
水稻种子休眠性及相关基因定位研究进展
2016-04-01 08:12方希林关列梅邓跃军李恩宇
作物研究 2016年2期
方希林,王 悦,关列梅,邓跃军,张 晶,李恩宇*
(1湖南农业大学农学院,长沙410128;2湖南希望种业科技股份有限公司,长沙410126)
水稻种子休眠性及相关基因定位研究进展
方希林1,王 悦1,关列梅1,邓跃军2,张 晶2,李恩宇1*
(1湖南农业大学农学院,长沙410128;2湖南希望种业科技股份有限公司,长沙410126)
摘 要:水稻种子的休眠特性对水稻具有非常重要的生物学意义,并长期影响着水稻的生产。综述了水稻种子休眠的原因及对水稻生产的影响,介绍了水稻休眠基因的研究进展,提出了今后对水稻种子休眠特性需要进一步研究的内容,包括休眠基因的定位和克隆,休眠特性与其他农艺性状的关系,以及抑制物的存在方式等。
关键词:水稻;种子;休眠;基因
种子休眠性是植物在长期进化过程中,为了规避不利环境而演化出来的,并且广泛存在,对农业生产的影响很大,直接关系到农作物的产量和质量。在水稻生产中,种子的休眠性具有双重性:一方面,强休眠性的水稻品种在播种后发芽率低或出苗不齐等,影响生产进程;另一方面,弱休眠性或无休眠性的水稻品种在成熟灌浆期间,在高温高湿的环境下易发生穗发芽现象,导致减产和降低品质[1]。保持种子适度的休眠特性则有利于维持种子的生活力,促进水稻选种、育种进程。因此,研究掌握水稻的休眠特性对水稻生产有重要意义。
1 水稻种子休眠的主要原因
导致种子休眠的原因有多种,有些种子休眠不仅仅是单一因素引起的,很可能是多种因素引起的,且各因素之间相互影响。就水稻种子而言,可能导致休眠的原因主要是胚休眠、种被的障碍、抑制物的存在和不利环境的影响,这几种因素之间关系密切。如:抑制物能够导致胚休眠,外界环境刺激能够影响激素水平,种被能影响抑制物的排出等[2~5]。
1.1 胚休眠
一般来说,胚休眠分为两种不同的类型。一种是种子的胚还未成熟,种子处于休眠状态,胚需要在适宜的条件下继续吸收营养,直到各器官分化完全[6]。另一种是种子中存在代谢缺陷尚未完全后熟,而抑制萌发的物质相对占优势,给予适宜的条件亦不能萌发,这类种子一般经过几周乃至几个月的低温高湿处理才能完成生理后熟,此后才可以解除休眠[7]。一般水稻如果收获较早,成熟度较低,胚未完全成熟的种子容易表现出休眠现象。种子胚休眠的现象在野生稻中发生相对较多,在一般栽培的水稻品种发生较少,有时反而还会发生穗发芽的现象,这往往与水稻品种以及种子的成熟度有关。
1.2 颖壳障碍
颖壳障碍是导致水稻种子休眠的重要原因,主要表现在以下几个方面:第一,阻碍水分的吸收;第二,阻碍气体交换;第三,阻碍了脱落酸(ABA)、香豆素、儿茶酚等化学抑制剂的逸出;第四,颖壳的结构和厚度减少了光线到达胚部的程度;第五,颖壳的机械约束作用[8,9]。曹雅君等[10]对13个水稻主栽品种进行发芽实验表明,当去除水稻颖壳后,发芽率有明显的提高。张桂莲等[11]通过电镜观察发现,具有强休眠性的水稻品种“4628”颖壳背面的细胞排列紧密、茸毛少、孔洞小、萌发孔小,导致种子透水透气性差。
1.3 抑制物的存在与激素水平
水稻种子中存在各种抑制物,当其达到一定浓度时将导致种子休眠。抑制剂的种类多种多样,主要有ABA、有机酸、酚类、醛类等,存在于颖壳、种皮、胚乳和胚中[14,15]。Yano等[16]和Angel等[17]研究表明,ABA是抑制种子萌发的主要激素,主要受NCED与CYP707A基因家族的调控,不同时期基因的表达量不一样,在后熟或者经过处理后可以发现其种子对ABA的敏感性降低,休眠减弱。Okamoto等[18,19]等研究发现,在带有编码ABA的基因CYP707A的突变体转基因系中,当CYP707A过量表达时,其ABA含量大量积累,休眠性明显增强。而GA与ABA有明显的拮抗作用,只有ABA含量降低GA的含量才会增加,在种子后熟的过程中GA的编码基因GA3ox2的表达量会明显升高[20]。此外,其他几种主要激素也都能在一定程度上促进种子的萌发。
1.4 不良环境因素
种子的萌发环境主要指水分、氧气、温度和光照这四个方面,不同种子所要求的环境因素不一样。就一般的水稻种子来说,萌发的最低温度为8~14℃,发芽的最低需水量为26%,低于最低温度或种子体内水分不足时,种子内酶和一些大分子化合物难以活化,种子代谢缓慢,不易萌发。赵明[21]发现,当周围环境中的CO2含量大于30%且缺氧时,呼吸强度维持在较低水平,种子容易进入休眠。
2 种子休眠性对水稻生产的影响
在杂交水稻制种过程中,因种子的弱休眠特性,在高温高湿的条件下经常会发生穗发芽现象,发生率能达到10%~30%,甚至更高[22,23]。有研究认为,穗发芽现象的发生主要与α-淀粉酶有关,当α-淀粉酶活性高时淀粉水解,代谢增强,而GA可能会诱导α-淀粉酶活性增强[24,25]。为了防止穗发芽现象的发生,在制种时可以采取喷施多效唑和ABA,增施氮肥、磷肥,降低种植密度等措施。王小春等[26]试验发现,在齐穗后13~19 d内喷施50 mg/L的ABA能有效防止穗发芽发生。有研究发现,在母本齐穗后14 d左右用0.01%的穗萌抑制剂喷施1~2次,防治效果明显[27]。虽然这些措施可以在一定程度上防止水稻穗发芽的发生,但同时也增加了水稻生产的成本,并且有些化学药剂存在安全等问题。此外,在水稻生产上,种子的弱休眠特性还会影响水稻种子的品质、产量及种子的储藏等,给水稻的生产带来一系列问题。
然而种子的休眠性过强会导致种子发芽率低,出苗不齐,影响生产进程,难以适应现代农业规模化和标准化的要求。虽然在生产上可以采取一些物理的和化学的措施来破除休眠,但也存在诸多问题。首先,增加了生产成本,污染环境。其次,因水稻品种不同其破除休眠的效果也会不同,难以准确把握。陈云风等和陈年伟等[28,29]分别用赤霉素、硝酸钾、双氧水以及加热干燥的方法处理水稻种子,但处理的效果和得出的结果存在差异。此外,一些化学药剂本身还可能会对种子产生毒害作用。戴剑等[30]研究发现,用3%的双氧水处理雨水浸泡过的水稻种子反而使发芽率下降,这可能因为浸泡过的水稻种子颖壳的保护作用减弱或消除了,导致双氧水直接进入胚,产生了毒害作用。
3 水稻种子休眠基因的 QTL定位
种子的休眠性从根本上来说是由其基因决定的。野生稻是栽培稻的始祖,种质资源非常丰富,在进化过程当中,休眠基因的功能并没有丧失。而在栽培稻的驯化过程中,一些品种保留或丧失了这一特性。因此,今后可以利用现代分子生物学手段,并结合育种的需要进行分子育种,培育出具有适当休眠性的品种。
目前,对水稻的休眠基因有了初步的研究,检测到与水稻休眠性相关的QTL大约有154个,在水稻12条染色体上均有分布,主要分布在1、3、5、6、7、11号染色体上(表1),但大多数QTL没有被精细定位和克隆[31]。水稻的休眠特性并不是由一个主效基因单独调控的,而是多个主效或微效基因共同表达,相互加强的结果。在那些具有较强休眠特性的水稻品种中,其休眠主效基因往往不只1个。
Takeuchi等[32]利用日本晴和Kasalath构建的回交群体,将水稻种子休眠基因Sdr1定位在第3染色体上,位于标记R10942到C2045之间,且与标记C1488连锁。Gu等[33]将水稻种子休眠的主效基因qSD12定位在75 kb区间范围内,位于标记RM28642到SD12m50之间,发现携带qSD12基因的基因系的ABA含量比不携带qSD12基因的高。唐九友等[34]通过杂交组合KIlunaze/DV85衍生出重组自交F11家系,通过种子休眠性QTL分析,共检测到4个种子休眠性QTL,分别位于第2(2个)、5 (1个)、11(1个)染色体上,经过50℃干热处理7 d,发现qDOR-5、qDOR-2-1以及qDOR-11对干热敏感,种子易破除休眠,而qDoR-2-2则不敏感。Kazuhiro等[35]利用Nipponbare和Kasalath构建的回交群体,定位出了Kasalath的3个休眠基因:qSD3.1、qSD6.1和qSD7.1,分别位于3、6、7号染色体上,其中qSD6.1对休眠的影响最大。Li等[36]利用强休眠性的Minghui 63和无休眠性的Zhenshan 97,构建近等基因系,将qDGR5a、qDGR5b定位在5号染色体上,qDGR7定位在7号染色体上,qDGE1定位在1号染色体上;利用已克隆的拟南芥的休眠基因DOG1和水稻休眠基因Sdr4进行蛋白的同源性检测,发现qDGE1中含有LOC_Os01g06560,qDGR5b中含有LOC_Os05g48650和LOC_Os05g41280,qDGR7包含了Sdr4基因,而qDGR5a没有检测到同源性。卢丙越[37]以无休眠的水稻品种南粳25为轮回母本,强休眠的N22为父本,构建回交群体BC5F2和BC5F3,最终将休眠基因qsdn-1定位在1号染色体的655 kb区间内,并与标记L27共分离;将休眠基因qsdn-5定位在5号染色体的122 kb区间内,并与标记15-6共分离,并构建了这两个基因的近等基因系,进行发芽实验,NIL(qSdn-1)、NIL(qSdn-5)和NIL(CK)的发芽率分别为23%、35%和98%,进一步验证了qSdn-1和qSdn-5。
Sdr4是水稻休眠基因中克隆较早的一个基因。Sugimoto等[38]以日本晴为背景,并带上Kasalath中含Sdr4基因的7.5 Mb区间,构建了水稻种子休眠基因Sdr4的近等基因系,在2515个群体的遗传分析中,再发现8个重组子,最终将候选基因Sdr4确定在8.7 kb内,发现在Kasalath的3.3 kb内有一个单基因Os07g0585700,通过基因敲除证明Os07g0585700就是Sdr4,进一步研究发现该基因受OsVP1调控,直接影响其表达,从而造成对休眠的影响。Du等[39]研究发现,在经过3个月后熟的水稻种子中,Sdr4基因的表达量明显下降,而GA相关的基因(OsGA20ox1,Os-GA20ox2,OsGA20ox3,OsGA2ox1)和IAA相关的基因(OsILR1,OsGH3-2)的表达量却有所上升。
Ye等[40]将qSD1-2定位在20 kb的范围内,其中含有与种子萌发的相关基因OsGA20ox2,研究发现qSD1-2可能主要是通过调控GA的合成来控制种子的休眠,同时对株高也有影响。还有研究发现,α-淀粉酶活性相关的基因与水稻种子的休眠特性相关的基因在同一个标记附近[41]。这说明水稻休眠基因的表达可能会从多个方面影响水稻,休眠基因也许就是某种酶的调控基因,或者是某种激素的调控基因,只是在不同的内部和外部条件刺激下,可能会导致其所表达的量不同。然而这其中的信号传导和表达机制是相当复杂的,这同时也为研究水稻休眠基因提供了另一种思路。
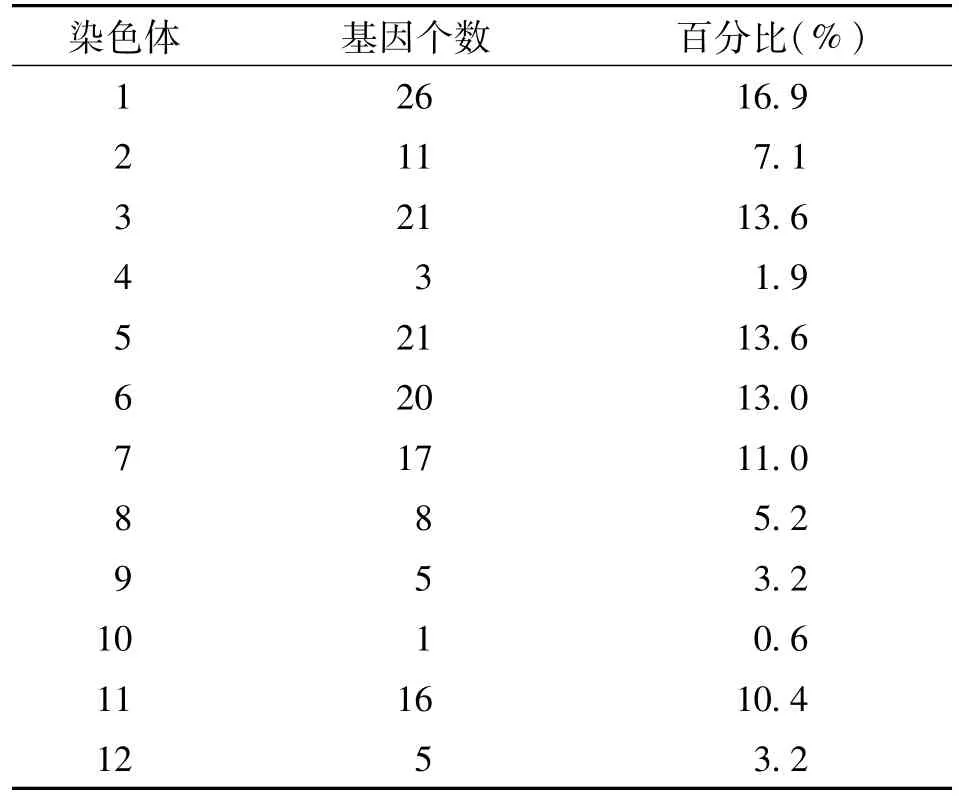
__表 1 水稻各染色体上与休眠性相关 QTL的数量[28]
4 展望
水稻种子的休眠性除了受品种自身的遗传基因控制外,同时也受环境因子的影响。目前,研究者们对水稻种子的休眠原因及破除方法做了大量研究,但水稻种子的休眠特性研究还有许多问题亟需解决,如休眠基因的表达与调控方式、休眠时期种子自身各种抑制物的存在方式与作用、各类休眠相关激素的信号分子等。
在今后的研究中,首先应继续挖掘、定位和克隆更多的休眠基因,并筛选出能够稳定表达的休眠基因,为以后的分子育种奠定基础。其次,因许多休眠基因可能与其他农艺性状的基因存在一定的关联性,可以以其他农艺性状为切入点来进行研究,或许能为休眠基因的研究打开另一条途径。此外,对激素的研究仍需继续深入。目前已从种子萌发与休眠时对各类激素的敏感性进行了研究,在分子层面的研究也有所突破,应逐步理清各类激素与休眠基因表达之间的关系。
为了适应现代水稻生产的基本要求,在未来应充分利用现代分子生物技术,精确掌握水稻的休眠特性,培育出具有适度休眠性的水稻品种,从根本上解决因水稻休眠特性引起的问题,以达到减少生产损失、降低生产成本、便于生产操作的目的。
参考文献:
[1] 沈贝贝.水稻品种N22休眠性QTL qSd-1定位及生理特性解析[D].南京:南京农业大学硕士学位论文,2013.
[2] 潘 琳,徐程扬.种子休眠与萌发过程的生理调控机理[J].种子,2010,29(6):42-47.
[3] 江 玲,万建民.植物激素ABA和GA调控种子休眠和萌发的研究进展[J].江苏农业学报,2007,23(4):360-365.
[4] 杨荣超,张海军,王 倩.植物激素对种子休眠和萌发调控机理的研究进展[J].草地学报,2012,20(1):1 -9.
[5] 颜启传.种子学[M].北京:中国农业出版社,2001.64 -65.
[6] 王方明,张继春,何素华.不同处理方法对水稻恢复系休眠特性的影响[J].种子科技,1997(4):32-33.
[7] 黄 明,陈立云.水稻种子休眠性及其QTL定位研究进展[J].作物研究,2007,21(5):561-566.
[8] 史桂琴,胡辉林.休眠影响种子发芽的生理机制及措施[J].种子世界,1998(2):20-21.
[9] 王丽萍,彭欣怡,程 琴,等.水稻穗发芽与种子休眠研究现状[J].农业研究与应用,2015(4):49-53.
[10]曹雅君,江 玲,罗林广,等.水稻品种休眠特性的研究[J].南京农业大学学报,2001,24(2):1-5.
[11]张桂莲,张顺堂,童佳丽,等.水稻种子休眠生理特性研究[J].中国农学通报,2011,27(27):65-69.
[12]侯冬花,萨拉木·艾尼瓦尔,海利力·库尔班.种子休眠与休眠解除的研究进展[J].新疆农业科学,2007,44(3):349-354.
[13]比尤利JD,布莱克M.种子萌发的生理生化(第二卷):生活力,休眠与环境控制[M].南京:东南大学出版社,1989.45-152.
[14]陈 伟,马绍宾,陈宏伟.种子休眠类型及其破除方法概述[J].安徽农业科学,2009,37(33):16237-16239.
[15]王丽萍.水稻种子的休眠特性研究[D].长沙:湖南师范大学硕士学位论文,2012.
[16]Yano R,Kanno Y,Jikumaru Y,et al.CHOTTO1,a putative double APETALA2 repeat transcription factor,is involved in abscisic acid-mediated repression of gibberellin biosynthesis during seed germination in Arabidopsis [J].Plant Physiology,2009,151(2):641-654.
[17]Angel JM,Nestor CB,María del CRG.An update on the role of NCED and CYP707A ABA metabolism genes in seed dormancy induction and the response to after-ripening and nitrate[J].Plant Growth Regulation,2015,34 (2):274-293.
[18]Okamoto M,Tatematsu K,Matsui A,et al.Genome-wide analysis of endogenous abscisic acid-mediated transcription in dry and imbibed seeds of Arabidopsis using tiling arrays[J].Plant Journal,2010,62(1):39-51.
[19]Okamoto M,Kuwahara A,Seo M,et al.CYP707A1 and CYP707A2,which encod ABA 80-hydroxylases,are indispensable for a proper control of dormancy and germination in Arabidopsis[J].Plant Journal,2006,141:97-107.
[20]杨荣超,张海军,王 倩,等.植物激素对种子休眠和萌发调控机理的研究进展[J].草地学报,2012,20 (1):1-9.
[21]赵 明.水稻收获期休眠性(穗发芽抗性)的鉴定、遗传与育种的应用[D].南京:南京农业大学硕士学位论文,2004.
[22]黄四齐,熊显南,何慕文,等.郴州地区1994年杂交水稻制种减产原因分析及对策探讨[J].杂交水稻,1996(6):17-21.
[23]涂娥英,肖层林.杂交水稻种子特征特性研究[J].杂交水稻,1995(3):15-18.
[24]兰秀锦.小麦新材料RSP的人工合成及其穗发芽抗性研究[D].雅安:四川农业大学硕士学位论文,2005.
[25]胡汉桥.春小麦穗发芽抗性鉴定及机理研究[J].麦类作物学报,2001,31(3):12-17.
[26]王小春,杨文钰.作物种子穗发芽研究现状[J].种子,2003,23(4):51-52.
[27]周新国.杂交水稻制种喷施穗萌抑制剂的效果初探[J].杂交水稻,2003,18(4):37-38.
[28]陈云风,张金才.解除水稻种子休眠效应的研究[J].种子,2014,33(1):87-89.
[29]陈年伟,张体刚,徐昌能,等.解除水稻种子休眠的几种方法比较[J].杂交水稻,2010,25(3):30.
[30]戴 剑,张继红,朱 伟.水稻种子破休眠处理与发芽[J].中国种业,2002(3):25.
[31]谢 坤,白 静,王效睦,等.水稻种子休眠性研究进展[J].作物杂志,2015(5):6-10.
[32]Takeuchi Y,Lin SY,Sasaki T,et al.Fine linkage mapping enables dissection of closely linked quantitative trait loci for seed dormancy and heading in rice[J].Theoretical and Applied Genetics,2003,107(7):1174-1180.
[33]Gu XY,Liu T,Feng J,et al.The qSD12 underlying gene promotes abscisic acid accumulation in early developing seeds to induce primary dormancy in rice[J].Plant Molecular Biology,2010,73(1):97-104.
[34]唐九友,江 玲,王春明,等.水稻种子休眠性QTL定位及其对干热处理的响应[J].中国农业科学,2004,37(12):1791-1796.
[35]Kazuhiro S,Yuri K,Youn C,et al.Confirmation of novel quantitative trait loci for seed dormancy at different ripening stages in rice[J].Rice Science,2013,20(2):110 -116.
[36]Li W,Xu L,Bai XF.Quantitative trait loci for seed dormancy in rice[J].Euphytica,2011,178(3):427-435.
[37]卢丙越.水稻品种N22强休眠性的QTL定位及遗传解析[D].南京:南京农业大学硕士学位论文,2011.
[38]Sugimoto K,Takeuchi Y,Ebana K,et al.Molecular cloning of Sdr4,a regulator involoved in seed dormancy and domestication of rice[J].Proceedings of the National A-cademy of Sciences,2010,107(13):5792-5797.
[39]Du W,Cheng J,Cheng Y,et al.Physiological characteristics and related gene expression of after-ripening on seed dormancy release in rice[J].Plant Biology,2015,17(6):1156-1164.
[40]Ye H,Feng J,Zhang L.Map-based cloning of qSD1-2 identified a gibberellin synthesis gene regulating the development of endosperm-imposed dormancy in rice[J]. Plant Physiol,2015,169(3):2152-2165.
[41]Rathi S,Pathak K,Yadav RNS,et al.Association studies of dormancy and cooking quality traits in direct-seeded indica rice[J].Genetics,2014,93(1):3-12.
Research Progress of Seed Dormancy and Related Gene Mapping in Rice
FANG Xilin1,WANG Yue1,GUAN Liemei1,DENG Yuejun2,ZHANG Jin2,LI Enyu1*
(1 College of Agronomy,Hunan Agricultural University,Changsha,Hunan 410128,China;2 Hunan Hoping Seed Science and Technology Incorporated Company,Changsha,Hunan 410126,China)
Abstract:Seed dormancy characteristics in rice has very important biological significance,and long-term influence on rice production.The reasons of rice seed dormancy and its effects on rice production was summarized,the research progress of dormmancy gene was introduced,the future study about rice seed dormancy research was prospectde.
Keywords:rice;seed;dormancy;gene
基金项目:国家自然科学基金(31301650);湖南省教育厅科学研究项目(13C392);湖南省大学生创新性实验计划项目(XCX14008)。
作者简介:方希林(1995-),男,Email:1975837501@qq.com。*通信作者:李恩宇,助教,主要从事水稻分子育种,Email:lienyu1686@163.com。
收稿日期:2015-10-25
文章编号:1001-5280(2016)02-0210-05
DOI:10.16848/j.cnki.issn.1001-5280.2016.02.24
中图分类号:S511.032
文献标识码:A
猜你喜欢
青少年科技博览(中学版)(2022年6期)2022-12-27
疯狂英语·新读写(2021年10期)2021-12-07
军事文摘(2021年22期)2021-11-26
文苑(2020年6期)2020-06-22
文苑(2019年22期)2019-12-07
儿童时代·幸福宝宝(2019年9期)2019-10-28
奥秘(2019年8期)2019-08-28
商周刊(2017年7期)2017-08-22
莫愁·家教与成才(2017年7期)2017-07-11
小猕猴智力画刊(2016年6期)2016-05-14
