
大尾须卵翅蝗和犁须卵翅蝗的杂交试验初报
2016-02-08 09:30毛本勇徐吉山
大理大学学报 2016年12期
毛本勇,李 淼,徐吉山
(大理大学农学与生物科学学院,云南大理 671003)
大尾须卵翅蝗和犁须卵翅蝗的杂交试验初报
毛本勇,李 淼,徐吉山
(大理大学农学与生物科学学院,云南大理 671003)
对大尾须卵翅蝗和犁须卵翅蝗开展室内受控杂交实验。结果显示,在正交和反交情况下均能产生有生育力的F1代,F1代自交产生F2;F2表征同F1,二者示差特征介于亲本之间。上述结果表明大尾须卵翅蝗和犁须卵翅蝗之间尚未形成生殖隔离,是同一物种的不同亚种;其示差性状的遗传未表现单基因遗传规律中的显隐关系特点。
大尾须卵翅蝗;犁须卵翅蝗;杂交;生殖隔离;物种
大尾须卵翅蝗Caryanda macrocercusa(Mao et Ren,2007)〔1〕隶属于昆虫纲Insecta、直翅目Orthoptera、斑腿蝗科Catantopidae、卵翅蝗亚科Caryandinae,2011年毛本勇等〔2〕将其移入卵翅蝗属Caryanda Stål,1878,模式产地为云南南涧无量山国家级自然保护区。后续调查发现,该种尚分布于凤庆三沟水、临沧五老山、双江大浪坝,呈点状分布状态,是云南地区的特有物种。
犁须卵翅蝗CaryandacultricercaOu,LiuetZheng,2007,目前仅知模式产地为云南永德忙海水库〔3〕,经多次调查未发现其他分布地,为云南特有种,其形态结构和生活环境与大尾须卵翅蝗基本相似。
此二种蝗虫的翅均退化为鳞翅状,已经失去飞行的能力,迁移扩散能力弱,成为典型的地理隔离状态下,适应特殊小生境的小种群物种,是生境破碎化地区小种群物种形成机制研究的适宜材料〔4〕。
本试验旨在通过两种卵翅蝗的正反交和自交实验,考查杂交后代生殖能力及性状的分化状况,从而提供推断此二种蝗虫亲缘关系及其杂交遗传规律的依据。
1 材料及方法
1.1材料大尾须卵翅蝗活体标本2~3龄幼虫40只,采集地:云南省临沧地区凤庆县三沟水,地理坐标24°40′N,99°55′E、海拔:2 186 m,采集时间:2013年6月18日,采集人:毛本勇、李淼、徐吉山。采集地为凤庆县城东北方向的一片山区林缘草地,草地周围有云南松Pinus yunnanensis、蓝桉Eucalyptus globulus等高大乔木,草被层植物丰富,有荩草Arthraxon hispidus、竹叶草Oplismenus compositus等多种禾本科植物及蕨类植物,还有稀疏的低矮灌木。
犁须卵翅蝗活体标本3~4龄幼虫60只,采集地:云南省临沧市永德县忙海水库,地理坐标23°2′N,99°28′E,海拔2 000~2 060 m,采集时间:2013年6月17日,采集人:毛本勇、李淼、徐吉山。小生境与大尾须卵翅蝗的生境相似。
1.2方法
1.2.1 饲养瓶准备 购置直径15 cm,高30 cm的透明塑料瓶,瓶壁、瓶盖打小孔以透气,瓶底垫5 cm厚的原产地土壤〔4〕。
1.2.2 蝗蝻饲养 将野外采集的上述两种蝗蝻带回实验室,置饲养瓶内饲养。每日投喂荩草、竹叶草等禾本科植物,保持相对湿度60%~80%、室温19~25°C,每日换草、清洁粪便、及时检出死虫,逐日观察记录蝗蝻发育情况〔4〕。2013年6月29日犁须卵翅蝗首先羽化,1 d后大尾须卵翅蝗也陆续羽化,于羽化的当日及时将成虫检出,分瓶饲养,以避免同种交配。最后用于杂交实验的共有大尾须卵翅蝗(用“D”表示)15雄13雌和犁须卵翅蝗(用“L”表示)9雄17雌。
1.2.3 正反杂交实验 2013年7月9日开展正反杂交实验。正交实验搭配为D♂(15只)×L♀(17只),置于标记为“正交”饲养瓶中饲养观察;反交实验搭配为L♂(9只)×D♀(13只),置于标记为“反交”饲养瓶中饲养观察。每日观察记录蝗虫生活、交尾、产卵及死亡现象情况。死亡的雄性制作成针插标本或无水乙醇浸泡标本。
1.2.4 自交实验 将正反交实验后的饲养瓶置室内自然温度下,保持湿度为60%~80%。2014年4月上旬卵开始孵化,继续上述常规饲养工作,直至子一代(F1)自交、产卵、死亡。重复上述操作至2015年子二代(F2)孵化、羽化、交配、死亡为止。
采用超景深三维显微系统(VHX-5000)拍摄雄虫腹部末端、外生殖器形态照片。
2 结果
2.1行为文献〔4〕报道了大尾须卵翅蝗的生物学特性。正反交实验和自交实验中,亲本、F1代和F2代蝗虫的觅食、停息、爬动、清洁身体污物、蜕皮、交尾、产卵等活动均与大尾须卵翅蝗相似,未见异常行为。
2.2子代个体数正反交实验得到F1代,F1代自交得到F2代,见表1。
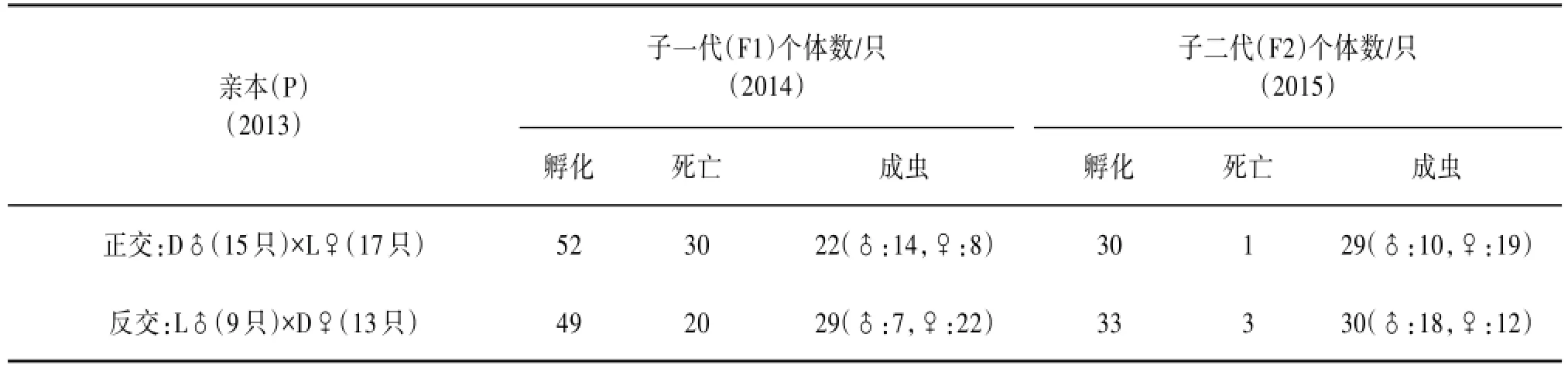
表1 大尾须卵翅蝗和犁须卵翅蝗杂交实验F1代、F2代个体数
由表1可知,正交(D15♂×L17♀)实验(2013年):2014年共孵化出F1代1龄蝗蝻52只,养殖过程中死亡30只,得到成虫22只(♂14只,♀8只),通过自交在2015年共孵化出F2代1龄蝗蝻30只孵化,养殖过程中死亡1只,得到成虫22只(♂10只,♀19只)。反交(L9♂×D13♀)实验(2013年):2014年共孵化出F1代1龄蝗蝻49只,养殖过程中死亡20只,得到成虫29只(♂7只,♀22只);通过自交在2015年共孵化出F2代1龄蝗蝻33只孵化,养殖过程中死亡3只,得到成虫30只(♂14只,♀8只)。
2.3性状分化两种蝗虫的表征差异主要表现在雄性腹部末端和雄性外生殖器形态上〔1-3〕。检视了所有正反杂交实验得到的F1和F2,发现F1和F2的表征相同,其与亲本之间的差别也主要反映在上述雄性特征上。三者之间的区别见表2和图1。

表2 大尾须卵翅蝗、犁须卵翅蝗及其F1的主要区别
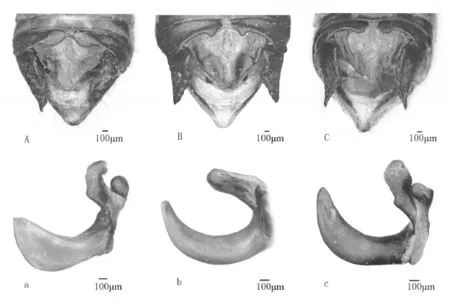
图1 亲本与F1示差特征比较
由表2和图1可知,无论正交或反交得到的F1及F1自交得到的F2,它们的表征均相似。F1及F2与亲本之间的差异主要表现在雄性腹部末端形态,即:第10腹节背板后缘尾片的突出部分较小,狭圆形;肛上板正三角形;雄性尾须宽度适中,上缘近平直,近端部斜切;阳具基背片冠突阔,侧观基部与前突近之间夹角近60°。这些性状均介于两亲本之间。
3 结论及讨论
3.1大尾须卵翅蝗和犁须卵翅蝗的物种地位目前学术界普遍接受的是“生物学物种”概念,认为对于有性生殖生物而言,“物种是一群可以相互进行自由交配的自然种群,并且这些种群和自然界其他的物种的种群之间存在着生殖隔离”〔5〕。Shafer和Wolf的研究〔6〕认为,由地理隔离和遗传漂变导致形成的小种群逐渐适应于不同的生态环境,是为不同的生态型或地理宗。随着时间的推移,这些生态型或地理宗之间的性状逐渐发生分化,最终产生了生殖隔离,并导致了新物种的形成。杂交实验常常用于考查对象间的生殖隔离程度,以此判别它们的亲缘关系〔7-11〕。通常,考查对象如果表型差异,又有不同的分布区,彼此之间却无生殖隔离,应作为不同亚种处理。
本次正交、反交和自交实验中的生殖行为与同种个体之间的生殖行为相似〔4〕;正交和反交能够产生F1,F1自交能够产生F2,F1雌雄虫具有正常的生殖能力,表明大尾须卵翅蝗和犁须卵翅蝗之间仍未形成生殖隔离。大尾须卵翅蝗分布于南涧无量山、凤庆三沟水、临沧五老山及双江大浪坝,基本上沿北纬23°~25°呈点状分布,种群间不存在基因流;犁须卵翅蝗仅分布于永德忙海水库周围。两者分布地之间相隔永德大雪山,种间不存在基因流,符合具有不同分布区的条件。两者之间表型差异明显,符合由地理隔离和遗传漂变介导形成的小种群特点。综上所述,大尾须卵翅蝗和犁须卵翅蝗在自然界不存在基因流,它们表型差异明显,似乎可以作为不同物种看待;但在实验室条件下,它们可以相互配育,产生可育的F1代,按严格的物种定义,它们应该作为同一种的不同亚种对待。它们正处于分化成不同物种的进程中。
然而F2代虽有自交行为,却未产生F3代。F2代是否存在生殖隔离仍需开展重复实验证实。
需要说明的是F1代养殖过程中,可能由于饲养瓶空间狭小,空气流动不畅,导致大约一半的虫体死亡。由于样本数量较少,未能开展杂交组合与同种间生殖力对照实验,故今后仍有开展大样本杂交对照实验的必要。
3.2表型遗传实验结果在F1代和F2代示差鉴别性状介于亲本之间,没有表现单基因遗传规律中的显隐关系特点,提示上述性状可能受多基因〔11〕抑或共显性等位基因控制,这需要深入研究。
〔1〕MAO B Y,REN G D.A new genus and a new species of acrididae(orthoptera)from yunnan,china〔J〕.Entomological News,2007,118(4):366-370.
〔2〕毛本勇,任国栋,欧晓红.云南蝗虫区系、分布格局及适应特性〔M〕.北京:中国林业出版社,2011:74-75.
〔3〕OU X H,LIU Q,ZHENG Z M.Two new species of the genus Caryanda(Orthoptera,Catantopidae)from Yunnan province,China〔J〕.Acta Zootaxonomica Sinica,2007,35(4):758-762.
〔4〕杜燕林,聂秋月,李淼,等.大尾须卵翅蝗生活史及生物学特性观察〔J〕.大理大学学报,2016,1(6):66-68.
〔5〕MAYR E.Animal species and evolution〔M〕.Boston:Belknap Press of Harvard University Press,1963.
〔6〕SHAFER A B,WOLF J B.Widespread evidence for incipient ecological speciation:a meta-analysis of isolation-byecology〔J〕.Ecol Lett,2013,16(7):940-950.
〔7〕朱道弘,ANDO Y.稻蝗属三近缘种间的生殖隔离机制及其进化意义〔J〕.昆虫学报,2004,47(1):67-72.
〔8〕刘春琴,田雷雷,李克斌,等.基于COI基因对三种大黑鳃金龟分类地位的研究〔J〕.应用昆虫学报,2013,50(1):93-100.
〔9〕赵清山,邬文波,吕国平,等.松毛虫的杂交遗传试验〔J〕.昆虫学报,1992,35(1):28-32.
〔10〕叶奕英,许政拱,黄若密,等.广西和云南两地微小按蚊的杂交试验〔J〕.中国寄生虫学与寄生虫病杂志,1992,10(4):287-289.
〔11〕高瑞桐,王保德.光肩星天牛和黄斑星天牛杂交研究〔J〕.北京林业大学学报,2000,22(3):23-26.
A Preliminary Report on the Crossbreed between Caryanda macrocercusa and C.cultricerca
Mao Benyong,Li Miao,Xu Jishan
(College of Agriculture and Biology Science,Dali University,Dali,Yunnan 671003,China)
An indoor hybridization experiment by forced mating between two grasshoppers:Caryanda macrocercusa and C.cultricerca were carried out.Results showed that the reciprocal crosses in the two species could produce fertile progeny F1;these differential characters of F1 same as F2 were intermediate between the parents.The results indicated that the reproductive isolation between C. macrocercusa and C.cultricerca was not yet formed.The two nominal species were the different subspecies of the same species.The heredity of the differential characters didn't conform to the single-gene dominant genetic law.
Caryanda macrocercusa;C.cultricerca;hybridization;reproductive isolation;species
Q969.97
A
2096-2266(2016)12-0053-04
10.3969∕j.issn.2096-2266.2016.12.013
(责任编辑 李 杨)
国家自然科学基金资助项目(31260520;31560605)
2016-08-26
毛本勇,教授,博士,主要从事昆虫系统学及进化生物学研究.
猜你喜欢
今日农业(2022年15期)2022-09-20
疯狂英语·初中天地(2021年6期)2021-08-06
中国糖料(2021年3期)2021-07-13
湖南电力(2021年1期)2021-04-13
热带农业科技(2019年1期)2019-01-14
中国果业信息(2019年11期)2019-01-05
红土地(2018年7期)2018-09-26
中央民族大学学报(自然科学版)(2016年3期)2016-06-27
中央民族大学学报(自然科学版)(2015年2期)2015-06-09
小说月刊(2014年11期)2014-04-18
