分析;其聚类分析趋势与全部个体聚类图相一致,具有代表性
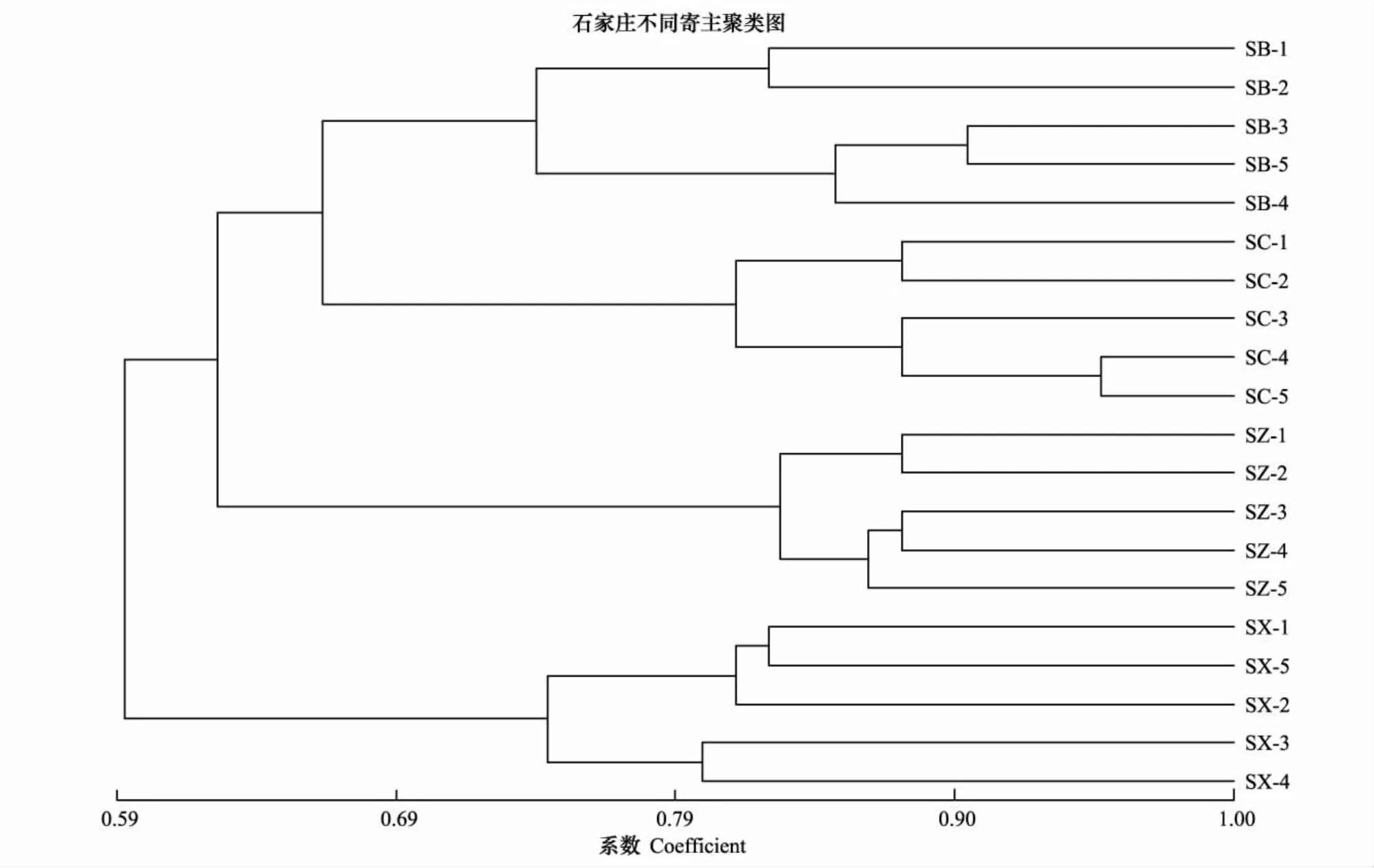
图3 石家庄草履蚧种群聚类分析图Fig.3 The UPGMA dendrogram cluster of Drosicha corpul populations of Shijiazhuang
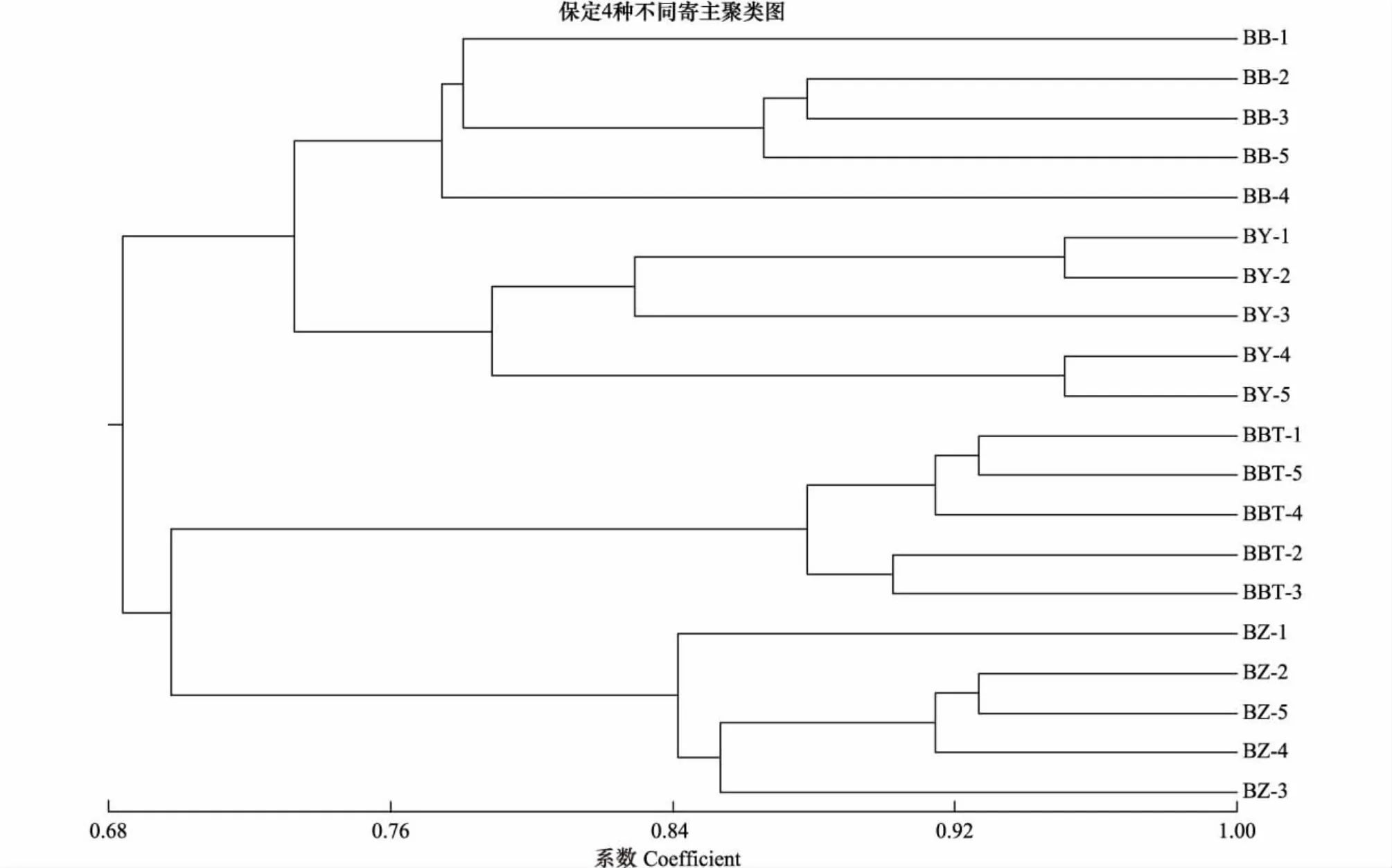
图4 保定草履蚧种群聚类分析图Fig.4 The UPGMA dendrogram cluster of Drosicha corpul populations of Baoding
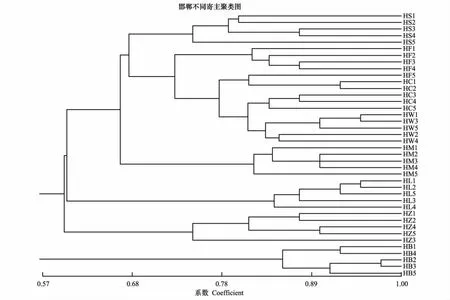
图5 邯郸草履蚧种群聚类分析图Fig.5 The UPGMA dendrogram cluster of Drosicha corpul populations of Handan
参考文献(References):
[1]梁隐泉,高宝嘉,甄志先,王江柱,刘志群,牛建忠.枣园昆虫群落及其与日本龟蜡蚧发生的关系.应用生态学,2006,17(3): 472-476.
[2]薛皎亮,谢映平,范三红,李艳芳.城市污染促进瘤坚大球蚧种群增长的营养学研究.林业科学.2004,40(1): 111-116.
[3]崔巍,高宝嘉.华北经济树种主要蚧虫及其防治.北京:林业出版社,1995.
[4]Beuning L L,Murphy P,Wu E,Batchelor T A,Morris B A M.Molecular-based approach to the differentiation of mealybug(Hemiptera: Pseudococcidae)species.Journal of Economic Entomology,1999,92(2): 463-472.
[5]高宝嘉,李梦,王春和,高立杰.皱大球蚧(Eulecanium kuwanai)和瘤坚大球蚧(Eulecanium gigantean)地理种群的RAPD遗传分化.生态学报,2007,27(3): 918-923.
[6]Burban C,Petit R J,Carcreff E,Jactel H.Rangewide variation of the maritime pine bast scale Matsucoccus feytaudi Duc.(Homoptera: Matsucoccidae)in relation to the genetic structure of its host.Molecular Ecology,1999,10(8): 1593-1602.
[7]Boyce T M,Zwick M E,Aquado C F.Mitochondrial DNA in the bark weevils: size,structure and heteroplasmy.Genetics,1989,123(4): 825-836.
[8]Williams J G,Kubelik A R,Livak K J,Rafalski J A,Tinqey S V.DNA polymorphisms amplified by arbitrary primers are useful as genetic markers.Nucleic Acids Research,1990,18(22): 6531-6535.
[9]Kelley S T,Farrell B D,Mitton J B.Effects of specialization on genetic differentiation in sister species of bark beetles.Heredity,2000,84(2): 218-227.
[10]Shivayogeshwara B,Mallikharjunaiah H,Krishnaprasad N K,Shetty M V N.Integrated management of spodoptera litura Fabricius(Noctuidae: Lepidoptera)in FCV tobacco crop.Tobacco Research,1991,17(2): 59-61.
[11]Takada H.Differences in adaptation to tobacco between host forms of Myzus persicae(Sulzer)(Homoptera: Aphididae).Japanese Journal of Applied Entomology and Zoology,1988,32(2): 115-119.
[12]Blackman R L.Morphological discrimination of a tobacco-feeding form from Myzus persicae(Sulzer)(Homoptera: Aphididae)and a key to new world Myzus(nectarosiphon)species.Bulletin of Entomological Research,1987,77: 713-730.
[13]Weber G.Genetic variability in host plant adaptation of the green peach aphid,Myzus persicae.Entomologia Experimentalis et Applicata,1985,38(1): 49-56.
[14]Nelson B W,John M.Lead residues in Eastern Tent caterpillars(Malacosoma americanum)and their host plant(Prunus serotina)close to a major highway.Environment Entomology,1980,9(1): 10-12.
[15]Alstad D N,Edmunds G F Jr,Weinstein L H.Effects of air pollutants on insect populations.Annual Review of Entomology,1982,27: 369-384.
RAPD analysis of genetic differentiation in the different hosts and geographic populations of Drosicha corpulenta
LIU Junxia1,2,LIU Quanchao1,JIANG Wenhu1,SONG Xiaoying1,GAO Baojia1,2,*
1 College of Forestry,Agricultural University of Hebei,Baoding 071000,China
2 Key Laboratory of Germplasm Resources of Forest Trees and Forest Protection College of Forest of Hebei,Baoding 071000,China
Abstract:The genetic diversity and population differentiation among geographic populations of 16 different Drosicha corpulenta host collected from Baoding,Shijiazhuang,and Handan in Hebei were analyzed by RAPD.The results showed that four RAPD primers were amplified out of 41 polymorphism loci,and that the percent of polymorphic loci was 100%.The genetic distance index of the populations ranged from 0.0701 to 0.4360 and the average value was 0.2395.The genetic distance index for D.corpulenta populations on Pterocarya stenoptera and Malus halliana(Voss.)Koehne in Handan was the smallest(0.0701).The genetic distance index for D.corpulenta populations on Prunus cerasifera in Shijiazhuang and Hibiscus syriacus in Handan was the largest of all,and the similarity coefficients ranged from 0.6466 to 0.9290.The genetic diversity of the different D.corpulenta populations was classified as abundant,and genetic differences were found.According to the results of cluster analysis,the genetic diversity of D.corpulenta populations was affected by both host and geographic location,and the different host populations of D.corpulenta showed obvious genetic differentiation.
Key Words:Drosicha corpulenta; host plant; population differentiation; RAPD
*通讯作者
Corresponding author.E-mail: baojiagao@ 163.com
收稿日期:2013-06-13;网络出版日期: 2015-06-10
基金项目:河北省自然基金(C2009000592)
DOI:10.5846/stxb201306131697
猜你喜欢
植物保护(2019年2期)2019-07-23
植物保护(2019年2期)2019-07-23
植物保护(2019年6期)2019-02-10
安徽农业科学(2018年16期)2018-05-14
安徽农学通报(2018年23期)2018-02-14
现代园艺(2017年1期)2017-07-14
江苏农业科学(2017年7期)2017-05-23
山东农业科学(2014年9期)2015-01-07
湖南中医药大学学报(2011年11期)2011-03-20
