
象山港典型增殖种类的生态容量评估
2016-03-04 07:32杨林林姜亚洲袁兴伟刘尊雷李圣法
海洋渔业 2016年3期
杨林林,姜亚洲,袁兴伟,林 楠,刘尊雷,李圣法
(中国水产科学研究院东海水产研究所,农业部东海与远洋渔业资源开发利用重点实验室,上海 200090)
象山港典型增殖种类的生态容量评估
杨林林,姜亚洲,袁兴伟,林 楠,刘尊雷,李圣法
(中国水产科学研究院东海水产研究所,农业部东海与远洋渔业资源开发利用重点实验室,上海 200090)
为改善象山港生态系统的种群结构和生物多样性,开展科学的增殖放流工作,实现象山港渔业资源的有效养护和修复,根据2011~2014年在象山港开展的渔业资源和生态环境定点调查数据,利用Ecopath with Ecosim 6.4软件构建了包含浮游植物、大型海藻、浮游动物、游泳动物等25个功能组的象山港生态系统的Ecopath模型,大体上涵盖了象山港生态系统能量流动的整个过程。利用模型结果系统分析了象山港生态系统功能的总体特征;并结合食物网结构、能量流动的分析结果,估算了日本囊对虾(Marspenaeus japonicus)、黄姑鱼(Nibea albiflora)、黑棘鲷(Acanthopagrus schlegelii)等象山港典型增殖种类的生态容量。结果表明,象山港生态系统的营养关系较为简单,系统内的物质和能量循环不畅,大量的初级生产力和次级生产力未能进入更高的营养层次;食物网简化,生态系统的成熟度和稳定性偏低,易受外界因素的干扰;渔业资源的服务和产出功能严重退化。在不改变象山港生态系统结构和功能的前提下,本研究评估得出日本囊对虾的增殖生态容量为0.129 210 t·km-2;黄姑鱼的增殖生态容量为0.017 853 t·km-2;黑棘鲷的增殖生态容量为0.115 965 t·km-2。相比于生态系统内现存的生物量,象山港日本囊对虾、黄姑鱼和黑棘鲷均具有较大的增殖空间。
象山港;增殖;Ecopath模型;生态容量
象山港是位于中国浙江省宁波市沿海的一个半封闭的狭长形海湾。海湾内的原始生态环境良好,营养盐丰富,是多种渔业资源的产卵场、育幼场及栖息地[1]。近年来,随着渔业过度捕捞、环境污染以及海洋开发活动的加剧,象山港的渔业资源显著衰退,海洋环境和生态结构发生了较大变化[2],包括渔业资源恢复在内的生态系统保护和修复工作亟待开展。
增殖放流是养护渔业资源、修复渔业环境的重要手段,为当前国内外所普遍采用。而要想实现增殖目的必须增加对生态风险的防控,实现生态型负责任增殖放流[2-4]。当前国内外增殖放流达到预期效果的案例并不多[5]。影响增殖放流效果的几个重要因素是对放流水域生态环境认识的不足;增殖选种没有考虑食物网中各功能组之间的相互关系;放流数量没有考虑生态系统所能支撑的最大容量等。象山港作为海洋牧场示范区,近些年正持续开展多批次、多品种的增殖放流活动[6]。但目前对象山港现阶段生态系统的总体特征、营养相互关系、典型增殖种类生态容量等方面还缺乏足够的认识,在一定程度上影响了增殖放流的效果。鉴于此,本研究根据2011~2014年在象山港水域开展的渔业资源和生态环境定点调查数据,利用Ecopath with Ecosim 6.4(EwE 6.4)软件来构建象山港生态系统的Ecopath模型,在营养相互关系的基础上评估了日本囊对虾(Marspenaeus japonicus)、黄姑鱼(Nibea albiflora)、黑棘鲷(Acanthopagrus schlegelii)等象山港典型增殖种类的生态容量,分析了当前及达到生态容量后生态系统的总体特征,旨在为优化象山港增殖放流策略,开展增殖放流生态风险的适应性管理,实现渔业的可持续发展提供科学依据。
1 材料与方法
1.1 模型原理
Ecopath模型定义的生态系统由若干个基本涵盖生态系统营养级结构和能量流动过程的生物生态学特征相似的功能组构成[7]。模型假定所有功能组能量的输入、输出保持平衡,即生产量等于产出量与死亡量(捕捞死亡和自然死亡)之和。模型以一组线性方程来定义整个生态系统,而各线性方程代表一个功能组:

式中:Bi为功能组i的生物量;Pi为功能组i的生产量;(P/B)i则为功能组i的生产量和生物量的比值;(Q/B)j为消耗量和生物量的比值;EEi为功能组i的营养转换效率;DCij为捕食组j摄食被捕食组i的比例;EXi为功能组i的产出量。
Ecopath模型保证功能组间的能量流动保持平衡,进而定量描述生态系统各个组成部分的生物学参数。构建Ecopath模型输入的基本参数有Bi、(P/B)i、(Q/B)j、EEi、DCij和EXi。Bi、(P/B)i、(Q/B)j、EEi中出现的任一未知数可由模型计算得知,而DCij和EXi则需已知。
1.2 功能组划分
本研究所用数据来源于2011~2014年(4月、7月、11月)在象山港开展的渔业资源和生态环境定点调查的均值。调查的区域及站位分布如图1所示。定点调查的内容为浮游植物、浮游动物、大型海藻、游泳生物、底栖生物的生物量以及环境要素等。调查方法依照《海洋调查规范》[8]进行。游泳动物采样采取桁杆拖虾网作业,调查网具的桁杆长度为7 m,囊网网目尺寸为13 mm,航速约2 km·h-1,调查期间每站拖曳30 min。底栖动物通过箱式采泥器(面积0.04 m2)采集,所得底栖动物通过0.5 mm的套筛淘洗过滤。
本研究主要根据不同种类的个体大小、食性等特性划分功能组。本研究中的象山港生态系统Ecopath模型由25个功能组组成,基本上涵盖了象山港生态系统的营养级结构和能量流动过程,分别为浮游植物、大型海藻、浮游动物、头足类、日本囊对虾、中国明对虾(Fenneropenaeus chinensis)、小型虾类、大型蟹类、小型蟹类、虾姑类、虾虎鱼类、鲆鲽类、鲷类、鲬类、黄姑鱼、其它石首鱼类、中国花鲈(Lateolabrax maculatus)、腐屑食性鱼类、其它浮游生物食性鱼类、其它游泳动物食性鱼类、其它底栖动物食性鱼类、贝类、棘皮类、多毛类和碎屑类(表1)。这其中具有重要经济价值、增殖潜力或生态功能的种类作为一个独立的功能组。其中,日本囊对虾、黄姑鱼被列为单独的功能组,而鲷类包含黑棘鲷和真鲷(Pagrosomusmajor),由于真鲷的生物量极低,因此鲷类的生物量可视为黑棘鲷的生物量。
图1 象山港调查站位示意图Fig.1 Location of Xiangshan Bay sam p ling sites
1.3 模型调试及生态容量估算
能量流动形式以湿重(t·km-2)表示。时间定义为1 a。鱼类及大型无脊椎动物的生物量通过扫海面积法估算。浮游植物湿重通过调查所测Chla浓度值进行换算[9-10]。单种类初级生产力P/B、Q/B值计算参考瞬时总死亡率和经验公式[11-12]。多种类功能组P/B和Q/B值参考纬度和生态系统特征与象山港近似的长江口及毗邻区[13]、杭州湾[14]、东海[15]模型中的类似功能组,并结合渔业数据网站[16]确定。碎屑量采用碎屑和初级生产碳的经验公式计算[17]。功能组的食物组成矩阵数据来源自样本胃含物分析,部分种类参考已有的食性研究结果[18-21]。
Ecopath模型的调试过程实际就是生态系统能量输入和输出保持平衡的过程。模型平衡的基本条件是EE≤1。而输入参数来源的准确性决定了模型置信度的高低。在输入原始数据后反复调整不平衡功能组的食物组成等参数,直至所有EE满足条件,进而获得合理的参数值[22]。在模型取得平衡后,通过不断增加放流种类的生物量,观察系统中饵料生物等功能组的变化,当模型中任意功能组EE>1时,模型打破平衡而改变当前状态,在模型即将不平衡前的生物量值即为增殖种类的生态容量。

表1 象山港生态系统的功能组及优势种类Tab.1 Functional groups and dom inant species in Xiangshan Bay ecosystem

续表1
2 结果与分析
2.1 模型输出
表2为象山港生态系统Ecopath模型的基本输入和输出情况。代表评价模型总体质量的Pedigree指数值为0.543,与其它生态系统已有的模型研究结果相比[23],本次象山港生态系统的建模质量较高。本研究主要基于实际调查,生物量数据的可信度较高。由表2可知,整个象山港生态系统的营养级可分为4级。其中日本囊对虾、黄姑鱼和鲷类(黑棘鲷)的营养级分别为2.59、3.51和3.31,在象山港内属于中高营养级生物。所有功能组生态营养转换效率在0.03~0.89之间。日本囊对虾和黄姑鱼的生态效率较高,分别为0.70和0.69;而鲷类(黑棘鲷)的生态效率较低,仅为0.07。
2.2 敏感性分析
敏感性分析反映的是输入参数对模型的敏感程度。以10%为步长改变输入参数,整个变动范围则在-50%~50%内,分析这种变化对其它参数的影响。结果表明,模型估计参数对同一功能组输入参数的变化最为敏感,影响最大。输入参数与被影响的参数呈正线性相关。各功能组生物量的变化对不同功能组的营养转换效率影响较大。功能组之间的营养联系在一定程度上也决定了估计参数对输入参数变化的敏感度。其中,当浮游动物的B值下降50%,浮游植物的EE值会下降34.49%。日本囊对虾和黄姑鱼B值在±50%的范围内变化时,对生态系统内其它功能组的EE值影响较小,影响最大的小型虾类和鲷类的EE值得变化率也小于10%。同样,鲷类B值的变化对虾姑类EE值的影响较大,当B值变化50%时,虾姑类的EE值会改变27.62%。

图2 象山港Ecopath模型的敏感性分析Fig.2 Sensitivity analysis of Ecopath m odel for Xiangshan Bay ecosystem
2.3 增殖容量评估
日本囊对虾、黄姑鱼和黑鲷是象山港三个典型的增殖种类,调查期间的生物量分别为0.003 1 t·km-2、0.004 6 t·km-2、0.096 t·km-2。敏感性分析的结果表明,日本囊对虾的B值变化对其它功能组EE值的影响较小,可增殖的潜力较大。当日本囊对虾的现有生物量扩大40倍时,生态系统仍然能够保持平衡,但对饵料生物(小型虾类等)的摄食压力也逐渐增大。当日本囊对虾的生物量扩大41.6倍,达到0.129 210 t·km-2时,小型虾类的EE=1;生物量继续增大时小型虾类的EE>1,模型将失去平衡。因此,象山港生态系统能够支撑的日本囊对虾的生态容量为0.129 210 t·km-2。

表2 象山港Ecopath模型的基本参数(粗体为模型得出)Tab.2 M ain input and output(in bold)param eters in Xiangshan Bay Ecopath m odel
而作为营养级较高的掠食者,黄姑鱼生物量的增加对营养级较低的饵料生物的摄食压力加大。当黄姑鱼的生物量扩容3.86倍,达到0.017 853 t·km-2时,模型仍能保持平衡,但中国明对虾的EE值已达到临界值1;生物量继续增大时,中国明对虾的EE值开始大于1,模型将失去平衡。因此,象山港生态系统能够支撑的黄姑鱼的生态容量为0.017 853 t·km-2。同样,黑棘鲷的营养级也较高,达到了3.31,生物量增加对小型虾类等鲷类饵料生物的摄食压力加大。当黑棘鲷的生物量扩大1.21倍时,其它浮游生物食性鱼类的EE=1;生物量继续增大时,其它浮游生物食性鱼类和小型虾类的EE值将大于1,模型将失去平衡。因此,象山港生态系统能够支撑的黑棘鲷的生态容量为0.115 965 t·km-2。
2.4 增殖前后的生态系统特征
象山港生态系统模型当前状态和3个典型增殖种类达到生态容量时的基本统计参数如表4所示。对比发现,大部分的特征参数变化不大或基本相同,当3个增殖种类达到生态容量时,系统的总流量、呼吸量、生产量、消耗量、总生物量均略有增加,但增加量较小,增幅最大的为日本囊对虾,总流量由2 226.61 t·km-2·a-1增加为2 229.18 t·km-2·a-1;呼吸量由603.72 t· km-2·a-1增加为605.13 t·km-2·a-1;生产量由1 043.58 t·km-2·a-1增加为1 044.64 t· km-2·a-1;消耗量由729.10 t·km-2·a-1增加为732.19 t·km-2·a-1;总生物量由20.64 t· km-2·a-1增加为20.77 t·km-2·a-1。输出量、流向碎屑量和净生产量有所降低,降幅最大的同样为日本囊对虾,输出量由314.60 t·km-2·a-1下降为313.18 t·km-2·a-1;流向碎屑量由579.19t·km-2·a-1下降为578.68 t·km-2·a-1;净生产量由314.58 t·km-2·a-1下降为313.17 t· km-2·a-1。以上这些参数虽有所增加或减少,但总体变化不大。
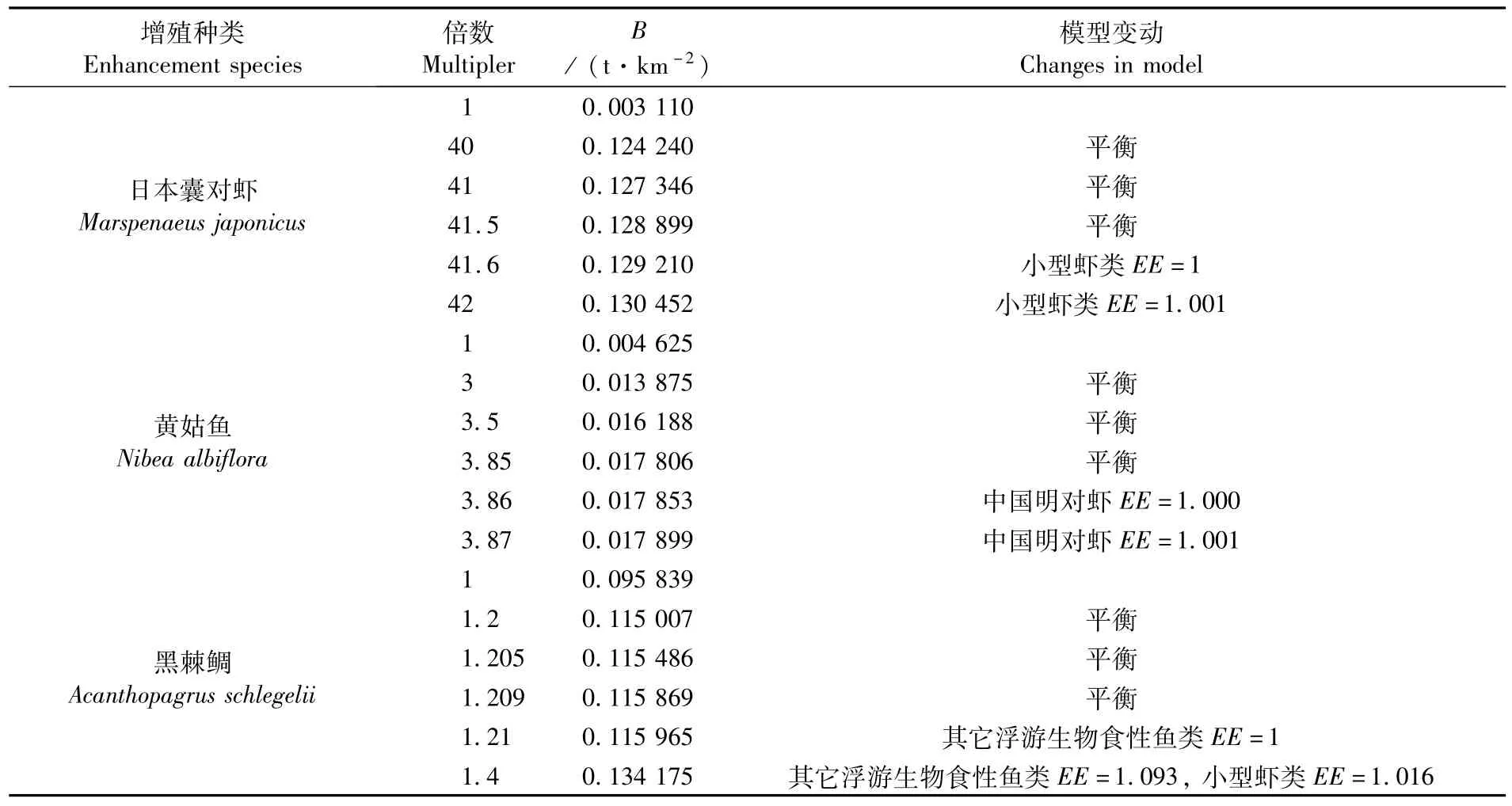
表3 象山港典型增殖种类生物量扩容后的模型变动情况Tab.3 Changes in Ecopath m odel after expanding biom ass of enhancem ent species

表4 增殖前后象山港生态系统的总体特征Tab.4 General characteristics of Xiangshan Bay ecosystem before and after fish resource enhancem ent
此外,净初级生产量前后相同,无变化;系统其它能量及比值也基本差别不大。这些特征表明,当日本囊对虾、黄姑鱼、黑棘鲷的生物量在象山港达到生态容量时,象山港生态系统的结构没有发生大的变化。初级生产量与呼吸量之比(TPP/TR)是生态系统成熟度的重要指标,而连接指数(CI)与系统杂食性指数(SOI)能够反映生态系统内部联系复杂程度。这些指标前后均没有改变或变化极小,这也表明日本囊对虾、黄姑鱼、黑棘鲷达到生态容量时并没有影响到生态系统的稳定性。
3 讨论
近年来,象山港有机污染较为严重,营养盐的含量丰富,富营养化的程度较高,生态系统中富余大量的初级生产力和碎屑[1]。本次生态系统模型的研究结果也印证了这一点。系统的总转换效率仅为3.8%,显著低于10%的林德曼效率[24],大量的初级生产力无法进入更高层次的营养流动。同时次级生产力也过剩,生态系统的营养流动发生阻塞,各营养级间的能量传递受限,转换效率较低。此外,象山港生态系统的TPP/TR、CI和SOI值均偏低,这说明象山港生态系统并不稳定,极易受到外来胁迫;营养关系也较为简单,生物多样性较低,食物网简化。以上这些表明,象山港生态系统在环境恶化和过度捕捞等多重因素的干扰和胁迫下,物质和能量的循环不畅,物种趋于单一,生态系统特别是渔业资源的服务和产出功能已经严重退化。
增殖放流是渔业资源养护、实现渔业可持续发展的重要手段,也是国内外在水生生物资源养护和水域生态修复方面采用的普遍做法。实现生态型负责任增殖放流需要防控增殖放流的生态风险[25],不破坏生态系统的结构和功能,不降低生态系统的健康程度和稳定性。Ecopath生态系统模型为增殖或养殖生态容量的研究提供了一个良好方法。国外学者利用该方法对海湾、泻湖等生态系统内的鱼类和贝类的生态容量进行了广泛研究[26-29];国内也在三疣梭子蟹(Portunus trituberculatus)[30]、中国明对虾[31]、刺参(Apostichopus japonicus)和皱纹盘鲍(Haliotisdiscus hannai)[32]等种类上取得了成功实践。日本囊对虾、黄姑鱼和黑棘鲷是象山港内重要的经济生物,野生资源由于过度捕捞以及环境污染等因素而严重衰退。象山港增殖放流品种涉及多种虾类和鱼类,日本囊对虾、黄姑鱼和黑棘鲷是其中较为典型的增殖种类。研究表明,象山港日本囊对虾的资源密度较低,营养生态效率达到了0.70,大部分被系统所利用。由于初级生产力及碎屑的富余,增加日本囊对虾的生物量不会面临饵料生物的压力,还可以提高低营养级的利用率。在增加象山港生态系统的总生物量的同时,还提高了初级生产者的转化效率。生物量低于生态容量时也不会影响生态系统的结构和功能。象山港目前日本囊对虾的生物量为0.003 11 t·km-2,仅为生态容量的2.41%。因此,象山港日本囊对虾拥有极大的增殖潜力。而黄姑鱼和黑棘鲷由于营养级较高,在象山港属于顶级捕食者,其生物量的增加会对饵料生物造成巨大压力。黄姑鱼和黑棘鲷现存的生物量分别达到生态容量的25.91%和82.64%,增殖潜力低于日本囊对虾,这也是由象山港营养级I到II、II到III的能量转换效率过低,大量初级生产力和次级生产力富余的现状所决定的。
研究认为,增殖放流选种应当首先考虑提高营养级之间的传递效率。除日本囊对虾、黄姑鱼、黑棘鲷等典型增殖种类以外,还需要结合象山港生态系统结构和功能的现状选择合适的增殖品种,提高生态系统的转化效率,改善生态环境,提高系统的稳定性。象山港浮游生物、碎屑的生物量较高,生态效率也不足0.5,港内的网箱养殖活动在一定程度上也加剧了能量的堆积,因此可以通过增加浮游生物食性鱼类和底栖滤食性的贝类的生物量来提高浮游生物和碎屑的利用率,使物质和能量可以有效地往上层传递。除此以外,贝类、多毛类等底栖动物的生态效率较低,大量能量淤积在较低营养级,可以增加一定数量的底栖动物食性鱼类来促进生态系统的能量流动。本研究利用象山港水域Ecopath模型从生态系统的角度阐述了象山港食物网结构、能量流动和系统功能的总体特征,在此基础上评估了日本囊对虾、黄姑鱼、黑棘鲷这三个典型增殖种类的增殖生态容量。该生态容量考虑了增殖种类与捕食者、饵料生物以及食物竞争者间的相关关系,以及增殖放流活动对整个生态系统的影响;对确定象山港日本囊对虾、黄姑鱼和黑棘鲷的合理放流生物量以及环境承载力提供了科学依据,有利于优化增殖放流策略、科学制定增殖放流规划、防控增殖放流的生态风险。但研究得出的增殖种类的生态容量仅为理论上限,实际增殖放流工作在考虑生态效益的同时还要兼顾经济和社会效益,确定放流规模则需要顾及放流活动同时包含多个品种。此外,模型没有考虑各营养级生物的生长变化以及空间变化,如何在静态模型中考虑动态变化以及经济和社会效益,进一步优化评估结果还需要后续深入的研究。
[1] 尤仲杰,焦海峰.象山港生态环境保护与修复技术研究[M].北京:海洋出版社,2011:6-8.
YOU Z J,JIAO H F.Research on water ecological protection and restoration for Xiangshan bay[M].Beijing:China Ocean Press,2011:6-8.
[2] LORENZEN K.Understanding andmanaging enhancement fisheries systems[J].Reviews in Fisheries Science,2008,16(1-3):10-23.
[3] BARTLEY D M,BELL J D.Restocking,stock enhancement and sea ranching:Arenas of progress[J].Reviews in Fisheries Science,2008,16(1-3):357-365.
[4] 姜亚洲,林 楠,袁兴伟,等.象山港游泳动物群落结构及多样性[J].生态学杂志,2013,32(4):920-926.
JIANG Y Z,LIN N,YUAN XW,etal.Community structure and species diversity of nektons Xiangshan Bay of East China[J].Chinese Journal of Ecology,2013,32(4):920-926.
[5] 程家骅,姜亚洲.海洋生物资源增殖放流回顾与展望[J].中国水产科学,2010,17(3):610-617.
CHENG J H,JIANG Y Z.Marine stock enhancement:Review and prospect[J].Journal of Fishery Science of China,2010,17(3):610-617.
[6] 姜亚洲,林 楠,袁兴伟,等.象山港游泳动物群落功能群组成与功能群多样性[J].海洋与湖沼,2014,45(1):108-114.
JIANG Y Z,LIN N,YUAN XW,et al.Functional group composition and functional diversity of nekyon community in the Xiangshan Bay[J].Oceanologia Et Limnologia Sinica,2014,45(1):108-114.
[7] CHRISTENSEN V,WALTERS C J,PAULY D.Ecopath with ecosim:A user’s guide[M].Vancouver,Canada:Fishereis Center,University of British Columbia,2005.
[8] 国家技术监督局.GB 12763.6-91,海洋调查规范[S].北京:标准出版社,1998.
State Bureau of Technical Supervision.GB 12763.6-91,Specification for oceanographic survey[S].Beijing:China Standard Press,1998.
[9] 王增焕,李纯厚,贾晓平.应用初级生产力估算南海北部的渔业资源量[J].海洋水产研究,2005,26(3):9-15.
WANG Z H,LI C H,JIA X P.The fishery production from the north partof the South China Sea using the primary productionmethod[J].Marine Fisheries Research,2005,26(3):9-15.
[10] AHLGREN G.Comparison ofmethods for estimation of phytoplankton carbon[J].Archiv Fur Hydrobiologie,1983,98(4):489-508.
[11] LAFONTAINE J,PETERSR H.Empirical relationships for marine primary production:The effect of environmental variables[J].Oceanologica Acta,1986,9(1):65-72.
[12] CHRISTENSEN V,WALTERS C J,PAULY D.Ecopath with Ecosim version 6 user guide[M].Vancouver:Fisheries Centre,University of British Columbia,2008.
[13] 林 群,金显仕,郭学武,等.基于Ecopath模型的长江口及毗邻水域生态系统结构和能量流动研究[J].水生态学杂志,2009,2(2):28-36.
LIN Q,JIN X S,GUO X W,et al.Study on the structure and energy flow of the Yangtze River Estuary and adjacent waters ecosystem based on Ecopathmodel[J].Journalof Hydroecology,2009,2(2):28-36.
[14] 徐姗楠,陈作志,何培民.杭州湾北岸大型围隔海域人工生态系统的能量流动和网络分析[J].生态学报,2008,28(5):2065-2072.
XU S N,CHEN Z Z,HE P M.Energy flux and network analysis for an artificial ecosystem of a large enclosed sea area in North Hangzhou Bay[J].Acta Ecologica Sinica,2008,28(5):2065-2072.
[15] 欧阳力剑,郭学武.东、黄海主要鱼类Q/B值与种群摄食量研究[J].渔业科学进展,2010,31(2):23-29.
OUYANG L J,GUO X W.Studies on the Q/B values and food consumption of major fishes in the East China Sea and the Yellow Sea[J].Progress in Fishery Sciences,2010,31(2):23-29.
[16] FRÖESER,PAULY D.Fishbase2006:World Wide Web electronic publication[OL].2006,Http://www.fishbase.org.
[17] PAULY D,SORIANO-BARTZ M L.Improved construction,parametrization and interpretation of steady-state ecosystem models[C]//CHRISTENSENV,PAULY D,eds,Trophic models of aquatic ecosystems.ICLAM Conference Proceedings,1993(26):1-13.
[18] 徐善良,王春琳,梅文骧,等.浙江北部海区口虾蛄繁殖和摄食习性的初步研究[J].浙江水产学院学报,1996,15(1):30-35.
XU SL,WANG C L,MEIW X,et al.Preliminary studies on propagation and feeding habits of Oratosquilla oratoria in northern Zhengjiang sea areas[J].Journal of Zhejiang College of Fisheries,1996,15(1):30-35.
[19] 柏怀萍.象山港黑鲷的摄食习性[J].宁波大学学报(理工版),1999,12(4):42-47.
BAI H P.Food habit of black pargy(Sparus microcephalus)in Xiangshan Port[J].Journal of Ningbo University(NSEE),1999,12(4):42-47.
[20] 林 楠,姜亚洲,袁兴伟,等.象山港黄姑鱼的食物组成与摄食习性[J].中国水产科学,2013,20(6):1284-1292.
LIN N,JIANG Y Z,YUAN X W,et al.Diet composition and feeding ecology ofNibea albiflorain Xiangshan Bay,East China Sea[J].Journal of Fishery Sciences of China,2013,20(6):1284-1292.
[21] 魏虎进,朱小明,纪雅宁,等.基于稳定同位素技术的象山港海洋牧场区食物网基础与营养级的研究[J].应用海洋学学报,2013,32(2):250-257.
WEIH J,ZHU X M,JIY N,et al.Study on the food web structure and their trophic levels ofmarine ranching area in Xiangshan Harbor[J].Journal of Applied Oceanography,2013,32(2):250-257.
[22] KAVANAGH P,NEWLANDS N,CHRISTENSEN V,et al.Automated parameter optimization for Ecopath ecosystem models[J].Ecological Modelling,2004,172(2-4):141-149.
[23] MORISSETTE L.Complexity,cost and quality of ecosystem models and their impact on resilience:A comparative analysis,with emphasis on marine mammals and the Gulf of St.Lawrence[D].Vancouver:University of British Columbia,2007.
[24] LINDEMAN R L.The trophic-dynamical aspect of ecology[J].Ecology,1942,23(4):399-418.
[25] 姜亚洲,林 楠,杨林林,等.渔业资源增殖放流的生态风险及其防控措施[J].中国水产科学,2014,21(2):413-422.
JIANG Y Z,LIN N,YANG L L,et al.The ecological risk of stock enhancement and the measures for prevention and control[J].Journal of Fishery Sciences of China,2014,21(2):413-422.
[26] YUN H K.Carrying capacity and fishery resources release in the Bangjukpo Surfzone ecosystem[J].Journal of the Korean Fisheries Society,2003,36(6):669-675.
[27] GAMITO S,KARIM E.Trophic food web and ecosystem attributes of a water reservoir of the Ria Formosa(south Portugal)[J].Ecological Modelling,2005,181(4):509-52.
[28] BYRON C,LINK J,COSTA-PIERCE B,et al.Calculating ecological carrying capacity of shellfish aquaculture using mass-balance modeling:Narragansett Bay,Rhode Island[J].Ecological Modelling,2011,222(10):1743-1755.
[29] BYRON C,LINK J,COSTA-PIERCE B,et al.Modeling ecological carrying capacity of shellfish aquaculture in highly flushed temperate lagoons[J].Aquaculture,2011,314(1-4):87-99.
[30] 张明亮,冷悦山,吕振波,等.莱州湾三疣梭子蟹生态容量估算[J].海洋渔业,2013,35(3):303-308.
ZHANG M L,LENG Y S,LV Z B,et al.Estimating the ecological carrying capacity ofPortunus trituberculatusin Laizhou Bay[J].Marine Fisheries,2013,35(3):303-308.
[31] 林 群,李显森,李忠义,等.基于Ecopath模型的莱州湾中国对虾增殖生态容量[J].应用生态学报,2013,24(4):1131-1140.
LIN Q,LI X S,LI Z Y,et al.Ecological carrying capacity of Chinese shrimp stock enhancement in Laizhou Bay of East China based on Ecopathmodel[J].Chinese Journal of Applied Ecology,2013,24(4):1131-1140.
[32] 吴忠鑫,张秀梅,张 磊,等.基于线性食物网模型估算荣成俚岛人工鱼礁区刺参和皱纹盘鲍的生态容纳量[J].中国水产科学,2013,20(2):327-337.
WU Z X,ZHANG X M,ZHANG L,et al.Predicting the ecological carrying capacity of the Lidao artificial reef zone of Shandong Procince for the sea cucumber,Apostichopus japonicus,(Selenck)and the abalone,Haliotis discus hannai,using a linear food web model[J],Journal of Fishery Sciences of China,2013,20(2):327-337.
Ecological carrying capacity of typical enhancement species in Xiangshan Bay
YANG Lin-lin,JIANG Ya-zhou,YUAN Xing-wei,LIN Nan,LIU Zun-lei,LISheng-fa
(Key Laboratory of East China Sea&Oceanic Fishery Resources Exploitation and Utilization,Ministry of Agriculture,East China Sea Fisheries Research Institute,Chinese Academy of Fishery Sciences,Shanghai200090,China)
To improve the ecosystem structure and biodiversity,develop the stock enhancement scientifically,and realize the fishery resources conservation and ecological restoration in Xiangshan Bay,the paper constructed the energy flow model using the Ecopath with Ecosim(EwE)package based on data collected from fishery resources,environmental and ecological surveys conducted in the Xiangshan Bay during 2011-2014.Themodel was comprised of 25 functional groups covering the main trophic flow of the Xiangshan Bay ecosystem,including phytoplankton,macroalgae,zooplankton,nektons and so on.The paper evaluated the trophic structure,energy flow and the system attribute,then assessed the ecological carrying capacity of Japanese prawn(Marspenaeus japonicus),spotted maigre(Nibea albiflora),and black porgy(Acanthopagrus schlegelii)in Xiangshan Bay.The results indicated that trophic relationship in Xiangshan Bay ecosystem was simple,material and energy circulation was unbalanced,the accumulating primary productivity and secondary productivity continually released and hindered the energy transfer from low trophic levels to high ones.In addition,the system showed a relatively low maturity and stability,and had weak resistance against outside interference.The service and output function of fishery resources in Xiangshan Bay were severely degraded.Under the condition of stable ecosystem construction and sustaining the ecosystem,the carrying capacity and biomass of Japanese prawn was 0.129 210 t·km-2,spotted maigre was 0.017 853 t·km-2and black porgy was 0.115 965 t·km-2.Compared with existing biomass,all three species had a huge potential for continuous enhancement.
Xiangshan Bay;stock enhancement;Ecopath model;ecological capacity
S 917
A
1004-2490(2016)03-0273-10
2015-10-09
国家公益性行业(农业)科研专项(201303047);宁波市重大科技攻关项目(2013C11014);中央级公益性科研院所基本科研业务专项资金(中国水产科学研究院东海水产研究所)(2011T01)
杨林林(1985-),男,江苏南通人,助理研究员,从事渔业资源与生态研究。E-mail:seasun107@126.com
李圣法,研究员。E-mail:shengfa@sh163.net
猜你喜欢
当代水产(2022年8期)2022-09-20
当代水产(2022年5期)2022-06-05
当代水产(2021年8期)2021-11-04
当代水产(2021年4期)2021-07-20
今日农业(2020年19期)2020-12-14
发明与创新·小学生(2020年4期)2020-08-14
学生天地(2017年19期)2017-11-06
发明与创新·小学生(2016年4期)2016-08-04
中国科技信息(2015年2期)2015-11-16
电测与仪表(2014年1期)2014-04-04
