
不同钙镁比处理对三种超富集植物生长及吸收重金属的影响
2016-04-21 08:11刘登彪郑俊宇蒋成爱
农业环境科学学报 2016年3期
赵 涛,刘登彪,2,郑俊宇,蒋成爱,3*
(1.华南农业大学资源环境学院,广州510642;2.深圳市铁汉生态环境股份有限公司,广东深圳518000;3.广东高校土壤环境与固体废物农业利用重点实验室,广州510642)
不同钙镁比处理对三种超富集植物生长及吸收重金属的影响
赵涛1,刘登彪1,2,郑俊宇1,蒋成爱1,3*
(1.华南农业大学资源环境学院,广州510642;2.深圳市铁汉生态环境股份有限公司,广东深圳518000;3.广东高校土壤环境与固体废物农业利用重点实验室,广州510642)
摘要:通过水培实验研究了不同钙镁比(Ca/Mg=0.2、0.5、1.0、2.0、4.0、6.0)对Zn/Cd超富集植物东南景天(Sedum alfredii)、Zn/Cd/Ni超富集植物遏蓝菜(Noccaca caerulescens)和Ni超富集植物庭荠属(Alyssum murale)在20 mg Zn·L(-1)、18 mg Ni·L(-1)、10 mg Pb·L(-1)和0.5 mg Cd·L(-1)胁迫下生长和吸收重金属的影响。研究结果显示:A.murale具有很强的将Ni从地下部转移至地上部的能力(转移系数TF达5.98~11.19)和较高的地上部Ca/Mg比(2.45~16.74);N.caerulescens对Mg转移能力(TF(Mg)为1.43~3.70)大于Ca(TF(Ca)为0.77~1.38);S.alfredii对Mg的转移能力与Ca相似。N.caerulescens在正常营养液(Ca/Mg=2)条件下获得最好的生长和地上部最大Zn/Cd/ Ni含量,增加Ca或Mg的供给都会产生抑制作用;A.murale在高Ca(Ca/Mg=6)供给时获得最大的生物量和Ni含量,增Ca有助于地上部Ni的累积,但增Mg降低了地上部Ni含量;增加Ca的供给会增加S.alfredii地上部和地下部的Zn含量,增加Ca或Mg的供给都会增加S.alfredii地下部Cd的含量,但S.alfredii在Ca/Mg=0.2时获得最大的生物量和最大的Zn和Cd含量。S.alfredii、A.murale 和N.caerulescens地上部分别在Ca/Mg比为6、0.2、1.0时获得最大的Pb含量,其他Ca/Mg比对三种超富集植物地上部吸收Pb均没有产生显著影响。Ca/Mg比对超富集植物富集重金属能力的影响因植物种类和元素而异。
关键词:钙镁比;超富集植物;锌;镍;镉;铅
赵涛,刘登彪,郑俊宇,等.不同钙镁比处理对三种超富集植物生长及吸收重金属的影响[J].农业环境科学学报, 2016, 35(3):440-448.
ZHAO Tao, LIU Deng-biao, ZHENG Jun-yu, et al. Effects of Ca/Mg ratios on growth and heavy metal uptake of three hyperaccumulators[J]. Journal of Agro-Environment Science, 2016, 35(3): 440-448.
重金属污染的植物修复技术是利用超富集植物对重金属的吸收、富集和转化能力来去除土壤中的重金属,具有经济、绿色环保等优点。国内外已发现的超富集植物绝大多数是在重金属胁迫环境下经过长期的适应基因变异而形成的生态耐受型,它们能够在高浓度重金属污染的土壤中生存,并将重金属转移富集在植物地上部,但它们存在生长缓慢、生育期推迟及生物量减少等问题,进而导致修复过程历时较长,制约着植物修复技术在实际生产中的应用[1-2]。通过农艺措施来优化调控这些植物的生长和重金属的吸收,是提高植物修复效率的重要途径之一。我们前期研究发现营养液中不同水平硼(B)和磷(P)的供给对不同超富集植物生长和吸收重金属产生了不同效应:除了高P对Alyssum murale(庭荠属)超积累重金属的促进作用外,不同浓度的B和P处理对Sedum alfredii(东南景天)、Noccaea caerulescens(遏蓝菜)和A.murale(庭荠属)超富集元素的累积都表现出低浓度促进和高浓度抑制的效应,但不同超富集植物生长和吸收重金属最佳的B浓度和P浓度均有不同[3-4]。
Ca和Mg都是植物必需的大量营养元素。Ca是植物细胞壁及细胞膜的结构成分,也是介导各种激素或环境条件引起的生理反应的第二信使,可与植物液泡中多种无机、有机阴离子结合,在植物生长发育及许多生理活动中起着重要作用[5]。Mg是叶绿素的成分,也是多种酶的活化剂,对植物光合作用有重要的作用,还对植物的活性氧代谢、酶活性、基因表达及其他离子的吸收等产生重要影响[6]。一些研究表明施加外源Ca或者Mg可以有效减少植物体内重金属的含量,并缓解重金属对植物的毒害效应[6-11]。Tian等[8]报道添加高浓度的外源Ca可以缓解超富集植物S.alfredii体内由Cd胁迫所诱导的脂质过氧化和细胞膜完整性被破坏等毒害效应,进而减弱Cd对其生长的抑制作用。汪骢跃等发现Ca2+对拟南芥幼苗Cd毒害具有缓解作用[9]。Juang等[6]通过15 d的水培实验研究了Mg对Cu在葡萄藤中累积和转运的影响,结果显示Mg2+浓度在0.15~2.01 mmol·L-1时能够有效降低植株对Cu的吸收及毒性。但在复合污染协同作用下,通过增加生长基质中Ca或Mg的浓度来调整Ca/Mg比对多种超富集植物吸收累积重金属的研究还未见报道。
不同植物对Ca或Mg吸收的偏好不同,大部分植物对Ca的需求大于Mg,但也有一些植物例外,如烤烟和生长在富Mg缺Ca蛇纹岩土壤上的Ni超富集植物[12,14]。本研究以正常营养液的Ca/Mg比为基础,通过增加Ca或Mg的浓度调节营养液Ca/Mg比,采用水培试验探讨在超富集重金属Zn、Cd、Ni及非富集重金属Pb复合污染胁迫下,不同Ca/Mg比处理对多种超富集植物的生长及吸收重金属的影响,为利用生长基质Ca/Mg比来优化调控超富集植物对重金属的提取效率提供应用基础。
1 材料与方法
1.1实验材料
本研究选取的三种超富集植物分别是:2002年在浙江衢州古老的铅锌矿区发现的一种本土Zn/Cd超富集植物矿山生态型东南景天(S.alfredii)[15]、生长在法国南部蛇纹岩土壤能超富集Zn/Ni/Cd等多种重金属的遏蓝菜(N.caerulescens)[4]和来自于希腊的十字花科庭荠属Ni的超富集植物(A.murale)[4,14]。S.alfredii的种苗取自浙江省铅锌矿山上,于2013年9月移至温室繁殖,用石英砂和完全培养液预培育2个月后,开始盆栽试验;N.caerulescens和A.murale的种子于2011年分别取自法国和希腊。
1.2实验设计和水培实验
水培实验在华南农业大学的网室进行。营养液采用华南农业大学叶菜配方[16],其组成为:Ca(NO3)2· 4H2O 472 mg·L-1,KNO3202 mg·L-1,NH4NO380 mg·L-1,KH2PO4100 mg·L-1,K2SO4174 mg·L-1,MgSO4·7H2O 246 mg·L-1,H3BO32.86 mg·L-1,MnSO4·4H2O 2.13 mg· L-1,ZnSO4·7H2O 0.22 mg·L-1,CuSO4·5H2O 0.08 mg·L-1,(NH4)6Mo7O24·4H2O 0.02 mg·L-1,FeSO4·7H2O 27.8 mg· L-1,EDTA-2Na 37.2 mg·L-1。用石英砂和稀释的营养液育种后,选取大小、长短一致的各种植物苗,每盆装营养液1.2 L种植4株。用完全营养液培养一个月后,将营养液中的Zn去掉,按表1进行不同Ca/Mg比处理和添加重金属,其中Ca/Mg比为2的是正常营养液,即为对照组,每个处理4个重复。
预培养和处理期间均保持24 h连续通气,每3 d更换一次营养液,处理40 d后,按地上部和地下部(根部)分开收获植物,用自来水清洗地上部并反复冲洗地下部,再用0.02 mol·L-1EDTA-2Na交换15 min,去除根系表面吸附的Zn2+、Ni2+、Pb2+和Cd2+,最后用去离子水冲洗干净,用吸水纸将植株表水吸干。然后将各植物样品置于105℃烘箱中杀青30 min,于75℃连续烘干48 h至恒重,冷却后称量植物体地上部和地下部的重量,得干生物量。研磨后供元素分析用。
1.3样品的分析测定
研磨后的植物样品,炭化后在马福炉550℃干灰化8 h,用1:1(V:V)HCl溶解、过滤、定容,用原子吸收光度法测定Ca、Mg、Zn、Cd、Ni和Pb含量。在植物样品的测试过程中每批次样品均同时做三个全程序的空白样,并随机抽取25%的样品做平行实验,同时采用国家标准参比物质(植物:GBW-08501)做质量控制。植株对金属的转运系数=地上部金属含量/地下部金属含量。
数据采用SPSS 18.0软件进行数据的统计分析,设定5%的显著性水平,运用单因素随机排列方差分析,并用Duncan氏法进行平均值间的多重比较分析。
2 结果与分析
2.1不同Ca/Mg比对超富集植物生长及生物量的影响
不同Ca/Mg比对三种植物的生长产生了不同的效应,N.caerulescens、S.alfredii和A.murale分别在Ca/ Mg比为2、0.2和6时生长相对较好,并获得最大的生物量(见表2)。增Mg显著地增加了S.alfredii生物量,Ca/Mg比为0.2时其地上部和地下部的生物量较Ca/Mg为2(正常营养液的配比)时分别提高了68.2% 和68.6%;增Ca促进了A.murale的生长,Ca/Mg比为0.2时长势比较差,根系分生少,Ca/Mg比为6的生物量远远大于Ca/Mg比为0.2的。以正常营养液的Ca/ Mg比为基础,增Ca或增Mg均抑制了N.caerulescens的生长,显著降低了其生物量。
2.2不同Ca/Mg比对超富集植物Ca、Mg含量的影响
不同Ca/Mg比处理下三种植物Ca和Mg含量分别如图1和图2所示。

表1 不同Ca/Mg比处理实验设计Table 1 Experimental design for different calcium to magnesium ratios
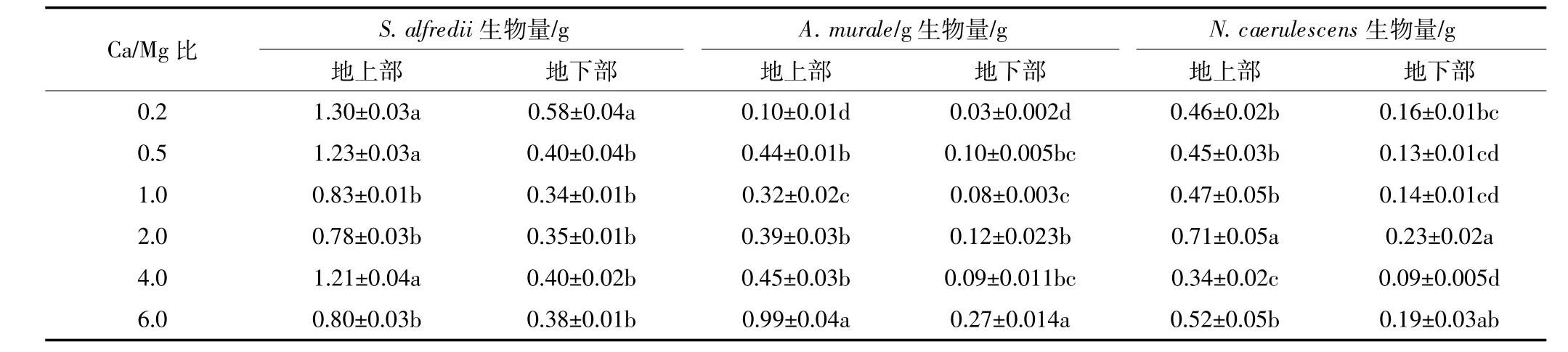
表2 不同Ca/Mg比对三种超富集植物生物量的影响Table 2 Biomass of three hyperaccumulators at different calcium to magnesium ratios
三种超富集植物地上部Ca含量随着营养液中Ca浓度的增加而升高,在Ca/Mg为6时获得最大含量,Mg浓度变化对N.caerulescens地上部Ca含量影响最明显。S.alfredii、A.murale和N.caerulescens地上部Ca浓度范围分别为10.1~16.6、27.4~42.7、6.2~17.4 g·kg-1,最大含量分别是各自最小含量的1.6、1.6、2.8倍;S.alfredii和N.caerulescens地上部Ca含量相接近,而A.murale地上部Ca含量显著高于二者。S.alfredii、A.murale和N.caerulescens根部Ca含量分别是8.1~12.8、3.5~5.1、2.9~3.7 g·kg-1,S.alfredii地下部Ca含量显著高于A.murale和N.caerulescens,后两种植物的地上部Ca含量又显著高于地下部,对Ca的转运系数分别达5.4~11.2和1.93~4.79。Ca/Mg比为0.2时显著地抑制了地上部Ca的浓度和转运;大多数Ca/Mg比则没有对三种植物根系含量产生显著影响。
三种超富集植物地上部和地下部Mg含量随着营养液中Mg浓度的增加有不同程度的增加,大部分都在Ca/Mg比为0.2时获得最大含量,而增加Ca的供给没有对植物地上部和地下部的Mg含量产生显著影响。S.alfredii、A.murale和N.caerulescens地上部Mg含量范围分别为2.6~3.3、2.8~11.2、2.3~4.8 g·kg-1,其最大含量分别是其最小含量的1.3、4.0、2.1倍;三种植物根部Mg含量分别是2.4~3.8、1.4~3.4、0.9~1.6 g·kg-1。A.murale地上部在最高Mg供给时获得远高于其他处理的Mg含量,A .murale和N. caerulescens地上部Mg含量显著高于其地下部。S. alfredii、A.murale和N.caerulescens地上部Ca/Mg比分别为3.47~6.14、2.45~15.53、1.48~5.97。
2.3不同Ca/Mg比对超富集植物植物提取Zn、Cd、Ni 和Pb的影响
不同Ca/Mg比处理下,三种超富集植物Zn、Cd、Ni和Pb的含量分别如图3、图4、图5和图6所示。

图1 不同Ca/Mg比对三种超富集植物Ca含量的影响Figure 1 Concentrations of Ca in shoots and roots of three hyperaccumulators at different calcium to magnesium ratios
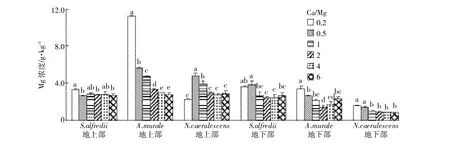
图2 不同Ca/Mg比对三种超富集植物Mg含量的影响Figure 2 Concentrations of Mg in shoots and roots of three hyperaccumulators at different calcium to magnesium ratios
S.alfredii、A.murale和N.caerulescens地上部Zn含量范围分别为3 403.3~6 146.7、2 021.0~2 971.4、2 007.1~2 777.6 mg·kg-1,地下部含量分别是7 573.2~12 448.0、8 502.2~13 838.4、1 599.2~4 062.3 mg·kg-1。随着Ca/Mg比从0.5增加到6,S.alfredii地上部Zn含量不断增加,在Ca/Mg比为6时获得最大Zn含量。在正常营养液的Ca/Mg比的基础上,增加Ca的供给不同程度地降低A.murale和N.caerulescens对Zn的吸收,但增加Mg的供给没有对A.murale地上部Zn含量产生明显影响,当Ca/Mg比减少到0.2时显著降低了N.caerulescens地上部Zn含量,A.murale和N. caerulescens在Ca/Mg比为2时获得最大Zn含量。S. alfredii地上部Zn含量显著高于A.murale和N. caerulescens,A.murale和N.caerulescens地上部Zn含量相近。增加营养液中Mg或Ca的供给不同程度地增加了三种植物地下部对Zn的吸收,但过高的供给又会抑制其地下部的吸收,三种植物起促进作用的Ca/Mg比范围存在差异。本试验条件下,Zn的超富集植物S.alfredii和N.caerulescens根系Zn含量均高于其地上部(见表3),造成这种异常结果的原因有待下一步试验来明确。

图3 不同Ca/Mg比对三种超富集植物Zn含量的影响Figure 3 Concentrations of Zn in shoots and roots in three hyperaccumulators at different calcium to magnesium ratios
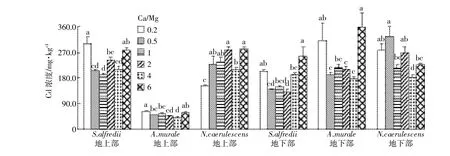
图4 不同Ca/Mg比对三种超富集植物Cd含量的影响Figure 4 Concentrations of Cd in shoots and roots of three hyperaccumulators at different calcium to magnesium ratios

图5 不同Ca/Mg比对三种超富集植物Ni含量的影响Figure 5 Concentrations of Ni in shoots and roots of three hyperaccumulators at different calcium to magnesium ratios
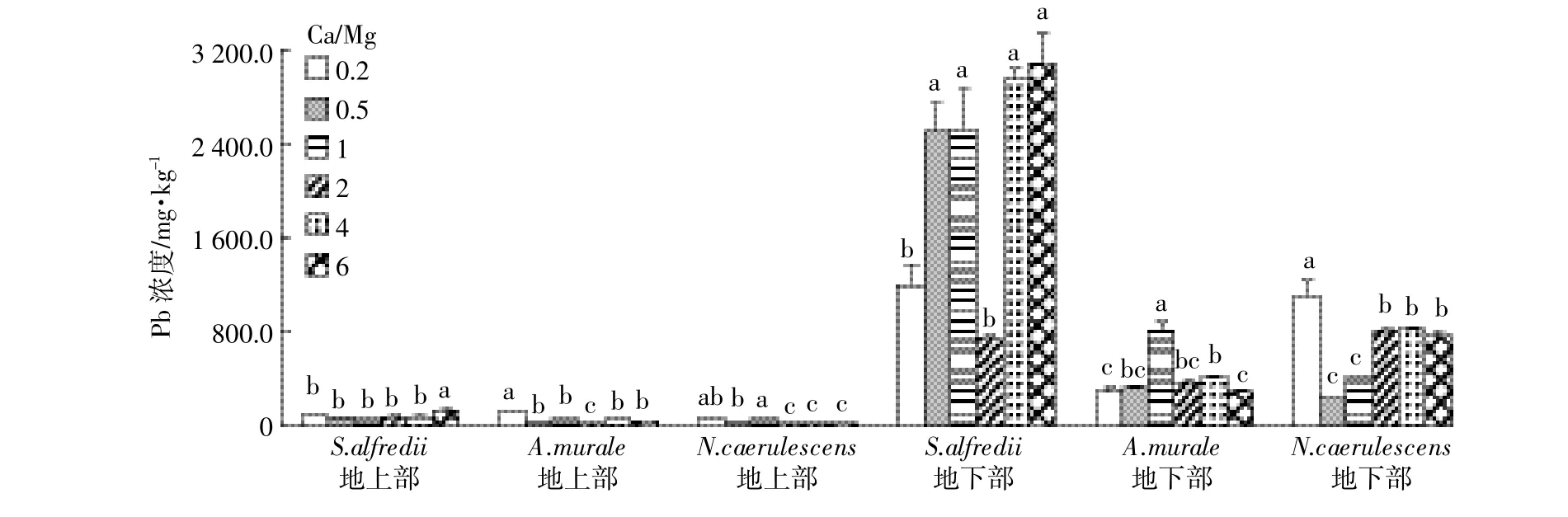
图6 不同Ca/Mg比对三种超富集植物Pb含量的影响Figure 6 Concentrations of Pb in shoots and roots of three hyperaccumulators at different calcium to magnesium ratios
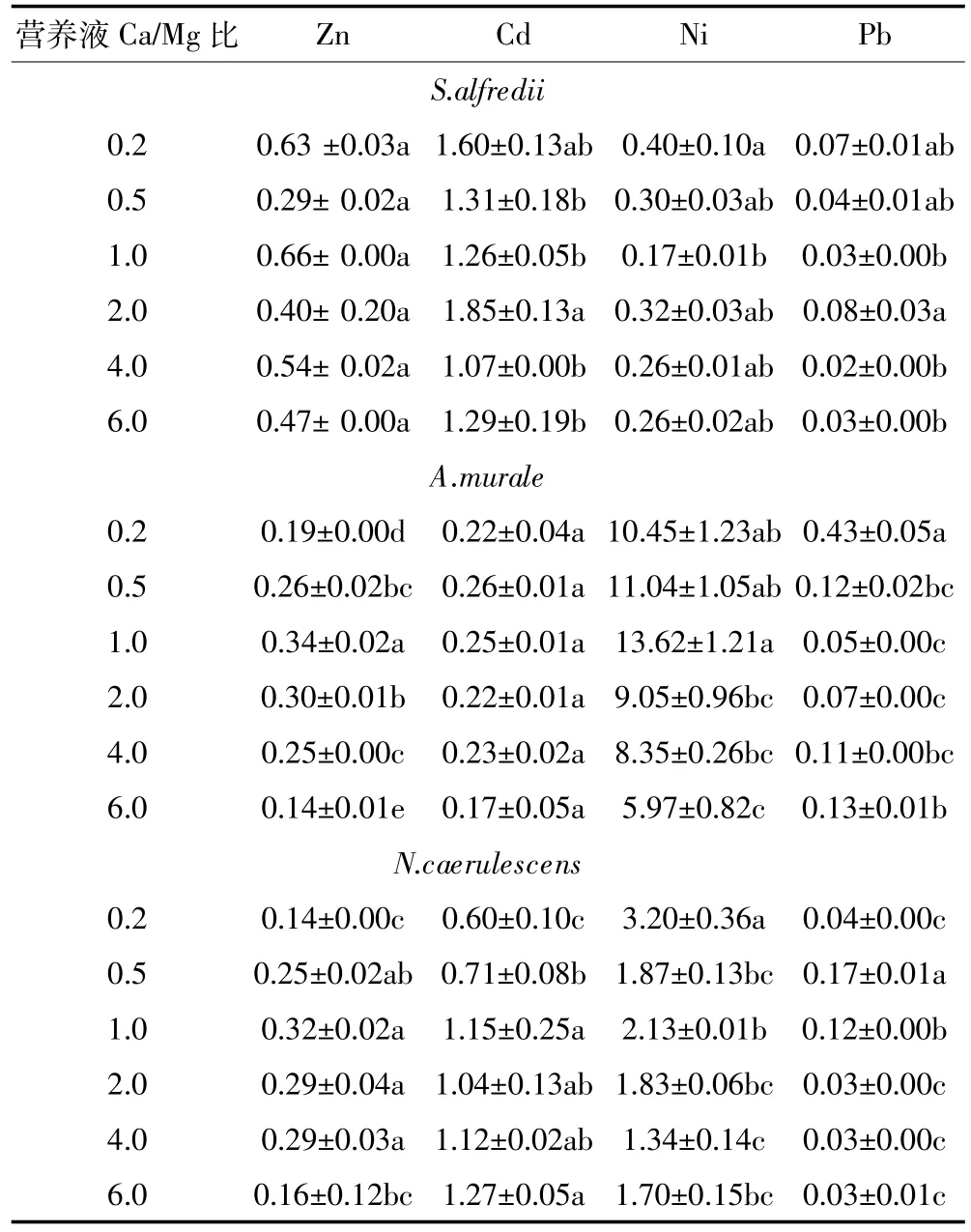
表3 不同Ca/Mg比处理下三种超富集植物重金属转运系数Table 3 Transfer coefficients of metals in three hyperaccumulators at different calcium to magnesium ratios
S.alfredii、A.murale和N.caerulescens地上部Cd浓度范围分别为191.6~300.8、40.2~61.9、153.5~281.2 mg·kg-1,地下部含量分别是140.4~256.1、175.8~358.6、182.7~327.3 mg·kg-1;三种超富集植物中A.murale不是Cd的超富集植物,其地上部Cd含量没有超过100 mg·kg-1,显著低于其地下部的含量。相似的趋势是,在正常营养液Ca/Mg比的基础上,增加Ca或Mg的供给会先降低三种植物地上部及地下部的Cd含量,再增加供给时植物体内Cd含量也会增加,但出现促进作用所需的Ca或Mg供给浓度因植物种类和器官不同而存在差异。在正常营养液的Ca/Mg比为2时,N. caerulescens体内Cd含量最高,但S.alfredii和A. murale则都在Ca/Mg比为0.5和6时获得相对较大的Cd含量。
S.alfredii、A.murale和N.caerulescens地上部Ni含量范围分别为148.3~196.9、1 703.1~2 659.8、1 104.2~1 537.0 mg·kg-1,根系含量分别是497.7~886.4、149.8~434.6、357.9~1 040.9 mg·kg-1。本试验供试的三种超富集植物中,A.murale和N.caerulescens是Ni的超富集植物,两者的地上部Ni含量均大于地下部,特别是A.murale,其转运系数高达6.1~14.48,三种植物地上部Ni的含量为A.murale>N.caerulescens≫S.alfredii。A.murale和N.caerulescens地上部分别在Ca/ Mg比为6和2(正常营养液的配比)时获得相对较大的Ni含量,增加营养液中Mg的供给能够降低二者地上部Ni含量,增加Ca的供给不会显著促进A.murale地上部Ni积累,并且会抑制N.caerulescens地上部Ni积累,营养液中Ca或Mg的浓度变化对这两种超富集植物根系Ni的吸收也有类似的效应。营养液中Ca或Mg浓度变化没有对S.alfredii地上部Ni含量产生显著或有规律的影响,但营养液中Mg浓度增加在一定范围内可以显著促进其根系对Ni的吸收。
S.alfredii、A.murale和N.caerulescens地上部Pb含量范围分别为57.0~126.3、25.1~121.7、24.7~46.5 mg·kg-1,地下部含量分别是742.8~3083.1、285.9~805.8、227.5~1 105.1 mg·kg-1。本试验中三种超富集植物都不是Pb的超富集植物,其地下部中Pb含量远大于地上部。增加Mg或Ca的供给显著地增加了A. murale地上部Pb含量,但对其地下部Pb含量影响不明显;增加Mg或Ca的供给显著增加了S.alfredii地下部Pb含量,但其地上部Pb含量无明显变化;增加Mg的供给显著增加N.caerulescens地上部Pb含量,但其地下部Pb含量显著降低,增加Ca的供给对N. caerulescens地下部和地上部Pb含量无显著影响。
3 讨论
相关研究表明,增加生长基质中Ca或Mg的供给会减少普通植物对Cd、Ni、Mn、Pb等二价重金属的吸收、缓解这些重金属对植物的毒性作用,这些重金属也会影响植物对Ca或Mg的吸收。其相关机制可能有以下几个方面[6-10,17-21]:(1)同为二价离子,半径相近,离子间竞争吸收运输位点[6-7,10,20-21];(2)Ca2+和Mg2+的存在有利于根系细胞维持正常的渗透系统,降低了重金属对矿质营养的胁迫作用[9-10,20];(3)Ca2+或Mg2+增加抗氧化酶的活性、减少脂质的过氧化[7-8];(4)Ca2+与细胞膜氧化胁迫等逆境条件下的信号传导和基因表达相关[8-10];(5)高浓度Ca或Mg对根系分泌的低分子量有机酸成分和浓度有影响[9,18-19]。人们对Ca的影响研究关注多于对Mg和Ca/Mg比的影响,对普通植物的研究多于对超富集植物的研究。相对于普通植物,超富集植物因为长期的进化作用形成了一些机制来增加土壤中重金属的生物有效性,具有较强的地上部转移效率和地上部耐受解毒机制,Ca2+、Mg2+或Ca/Mg比对植物生长和重金属吸收的影响也会有所不同。
Wang等[7]报道营养液中添加5 mmol·L-1CaCl2减弱了0.4、0.6 mmol·L-1Cd2+胁迫下三叶草(Trifolium repens L.)地上部和根系的积累和毒性,机制可能是减轻了脂质的过氧化和刺激了过氧化酶的活性。Magdziak等[19]发现相对于4:1 Ca/Mg比,1:10 Ca/Mg比条件下柳树(Salix viminalis)地上部和根系积累更多的Cd/Pb/Cu/Zn,S.viminalis根系有机酸分泌物(LMWOA)的成分和浓度因Ca/Mg比和胁迫重金属的种类而异,但是LMWOA总浓度与Ca/Mg比之间并非简单的线性关系。对超富集S.alfredii的系列研究发现,其体内Ca与Cd的吸收、转运和分布是相似的,外源添加Ca促进了其地上部对Cd的积累和植物的生长,高Ca供给[6.0 mmol·L-1Ca(NO3)2]减缓了Cd 对S.alfredii根系生长的抑制作用,减少了根系Cd的累积,增加了根系Ca的含量,促进了谷胱甘肽的生成[8,20-21]。
与Cd不同,外源添加Ca、Mg或改变Ca/Mg比对植物必需的元素如Zn和Ni的吸收具有促进作用。黄化刚等[22]报道外源Ca2+对S.alfredii吸收及积累Zn有促进作用。Chaney等[23]模拟蛇纹岩土壤条件下,在水培养营养液中添加0.128~5 mmol·L-1Ca和2 mmol·L-1Mg,分别用31.6~1000 μmol·L-1Ni胁迫A.murale和1.0~10 μmol·L-1Ni胁迫卷心菜,试验结果表明Ca减少了卷心菜对Ni的吸收和毒性,0.8~2 mmol·L-1Ca供给使A.murale获得最高的产量,而且在1000 μmol·L-1Ni胁迫下,过低或过高Ca供给会使该植物表现出中毒的症状,Ca的供给显著影响地上部Ni含量、地上部产量和从根系到地上部的转运系数,但影响并非单向,在不超过2 mmol·L-1Ca时是正向促进作用,Ca的供给再增加则表现抑制作用。与此相似,Ni的另一种超富集植物Alyssum inflatum在低Ca/Mg比(0.11~1.5)的生长基质中仍然生长良好,而且Ca/ Mg比的增加促进了该植物对Ni的吸收[24]。
在本试验条件下,三种植物在正常营养液Ca/Mg比的基础上,增加Ca和Mg的供给大多数情况下都会抑制N.caerulescens对重金属的吸收和生长,其对Ca和Mg的转运系数分别为0.77~1.38、1.43~3.70(见表4),对Mg的转运系数更高,除了供Mg最高的处理(Ca/Mg比为0.2)外,其他处理之间都无显著差异。这说明该植物体内能稳定维持Ca或Mg的转运,增Ca或增Mg都减少了三种超富集元素的累积,可能是Ca或Mg与重金属竞争酸根离子形成稳定的络合物。A.murale地上部Ca/Mg比(2.45~16.74)和其对Ca的转运系数(5.98~11.19)都随营养液中Ca/Mg比供给的增加而不同程度地增大(见表3),并远高于其他两种植物,而地下部的Ca/Mg比低于其他两种植物,说明该植物有很强的吸收Ca并将Ca转运至地上部的能力。增Ca多时也增加其对Ni的超积累,但增加Ca的供给会显著降低其对Mg的转运系数;增Mg对Mg 和Ca的转运系数变化影响都不明显,增Mg显著抑制其对Ni的吸收,也许该植物是利用Mg的吸收运输位点来吸收Ni[25-26]。只有在最高的Ca供给和较高的Mg供给时,S.alfredii地下部和地上部Ca、Mg含量及Ca/Mg比才显著高于其他处理,其对Ca和Mg的转运系数分别为1.24~1.45、0.88~1.66,两种元素的绝大部分处理的转运系数没有显著差异(见表4),说明该植物对Ca、Mg的吸收能较好地调控,保持稳定。增加Ca的供给会增加其地上部和地下部的Zn含量,除最高的Mg供给外,增Mg减少了其地上部和地下部的Zn含量,但该植物最高供Mg时获得最大的Zn 和Cd含量,可能与植物在高Mg时生长较好和本实验设计Mg的最大供给大于Ca有关。
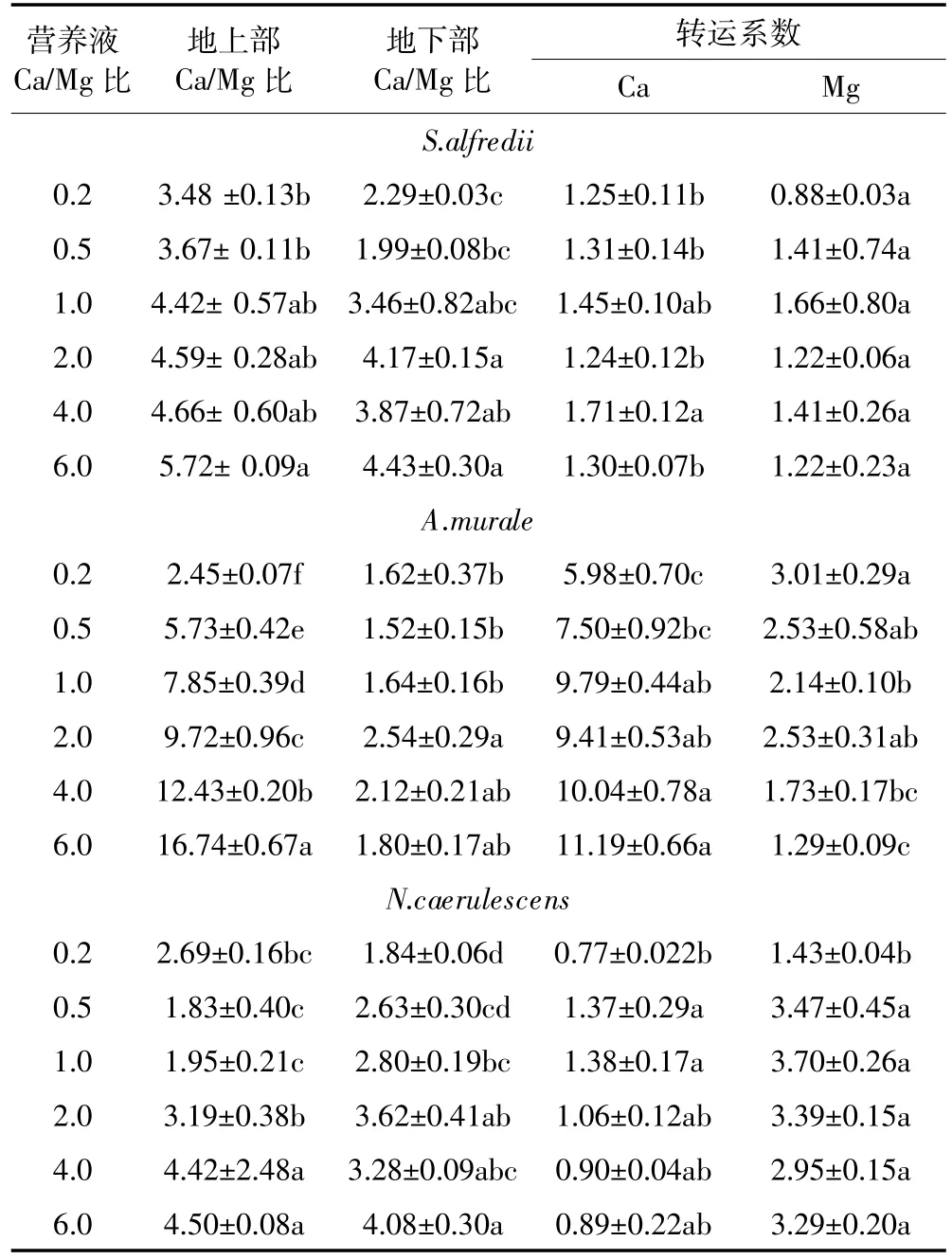
表4 不同Ca/Mg比处理下三种超富集植物Ca/Mg比和转运系数Table 4 Ratios and transfer coefficients of calcium and magnesium in three hyperaccumulators at different calcium to magnesium ratios
4 结论
(1)随着营养液中Ca或Mg供给的增加,三种超富集植物S.alfredii、A.murale和N.caerulescens体内Ca或Mg的含量也相应有不同程度增加,A.murale和N.caerulescens地上部Ca含量都分别大于其地下部,尤其是A.murale有很高的转运系数,营养液中Mg供给的增加显著降低了N.caerulescens地上部Ca含量。其他情况三种植物地上部Ca和Mg含量的变化趋势不明显。
(2)N.caerulescens在正常营养液(Ca/Mg比2)条件下获得最好的生长和地上部最大Zn/Cd/Ni含量,增加Ca或Mg的供给均产生抑制作用。A.murale在高Ca供给时获得最大的生物量和Ni含量,增Ca有助于地上部Ni的累积,但增Mg降低了地上部Ni含量,增Mg和增Ca分别显著降低和增加了地下部对Ni的吸收。增加Ca或Mg的供给均会增加S.alfredii地下部Cd的含量,只有最高的Mg或Ca供给才增加了地上部Cd的含量,其他处理地上部Cd的含量都有所降低;增加Ca的供给会增加其地上部和地下部的Zn含量,除最高的Mg供给外,增Mg减少了其地上部和地下部的Zn含量;S.alfredii在最小的Ca/Mg比(0.2)下获得最大的生物量和最大的Zn和Cd含量。
参考文献:
[1] Adki V S, Jadhav J P, Bapat V A. At the cross roads of environmental pollutants and phytoremediation:A promising bio remedial approach[J]. Journal of Plant Biochemistry and Biotechnology,2014, 23(2):125-140.
[2] Meers E, Tack F M, Van Slycken S, et al. Chemically assisted phytoextraction:A review of potential soil amendments for increasing plant uptake of heavy metals[J]. International Journal of Phytoremediation, 2008, 10(5):390-414.
[3]刘登彪,蒋成爱,张嘉慧,等.不同硼浓度对三种超富集植物吸收硼及重金属的影响[J].农业环境科学学报, 2014, 33(6):1106-1111. LIU Deng-biao, JIANG Cheng-ai, ZHANG Jia-hui, et al. Effects of boron concentrations on uptake of boron and heavy metals by three hyperaccumulators[J]. Journal of Agro-Environment Science, 2014, 33(6):1106-1111.
[4]刘登彪,郑骏宇,赵涛,等.不同供磷水平对四种超富集植物生长及吸收重金属的影响[J].环境科学学报, 2015, 35(4):1198-1204. LIU Deng-biao, ZHENG Jun-yu, ZHAO Tao, et al. Effect of different P levels on the growth and heavy metal uptake for four hyperaccumulators [J]. Acta Scientiae Circumstantiae, 2015, 35(4):1198-1204.
[5] Hepler P K. Calcium:A central regulator of plant growth and development[J]. Plant Cell, 2005, 17(8):2142-2155.
[6] Juang K, Lee Y, Lai H, et al. Influence of magnesium on copper phytotoxicity to and accumulation and translocation in grapevines[J]. Ecotoxicology and Environmental Safety, 2014, 104:36-42.
[7] Wang C Q, Song H. Calcium protects Trifolium repens L. seedlings against cadmium stress[J]. Plant Cell Reports, 2009, 28(9):1341-1349.
[8] Tian S, Lu L, Zhang J, et al. Calcium protects roots of Sedum alfredii H. against cadmium-induced oxidative stress[J]. Chemosphere, 2011, 84 (1):63-69.
[9]汪骢跃,王宇涛,曾琬淋,等. Ca2+和K+对拟南芥幼苗镉毒害的缓解作用[J].植物学报, 2014, 49(3):262-272.WANG Cong-yue, WANG Yu-tao, ZENG Wan-lin. et al. Alleviation of Cd toxicity in Arabidopsis thaliana seedlings by exogenous Ca2+or K+[J]. Chinese Bulletin of Botany, 2014, 49(3):262-272.
[10]蒋廷惠,占新华,徐阳春,等.钙对植物抗逆能力的影响及其生态学意义[J].应用生态学报, 2005, 16(5):971-976. JIANG Ting-hui, ZHAN Xin-hua, XU Yang-chun, et al. Roles of calcium in stress tolerance of plants and its ecological significance[J]. Chinese Journal of Applied Ecology, 2005, 16(5):971-976.
[11]安志装,王校常,施卫明,等.重金属与营养元素交互作用的植物生理效应[J].土壤与环境, 2002, 11(4):392-396. AN Zhi-zhuang, WANG Xiao-chang, SHI Wei-ming. et al. Plant physiological responses to the interactions between heavy metal and nutrients[J]. Soil and Environmental Sciences, 2002, 11(4):392-396.
[12]张习敏,宋庆发,刘伦衔,等.喜钙和嫌钙植物对外源的生长生理响应[J].西北植物学报, 2013, 33(8):1645-1650. ZHANG Xi-min, SONG Qing-fa, LIU Lun-xian, et al. Responses of growth and physiology of calcicoles and calcifuges to exogenous Ca2+[J]. Acta Botanica Boreali-Occidentalia Sinica, 2013, 33(8):1645-1650.
[13]鲁耀,郑波,段宗颜,等.钙镁比调控对烟叶产量、化学品质及镁吸收的影响[J].西北农业学报, 2010,19(11):69-74. LU Yao, ZHENG Bo, DUAN Zong-yan, et al. Effects of Ca/Mg regulation on the flue-cured tobacco yield, chemical quality and magnesium uptake[J]. Acta Agriculturae Boreali-Occidentalis Sinica, 2010,19 (11):69-74.
[14] Jiang C A, Wu Q T, Goudon R, et al. Biomass and metal yield of cocropped Alyssum murale and Lupinus albus[J]. Australian Journal of Botany, 2015, 63(2): 159-166.
[15]杨肖娥,龙新宪,倪吾钟,等.东南景天(Sedum alfredii H):一种新的锌超积累植物[J].科学通报, 2002, 47(13):1003-1006. YANG Xiao-e, LONG Xin-xian, NI Wu-zhong, et al. Sedum alfrdii:A new zine-hyperaccumulator[J]. Chinese Science Bulletin, 2002, 47 (13):1003-1006.
[16]刘士哲.现代实用无土栽培技术[M].北京:中国农业出版社, 2001:124. LIU Shi-zhe. The modern practical soilless cultivation technique[M]. Beijing:China Agriculture Press, 2001:124.
[17] Mleczek M, Gasecka M, Drzewiecka K, et al. Copper phytoextraction with willow(Salix viminalis L.)under various Ca/Mg ratios:Part 1. Copper accumulation and plant morphology changes[J]. Acta Physiologiae Plantarum, 2013, 35(11):3251-3259.
[18] Drzewiecka K, Mleczek M, Gasecka M, et al. Copper phytoextraction with Salix purpurea×viminalis under various Ca/Mg ratios:Part 2. Effect on organic acid, phenolics and salicylic acid contents[J]. Acta Physiologiae Plantarum, 2014, 36(4):903-913.
[19] Magdziak Z, Kozlowska M, Kaczmarek Z, et al. Influence of Ca/Mg ratio on phytoextraction properties of Salix viminalis:Ⅱ. Secretion of low molecular weight organic acids to the rhizosphere[J]. Ecotoxicology and Environmental Safety, 2011, 74(1):33-40.
[20] Lu L L, Tian S K, Yang X E, et al. Enhanced root-to-shoot translocation of cadmium in the hyperaccumulating ecotype of Sedum alfredii[J]. Journal of Experimental Botany, 2008, 59(11):3203-3213.
[21] Lu L L, Tian S K, Zhang M, et al. The role of Ca pathway in Cd uptake and translocation by the hyperaccumulator Sedum alfredii[J]. Journal of Hazardous Materials, 2010, 183(1): 22-28.
[22]黄化刚,李廷轩,张锡洲,等.外源钙离子对东南景天生长及锌积累的影响[J].应用生态学报, 2008, 19(4):831-837. HUANG Hua-gang, LI Ting-xuan, ZHANG Xi-zhou, et al. Effects of exogenous Ca2+on the growth and Zn accumulation of two Sedum alfredii Hance ecotypes[J]. Chinese Journal of Applied Ecology, 2008, 19 (4):831-837.
[23] Chaney R L, Chen K, Li Y, et al. Effects of calcium on nickel tolerance and accumulation in alyssum species and cabbage grown in nutrient solution[J]. Plant and Soil, 2008, 311(1/2):131-140.
[24] Ghasemi R, Ghaderian S M. Responses of two populations of an Iranian nickel-hyperaccumulating serpentine plant, Alyssum inflatum Nyar., to substrate Ca/Mg quotient and nickel[J]. Environmental and Experimental Botany, 2009, 67(1):260-268.
[25] Kazakou E, Dimitrakopoulos P G, Baker A J M, et al. Hypotheses, mechanisms and trade-offs of tolerance and daptation to serpentine soils:From species to ecosystem level[J]. Biology Review, 2008, 83(4):495-508.
[26] Bani A, Pavlova D, Echevarria G, et al. Nickel hyperaccumulation by the species of Alyssum and Thlaspi(Brassicaceae)from the ultramafic soils of the Balkans[J]. Botanica Serbica, 2010, 34(1):3-14.
Effects of Ca/Mg ratios on growth and heavy metal uptake of three hyperaccumulators
ZHAO Tao1, LIU Deng-biao1,2, ZHENG Jun-yu1, JIANG Cheng-ai1,3*
(1.College of Natural Resources and Environment, South China Agricultural University, Guangzhou 510642, China; 2.Shenzhen Techand Ecology & Environment Co.LTD, Shenzhen 518000, China; 3.Key Laboratory of Soil Environment and Waste Reuse in Agriculture of Guangdong Higher Education Institutes, Guangzhou 510642, China)
Abstract:Enhancing biomass and heavy metal uptake of hyperaccumulators would improve soil phytoremediation efficiencies. In this study, we investigated the effects of Ca/Mg ratios(0.2, 0.5, 1.0, 2.0, 4.0, and 6.0)on growth and metal uptake by Sedum alfredii(Zn and Cd hyperaccumulator), Noccaea caerulescens(Zn, Cd and Ni hyperaccumulator)and Alyssum murale(Ni hyperaccumulator)under hydroponic system spiked with 20 mg Zn·L(-1), 18 mg Ni·L(-1), 10 mg Pb·L(-1)and 0.5 mg Cd·L(-1). Results indicated that significant high transfer coefficient of Ca(TF, 5.98~11.19)from roots to shoots and Ca/Mg ratio in shoots(2.45~16.74)were found in A. murale. The TF of Mg(1.43~3.70)was higher than that of Ca(0.77~1.38)for N. caerulescens, while TF of Mg(1.24~1.45)was similar to that of Ca(0.88~1.66)for S. alfredii. For N. caerulescens, the greatest growth and Zn/Cd/Ni accumulation of shoots was obtained in normal nutrient solution(Ca/Mg 2.0), but plant growth and metal accumulation were restrained under increased Ca or Mg supply. For A. murale, the highest biomass and Ni accumulation ofbook=441,ebook=39shoots was observed at the highest Ca/Mg ratio(Ca/Mg 6.0). Increasing Ca supply promoted but increasing Mg supply decreased Ni accumulation in the shoots. For S. alfredii, Zn accumulation in shoots and roots increased with increasing Ca supply, and Cd concentration in roots increased with Ca or Mg supply, but the best growth and the highest Zn and Cd concentrations in shoots were observed at the lowest Ca/Mg ratio(Ca/Mg 0.2). Our results suggest that the effect of Ca/Mg ratio on hyperaccumulators depends on both plant and metal species.
Keywords:Ca/Mg ratio; hyperaccumulator; zinc; nickel; cadmium; lead
*通信作者:蒋成爱E-mail:chaijiang@scau.edu.cn
作者简介:赵涛(1987—),男,湖南衡阳人,硕士研究生,主要研究方向为环境化学和环境污染的治理与修复。E-mail:zhaotaohx@163.com
基金项目:国家自然科学基金(31270675);国家自然科学基金(410713061)
收稿日期:2015-07-31
中图分类号:X53
文献标志码:A
文章编号:1672-2043(2016)03-0440-09
doi:10.11654/jaes.2016.03.005
猜你喜欢
中国甜菜糖业(2022年4期)2023-01-04
载人航天(2021年5期)2021-11-20
四川劳动保障(2021年3期)2021-06-09
今日农业(2020年19期)2020-12-14
现代园艺(2017年13期)2018-01-19
电子制作(2017年20期)2017-04-26
西南农业学报(2016年5期)2016-05-17
中国农业文摘-农业工程(2016年5期)2016-04-12
中国科技信息(2015年2期)2015-11-16
天津护理(2015年4期)2015-11-10
