
Relationship of serum GDF11 levels with bone mineral density and bone turnover markers in postmenopausal Chinese women
2016-03-22 06:22:45YusiChenQiGuoMinZhangShuminSongTongguiQuanTiepengZhaoHongliangLiLijuanGuoTiejianJiangandGuangweiWang
Bone Research 2016年1期
Yusi Chen,Qi Guo,Min Zhang,Shumin Song,Tonggui Quan,Tiepeng Zhao,Hongliang Li,Lijuan Guo, Tiejian Jiangand Guangwei Wang
Relationship of serum GDF11 levels with bone mineral density and bone turnover markers in postmenopausal Chinese women
Yusi Chen1,Qi Guo2,Min Zhang2,Shumin Song3,Tonggui Quan2,Tiepeng Zhao2,Hongliang Li2,Lijuan Guo3, Tiejian Jiang3and Guangwei Wang2
Growth differentiation factor 11(GDF11)is an important circulating factor that regulates aging.However,the role of GDF11 in bone metabolism remains unclear.The present study was undertaken to investigate the relationship between serum GDF11 level,bone mass,and bone turnover markers in postmenopausal Chinese women.Serum GDF11 level,bone turnover biochemical markers,and bone mineral density(BMD)were determined in 169 postmenopausal Chinese women(47–78 years old).GDF11 serum levels increased with aging.There were negative correlations between GDF11 and BMD at the various skeletal sites.After adjusting for age and body mass index(BMI),the correlations remained statistically signi f i cant.In the multiple linear stepwise regression analysis,age or years since menopause,BMI,GDF11,and estradiol were independent predictors of BMD.A signi f i cant negative correlation between GDF11 and bone alkaline phosphatase(BAP)was identi f i ed and remained signi f i cant after adjusting for age and BMI.No signi f i cant correlation was noted between cross-linked N-telopeptides of type I collagen(NTX)and GDF11.In conclusion,GDF11 is an independent negative predictor of BMD and correlates with a biomarker of bone formation,BAP,in postmenopausal Chinese women.GDF11 potentially exerts a negative effect on bone mass by regulating bone formation.
INTRODUCTION
Osteoporosis,a classical age-related disease,is more common in women than in men.1–2Osteoporosis affects approximately half of women over the age of 75 years. During the process of aging,estrogen de f i ciency,oxidative stress and lower IGF-1 level induce bone loss and osteoporosis.1–5
Recently,GDF11 was identi f i ed as a circulating ageassociated factor that reverses age-related cardiac hypertrophy and skeletal muscle dysfunction and rejuvenates the aging neurodegenerative and neurovascular brain in mice.6–8GDF11 is a member of the TGF-β super-family that circulates in the bloodstream of mammals.9–11Some studies have shown that serum GDF11 levels in mice decreased during aging.6–8By contrast,Egerman et al.12showed that GDF11 increases with age in rats and humans and has a negative effect on muscle regeneration and satellite cell expansion in mice.Although the results regarding GDF11 are contradictory,it is certain that GDF11 has important roles in age-related diseases,including osteoporosis.
A recent study showed that GDF11 is a protective factor for osteoblastogenesis by inhibiting the activity of peroxisome proliferator-activated receptorγ(PPAR-γ).13However, Li et al.14demonstrated that antagonism of GDF11 activity in the transgenic mice promotes osteoblast activities and bone formation.Thus,the role of GDF11 in bone metabolism remains unclear.
The present study was undertaken to elucidate the relationship of serum GDF11 levels with bone mineraldensity(BMD)and bone turnover markers in postmenopausal Chinese women.
MATERIALS AND METHODS
Subjects
The study population included 169 healthy postmenopausal women(aged 47–78 years)selected randomly from Changsha(China)and its surrounding area.The mean duration from onset of menopause was 7.7±6.4 years with a median menopausal duration of 6(range,1–32)years.All subjects were screened using a detailed questionnaire, and a detailed medical history and physical examination was performed.Body height and weight were measured using a stadiometer and a standardized balance-beam scale,respectively.
Subjects were excluded from the study if they had conditions affecting bone metabolism,such as diseases of the kidney,liver,parathyroid,thyroid,diabetes mellitus, oligomenorrhea,menopause before the age of 40 years, hyperprolactinemia,oophorectomy,rheumatoid arthritis, ankylosing spondylitis,malabsorption syndromes,malignant tumors,or previous pathological fractures.Subjects were also excluded if they were taking glucocorticoids, estrogens,thyroid hormone,f l uoride,bisphosphonate, calcitonin,thiazide,diuretics,barbiturates,or anti-seizure medications.
The study was approved by the Ethical Committee of Xiangya Hospital of Central South University,and all the participants provided written consent to participate.
Serum GDF11 measurement
Blood samples were collected between 7:00 A.M.and 9:00 A.M.after fasting overnight and the samples were allowed to clot.Thereafter,the samples were centrifuged,divided into aliquots,and stored at-70°C until assayed.
To minimize potential cross-reactivity with GDF8,we measured serum GDF11 concentration with a sensitive enzyme-linked immunosorbent assay(ELISA)kit according to Egerman's12methods.High bind 96-well multi-array plates were coated with 50 μL per well of 2 mg·mL-1mouse anti-human GDF11(R&D Systems,clone 743833)in phosphate-buffered saline(PBS)overnight at 4°C.After washing with PBST,300 μL of PBS with 3%bovine serum albumin(BSA)were added to each well and incubated with shaking at room temperature for 1–2 h.The samples were then washed with PBST.An eight-point calibration curve was prepared using three-fold dilutions,starting with a prepared sample of 100 ng·mL-1recombinant human GDF11 in PBS with 1%BSA.PBS with 1%BSA was used as a blank.Samples were diluted 1:2 in PBS with 1%BSA.Fifty microliters of the calibration curve dilutions or samples were added in duplicate to wells and incubated with shaking for 3 h at room temperature.Following a PBST wash,50 μL of a ruthenium-labeled antibody of mouse anti-human GDF11 (clone 743833)diluted to 4 mg·mL-1in PBS with 1%BSA were added to each well and allowed to incubate in the dark with shaking for 2 h at room temperature.After an additional PBST wash step and the addition of 150 μL per well 2×MSD reader buffer with surfactant(4×MSD reader buffer with surfactant diluted 1:2 with distilled water),the plates were read using a μ-Quant Universal Microplate Spectrophotometer(Bio-Tek Instruments,Highland Park, Winooski,VT,USA).Sample concentrations were determined using a four-parameter logistic equation and 1/Y2weighting.The coef f i cient of variation was<10% for GDF11.
BMD measurements
BMD was measured by dual-energy X-ray absorptiometry using a Lunardevice(Lunar DPX IQ,MA,USA)as previously described.15–16The following body parts were scanned:the lumbar spine(L1–L4)and the left hip,including the femoral neck(FN)and total hip(Hip).All BMD results are expressed in g·cm-2.
The precision of the dual-energy X-ray absorptiometry was evaluated over three repeated measurements of different BMD values in 22 subjects,and a mean coef f i cient of variation(CV)for all regions of 0.75%±0.21%(mean±s.d.; range:0.47%–1.02%)was noted.A control spine phantom scan was performed daily with a long-term(more than 10 years)CV of 0.31%–0.43%,and the root-mean-square coef f i cient of variation was 0.35%.
Hormone assays
Serum estradiol levels were determined using a competitive immunoassay,which uses direct chemiluminescence (Automated Chemiluminescence System 180 Estradiol-6 Assay,Bayer HealthCare LLC Diagnostics,Tarrytown,NY, USA).In addition,25-hydroxyvitamin D was measured using ELISA kits(Biomedica Medizinprodukte GmbH&Co KG, Vienna,Austria).Serum intact parathyroid hormone(PTH) was measured using ELISA kits(Diagnostic System,Webster, TX,USA).The coef f i cient of variation was<10%for estradiol,25-hydroxyvitamin D and PTH.
Bone turnover markers measurement
The serum concentration of BAP,as the marker of bone formation,was measured using ELISA kits(BAP from Metra™BAP EIA kit,Quidel Corporation,San Diego,CA,USA).As a marker of bone resorption,serum NTX was measured using ELISA kits(Osteomark,Ostex,Seattle,WA,USA).The coef f i cient of variation was<10%for BAP.

Table 1.Subject characteristics and bone and biochemical parameters of 169 postmenopausal women
Statistical analysis
SPSS 18.0 was used for the statistical analyses.The results are provided as the mean and standard deviation (mean±s.d.).The correlations between GDF11,BMD,and bone turnover markers were calculated using Pearson's correlation analysis and partial correlation analysis.Multivariate linear stepwise analysis(forward selection)was performed to determine whether the level of variance in BMD at various skeletal regions could be explained by age, years since menopause,BMI,smoking habits,estradiol, 25-hydroxyvitamin D,PTH,and GDF11.
RESULTS
Characteristics of the subjects
A total of 169 postmenopausal women were studied. Characteristics of the study participants regarding clinical, biochemical,demographic,and anthropometric characteristics are presented in Table 1.
Correlation between GDF11 and age
As shown in Figure 1,GDF11 serum levels increased with age.A positive correlation was noted between serum GDF11 and age(r=0.601,P<0.01).
Relationships between GDF11 and BMD
Table 2 revealed signi f i cant negative correlations between GDF11 and BMD at the lumbar spine,total hip,and FN,and these results still remained signi f i cant after adjusting for age and BMI.
The parameters identi f i ed as signi f i cant and independent determinant factors of BMD by multiple linear regression analysis are presented in Table 3.The dependent variables were BMD at the lumbar spine,total hip and FN. The independent variables were entered in the regression analysis models,including age,years since menopause, BMI,smoking habit,serum 25-hydroxyvitamin D,PTH,estradiol,and GDF11.In the model with lumbar spine BMD as the dependent variable,the signi f i cantly independent variables were age,estradiol,and BMI,and these variables explained 20.9%of BMD variance(R2=0.209).In the model with total hip BMD as the dependent variable,the signi f i cantly independent variables were years since menopause,BMI, GDF11 and estradiol,and these variables explained 27.4% of BMD variance(R2=0.274).In the model with FN BMD as the dependent variable,the signi f i cantly independent variables were age,GDF11,estradiol,and BMI,and these variables explained 16.9%of BMD variance(R2=0.169). These results indicate that GDF11 was the negative signi f i cant in f l uencing factor for BMD of the total hip and FN.

Table 2.Correlation coef f i cients of serum GDF11 and BMD
Associations between GDF11 and bone turnover markers Serum BAP as a bone formation marker and serum NTX as a bone resorption marker were assayed.As shown in Table 4,a signi f i cant negative correlation was noted between GDF11 and BAP.The correlation remained signi f i cant after adjusting for age and BMI.No signi f i cant correlation was noted between GDF11 and NTX.
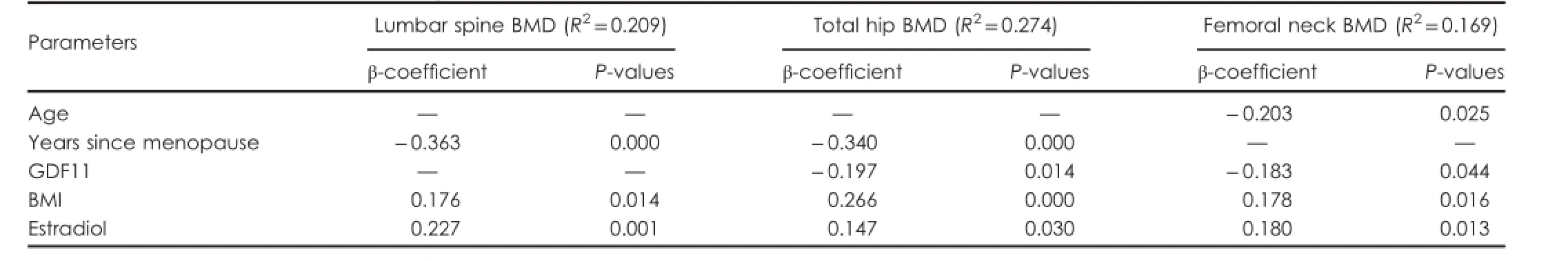
Table 3.Parameters identi f i ed as signi f i cant and independent predictors of BMD
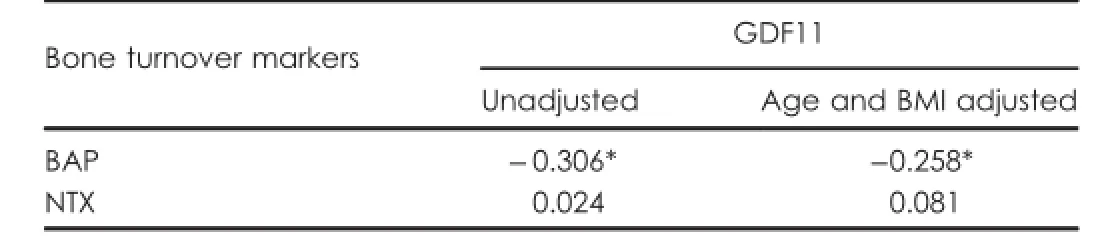
Table 4.Correlation of serum GDF11 levels with bone turnover markers
DISCUSSION
Osteoporosis is an age-related disease.1,17Sex hormone de fi ciencies,endogenous hyperglucocorticoidism,secondary parathyroidism,and oxidative stress induce agerelated osteoporosis.17–18A number of factors have an effect on the process of bone formation.Circulating cytokines also have important roles.Xian et al.5demonstrated that IGF-1 in bone marrow maintains bone mass by activation of mTOR in BMSCs,and bone marrow IGF-1 concentrations were reduced in age-related osteoporosis. Recently,GDF11 was identi f i ed as a circulating ageassociated factor that reverses age-related cardiac hypertrophy and skeletal muscle dysfunction,and rejuvenates the aging neurodegenerative and neurovascular brain in mice.6–8However,Egerman et al.12showed that GDF11 increases with age in rats and humans and has a negative effect on muscle regeneration and satellite cell expansion in mice.Research regarding the role of GDF11 during aging remains contradictory.Our study showed that serum GDF11 levels positively correlated and increased with age. Our results were consistent with Egerman's study,and found that serum GDF11 concentration gradually increased with age.Egerman et al.12developed aGDF11-speci f i c immunoassay to detect the GDF11 concentration in serum of young and aged individuals and revealed a trend toward increased GDF11 levels in serum of aged individuals.The GDF11 antibody used in the previous study exhibited crossreactivity with GDF8 and did notcorrectly measure the GDF11 concentration.Our results were not consistent with the recent study by Zhang et al.,13who showed that GDF11 levels signi f i cantly decreased in the serum of aging mice and human.They also showed thatserum GDF11 levels decreased in women with osteoporosis compared with normal patients.13One possible explanation is that the antibody to GDF11 used by Zhang'study was not speci f i c and that GDF8 was also detected.Our study is consistent with the results obtained by Egerman and showed that GDF11 levels increase with age.
Next,we investigated the relationship between GDF11 and BMD.Our results showed that GDF11 correlated negatively with BMD and was the negative determinant of total hip and FN BMD in postmenopausal women.Our study suggested that GDF11 exerts a negative effect on bone mass.To date,researchers report opposite conclusions about the function of GDF11;however,several studies support our results.Myostatin null mice exhibit increased osteoblast activity and increased bone mass.19–21The GDF11 amino-acid sequence is 90%identical to myostatin (GDF8).22–23Moreover,studies demonstrated that GDF11 and myostatin both induce SMAD2/3 phosphorylation and regulate identical downstream signaling.9–12GDF11 and myostatin similarly inhibit muscle regeneration and decrease satellite cell expansion in mice.12,24Together, these results suggest that GDF11 and myostatin have a similar role in bone metabolism.Li et al.14generated transgenic mice with skeleton-speci f i c overexpression of GDF11 propeptide cDNA,which antagonized GDF11 activity in osteoblasts.These transgenic mice have increased osteoblast activity and enhanced bone formation.Li et al.14suggested that inhibition of GDF11 activity stimulates bone formation by increasing the osteoblast function.
Furthermore,we found that GDF11 negatively correlates with the bone formation biochemical marker,BAP.This fi nding supports the hypothesis that GDF11 may induce bone loss by inhibiting bone formation.
In conclusion,the present study has provided evidence that GDF11 is an independent negative predictor of total hip and FN BMD in postmenopausal Chinese women anddemonstrates a negative correlation with a bone formation biochemical marker.Taken together,these data demonstrated that the increased GDF11 levels in serum with aging predicted bone loss,suggesting a potential role for GDF11 in regulating bone formation.The mechanism should be further clari f i ed.
Acknowledgements
This work was supported by Grant 81570806 from the National Natural Science Foundation of China.
Competing interests
The authors declare no con f l ict of interest.
1 Harvey N,Dennison E,Cooper C.Osteoporosis:a lifecourse approach. J Bone Miner Res 2014;29:1917–1925.
2 Liu Z,Piao J,Pang L et al.The diagnostic criteria for primary osteoporosis and the incidence of osteoporosis in China.J Bone Miner Metab 2002;20:181–189.
3 Li H,Xie H,Liu W et al.A novel microRNA targeting HDAC5 regulates osteoblast differentiation in mice and contributes to primary osteoporosis in humans.J Clin Invest 2009;119:3666–3677.
4 Li CJ,Cheng P,Liang MK et al.MicroRNA-188 regulates age-related switch between osteoblast and adipocyte differentiation.J Clin Invest 2015;125:1509–1522.
5 Xian L,Wu X,Pang L et al.Matrix IGF-1 maintains bone mass by activation of mTOR in mesenchymal stem cells.Nat Med 2012;18:1095–1101.
6 Loffredo FS,Steinhauser ML,Jay SM et al.Growth differentiation factor 11 is a circulating factor that reverses age-related cardiac hypertrophy. Cell 2013;153:828–839.
7 Katsimpardi L,Litterman NK,Schein PA et al.Vascular and neurogenic rejuvenation of the aging mouse brain by young systemic factors.Science 2014;344:630–634.
8 Sinha M,Jang YC,Oh J et al.Restoring systemic GDF11 levels reverses age-related dysfunction in mouse skeletal muscle.Science 2014;344: 649–652.
9 Wharton K,Derynck R.TGFbeta family signaling:novel insights in development and disease.Development 2009;136:3691–3697.
10 Wu MY,Hill CS.Tgf-beta superfamily signaling in embryonic development and homeostasis.Dev Cell 2009;16:329–343.
11 Sartori R,Sandri M.BMPs and the muscle-bone connection.Bone 2015; 80:37–42.
12 Egerman MA,Cadena SM,Gilbert JA et al.GDF11 increases with age and inhibits skeletal muscle regeneration.Cell Metab 2015;22:164–174.
13 Zhang Y,Shao J,Wang Z et al.Growth differentiation factor 11 is a protective factor for osteoblastogenesis by targeting PPARgamma.Gene 2015;557:209–214.
14 Li Z,Kawasumi M,Zhao B et al.Transgenic over-expression of growth differentiation factor 11 propeptide in skeleton results in transformation of the seventh cervical vertebra into a thoracic vertebra.Mol Reprod Dev 2010;77:990–997.
15 Guo LJ,Jiang TJ,Liao L et al.Relationship between serum omentin-1 level and bone mineral density in girls with anorexia nervosa.J Endocrinol Invest 2013;36:190–194.
16 Wang D,Jiang TJ,Liao L et al.Relationships between serum omentin-1 concentration and bone mineral density,and bone biochemical markers in Chinese women.Clin Chim Acta 2013;426:64–67.
17 Riggs BL,Khosla S,Melton LR.Sex steroids and the construction and conservation of the adult skeleton.Endocr Rev 2002;23:279–302.
18 Manolagas SC.From estrogen-centric to aging and oxidative stress:a revised perspective of the pathogenesis of osteoporosis.Endocr Rev 2010; 31:266–300.
19 Hamrick MW.Increased bone mineral density in the femora of GDF8 knockout mice.Anat Rec A Discov Mol Cell Evol Biol 2003;272: 388–391.
20 Hamrick MW,Shi X,Zhang W et al.Loss of myostatin(GDF8)function increases osteogenic differentiation of bone marrow-derived mesenchymal stem cells but the osteogenic effect is ablated with unloading.Bone 2007;40:1544–1553.
21 Hamrick MW,McPherron AC,Lovejoy CO.Bone mineral content and density in the humerus of adult myostatin-de f i cient mice.Calcif Tissue Int 2002;71:63–68.
22 Gamer LW,Wolfman NM,Celeste AJ et al.A novel BMP expressed in developing mouse limb,spinal cord,and tail bud is a potent mesoderm inducer in Xenopus embryos.Dev Biol 1999;208:222–232.
23 McPherron AC,Lawler AM,Lee SJ.Regulation of anterior/posterior patterning of the axial skeleton by growth/differentiation factor 11.Nat Genet 1999;22:260–264.
24 Hamrick MW,Arounleut P,Kellum E et al.Recombinant myostatin (GDF-8)propeptide enhances the repair and regeneration of both muscle and bone in a model of deep penetrant musculoskeletal injury.J Trauma 2010;69:579–583.
This work is licensed under a Creative Commons Attribution 4.0 International License.The images or other third party material in this article are included in the article's Creative Commons license,unless indicated otherwise in the credit line;if the material is not included under the Creative Commons license,users will need to obtain permission from the license holder to reproduce the material.To view a copy of this license,visit http://creativecommons.org/licenses/by/4.0/
©The Author(s)2016
Research(2016)4,16012;
10.1038/boneres.2016.12;Published online:21 June 2016
1Institute of Endocrinology and Metabolism,The Second Xiangya Hospital of Central South University,Changsha,China;2Hunan University of Medicine,Huaihua,China and3Department of Endocrinology,The Xiangya Hospital of Central South University,Changsha,China
Correspondence:Guangwei Wang(wanggwwmq323@hotmail.com)and Tiejian Jiang(jiangtj@tom.com)
Received:13 November 2015;Revised:28 March 2016;Accepted:22 April 2016

- Bone Research的其它文章
- Information for Submission
- Clinical and genetic f i ndings in a Chinese family with VDR-associated hereditary vitamin D-resistant rickets
- External f i xation is more suitable for intra-articular fractures of the distal radius in elderly patients
- In vivo radiometric analysis of glucose uptake and distribution in mouse bone
- TGF-β and BMP signaling in osteoblast,skeletal development,and bone formation,homeostasis and disease
- Assessment of bone turnover and bone quality in type 2 diabetic bone disease:current concepts and future directions