
木论喀斯特森林木本植物多样性垂直格局
2016-02-07 00:44兰斯安曾馥平宋同清彭晚霞覃文更
生态学报 2016年22期
兰斯安, 宋 敏,3, 曾馥平, 宋同清,*, 彭晚霞, 杜 虎, 韩 畅,4, 陈 莉,4, 覃文更
1 中国科学院亚热带农业生态研究所亚热带农业生态过程重点实验室, 长沙 410125 2 中国科学院环江喀斯特生态系统观测研究站, 环江 547100 3 湖南农业大学农学院, 长沙 410128 4 湖南农业大学生物科学技术学院, 长沙 410128 5 广西壮族自治区木论国家级自然保护区管理局,环江 547200
木论喀斯特森林木本植物多样性垂直格局
兰斯安1,2, 宋 敏1,2,3, 曾馥平1,2, 宋同清1,2,*, 彭晚霞1,2, 杜 虎1,2, 韩 畅1,2,4, 陈 莉1,2,4, 覃文更5
1 中国科学院亚热带农业生态研究所亚热带农业生态过程重点实验室, 长沙 410125 2 中国科学院环江喀斯特生态系统观测研究站, 环江 547100 3 湖南农业大学农学院, 长沙 410128 4 湖南农业大学生物科学技术学院, 长沙 410128 5 广西壮族自治区木论国家级自然保护区管理局,环江 547200
基于木论国家级自然保护区内西南坡一条垂直样带 16个样方的调查资料,运用丰富度指数、物种多样性指数和均匀度指数对木论喀斯特森林木本植物多样性的垂直格局进行研究。结果表明:(1) 在16个样方中共记录植株5089个,分属48科90属120种。(2) 喀斯特森林木本植物群落具有明显的垂直分布格局,采用等级聚类和非度量多维标度排序方法可将植物群落沿海拔梯度划分为 3个类型,分别是:下坡位以小果厚壳桂(Cryptocaryaaustrokweichouensis)和灰岩棒柄花(Cleidionbracteosum) 为主要优势种的群落;中坡位以长管越南茜(Rubovietnamiaaristata)和罗伞(Brassaiopsis glomerulata)为主要优势种的群落;上坡位以化香树(Platycaryastrobilacea)和密花树(Rapaneaneriifolia)为主要优势种的群落。沿海拔梯度升高,水热组合发生变化,地带性植被以及群落内的建群种和优势种都发生了变化。(3) 物种生活型方面,常绿树种所占的比例高于落叶阔叶树种,针叶树种仅出现在山顶。(4) 沿海拔梯度物种丰富度及α多样性指数中的 Shannon-Wiener 指数曲线均符合单峰格局,峰值出现在坡腰位置。(5)β多样性指数起伏较大,总体上β多样性随着海拔的升高呈递减的趋势。
垂直格局;α多样性;β多样性;木本植物;喀斯特
植物多样性的空间分布格局是植被生态学研究的核心,可以进一步揭示生物多样性的维持机制。例如,山地植物群落的物种多样性如何随海拔梯度的变化一直是生态学热点研究领域之一[1- 3]。而且由于环境因子沿海拔梯度的变化要比沿纬度梯度快1000倍[4]。沿环境梯度不同生活型的物种多样性差异较大,木本植物具有比草本植物更为明确的分布格局[5]。因此,研究木本植物多样性的海拔梯度格局对于揭示生物多样性随环境梯度的变化规律具有重要意义[6- 7]。
桂西北喀斯特山地既是广西生物多样性研究热点区域之一,也是我国生物多样性3个特有分布中心之一[8- 9]。木论国家自然保护区位于广西壮族自治区环江毛南族自治县西北部,与贵州省茂兰国家自然保护区连片构成当今世界上分布面积最广、保护最好、原生性最强的喀斯特非地带性植被区,受地球内动力、强烈的地质运动、高温多雨且分布不均、碳酸盐岩溶蚀性强、水文二维结构明显以及其适生植物具有嗜钙性、耐旱性和石生性等限制特点的综合影响,其生境和植被具有高度异质性。同时,该保护区位于我国植物区系的相汇交错区和交接过渡的中心地带[10]。从地貌类型来看,该区域属于喀斯特峰丛洼地地貌。前人研究表明,峰丛洼地集中分布在西南喀斯特地区南部斜坡地带,面积约12.5万km2[11],其特点为绝对高差小、水热条件差异大、植被垂直变化明显、景观异质性强、岩石裸露率高、土层浅薄、土壤稀少而干燥且以石生植物为主,顶极群落具备生境复杂、植被生产力低、生物种类多样等总体特征[12]。喀斯特原生性森林是喀斯特植被演替的顶级群落,对深入研究喀斯特的植被演替或恢复具有重要参考价值。
尽管诸多学者已对云南哈巴雪山、秦岭太白山、广东鼎湖山、海南尖峰岭以及神龙架地区的植物多样性海拔梯度分布格局进行了研究[13- 18],但其研究结果不一定适用于亚热带喀斯特森林。虽然已有一些研究报导了喀斯特森林植被的空间格局[12,19- 22],但由于受到样地大小或样地形状的限制,尚不能全面反映其多样性与海拔梯度的联系。为此,本研究通过建立一条从洼地到山顶的狭长样带来分析喀斯特森林的群落类型、结构及其物种多样性沿海拔梯度的变化趋势,以期为深入了解喀斯特原生性森林植物群落分布格局、阐明喀斯特退化植物群落的稳定性机制提供科学依据。
1 研究区概况与研究方法
1.1 研究区概况
木论国家自然保护区(107°54′01″—108°05′51″E,25°07′01″—25°12′22″N)位于广西环江毛南族自治县的西北部(图1),东西宽19.80 km,南北长10.75 km,林区总面积89.69 km2,属于中亚热带石灰岩区常绿落叶阔叶混交林生态系统,是我国生物区系相汇交错区和交接过渡的中心,生境异质性极高,区系成分复杂,生物种类丰富,生态环境脆弱,是目前世界上喀斯特地区保存最完好、面积最大的原生林,森林覆盖率达94.8%,其典型景观单元为峰丛洼地。研究区属中亚热带季风气候,年均气温19.3°C,极端最高温36°C,极端最低温- 5°C,1月平均3.4—8.7℃,7月平均23.0—26.7°C,≥10°C年活动积温6260°C,无霜期为310 d。年均日照1451.1 h,占1年可照时数(4422 h)的32.8%,月平均日照以7月份最多(190.7 h),2月份最少(61.5 h),年均降水量1529.2 mm (海拔420 m),降水集中在4—8月,占全年降水量的73.7%,林内阴湿,年均相对湿度79%,干燥度为0.69,湿润度为1.46(K>1.0)。基岩裸露达80%—90%,土被面积不足20%,土壤发育于白云岩、石灰岩的石灰土[23]。

图1 研究区及样地的地理位置Fig.1 The location of study area and the investigated plots
1.2 研究方法
1.2.1 样地设置
本研究旨在反映喀斯特森林植物多样性的海拔梯度格局,因此,在保护区内,从洼地底部至山顶建立一条较为整齐的森林群落样带,样带宽20 m,长320 m,样带最低点海拔448.6 m,最高点海拔633.8 m。坡度范围11.8—57.5 °,主坡向为西南。整个样带包括16个投影面积20 m×20 m的连续样方,样方的4个角用水泥桩做永久标记,在每个20 m×20 m的样方用插值法细分为4个10 m×10 m和16个5 m×5 m的小样方。以5 m ×5 m 小样方为基本测量单元,按顺时针挂牌标记样方内每个胸径(DBH)≥1 cm的木本植物个体,记录物种名称、测定树木的胸径、树高和冠幅、坐标和植物生长状况(斜生、断顶、倒伏等);利用GPS(Trimble XT6000)、手持罗盘测定每一调查样地的经纬度、海拔高度、坡度和坡向。调查样地基本情况见表1。
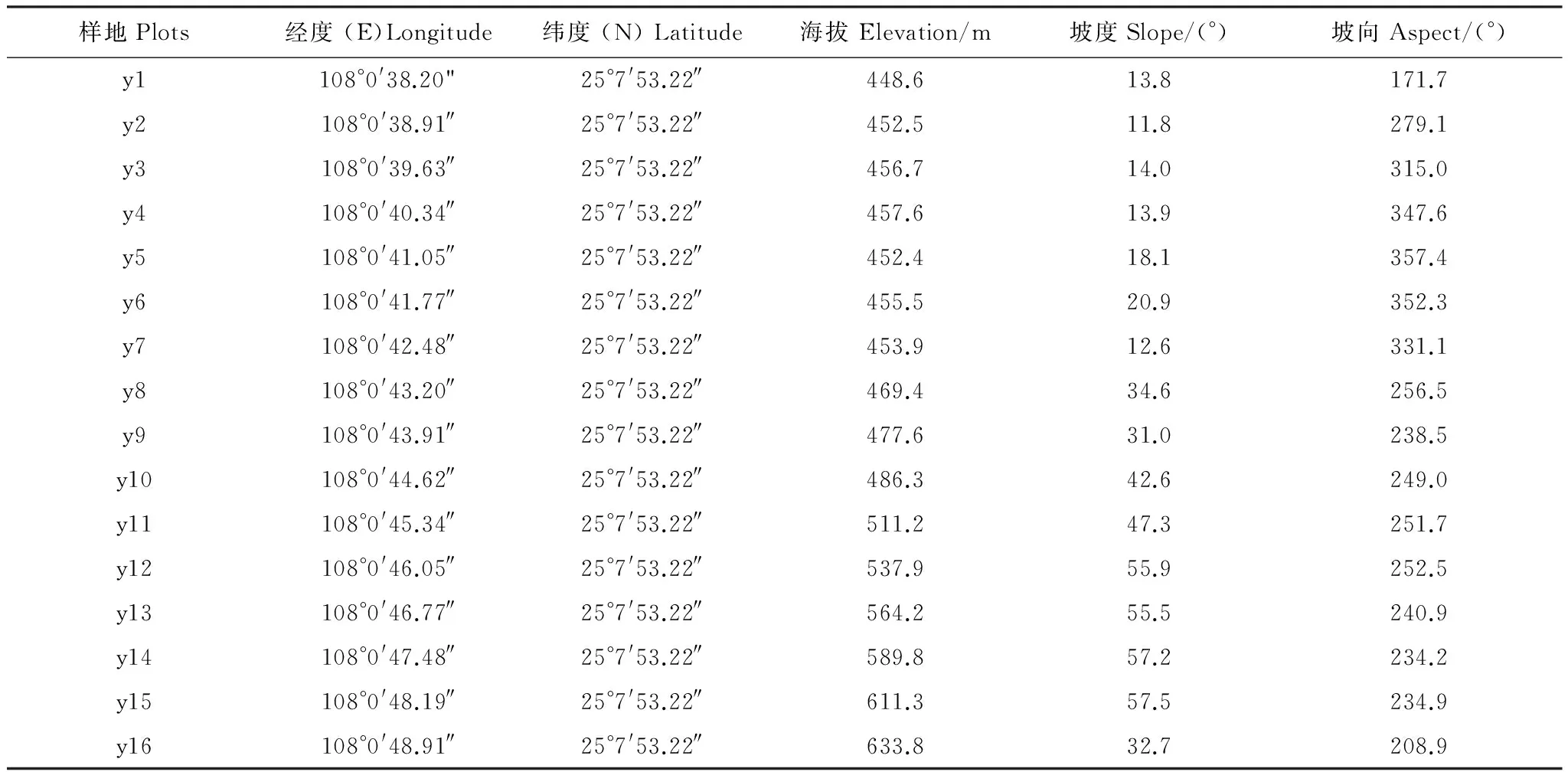
表1 调查样地基本情况
1.2.2 物种丰富度及其结构成分
(1) 物种丰富度 统计样带中维管束植物的科、属、种数,计算各样方内属数/科数、种数/属数、种数/科数的比值。
(2) 生态成分 根据物种的生活型、落叶性划分不同的生态成分,并计算其比值。
1.2.3 群落数量分类和排序
采用等级聚类分析(Hierarchical Cluster Analysis)方法,依据乔木层优势种重要值对样方进行聚类分析;同时,采用非度量多维标度排序(Non-metric Multidimensional Scaling,NMS)法进行矩阵排序。其中重要值计算方法为:
IV=(A+D+F)/3
式中,A为相对多度,指每个种的多度占所有种多度之和的百分比;D为相对优势度,指每个种的胸高断面积之和占所有种个体胸高断面积之和的百分比;F为相对频度,指每个种的频度占所有种频度之和的百分比。
1.2.4 多样性测度
丰富度指数S
S=样方内物种数
多样性指数Pielou均匀度指数
Jsw=-(∑PilnPi)/lnS
Shannon-Wiener指数
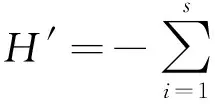
Simpson指数
D=1-∑(Ni/N)2
式中,Pi=Ni/N为第i个物种的个体在样地中所占的比例;N为所有种的个体总数;s为种数;Ni为第i种的个体数。
Bray-Curtis 指数
CN= 2jN/(Na+Nb)
式中,Na为样地a的所有物种个体数目和,Nb为样地b的所有物种个体数目和,jN为样地a和b共有种中个体数目较小者之和,即jN=∑min(jNa,jNb)。
所有统计分析均在R软件(R Development CoreTeam,2009)中进行。
2 结果与分析
2.1 物种组成与分类
2.1.1 群落种类组成
本次调查的16个森林群落样方中乔木层共记录到5089个胸径≥1.0 cm的植株,乔木树种120 种,隶属于48科90属,其中有落叶树种21科37属49种,常绿树种34科58属70种,针叶树种仅有1种,落叶与常绿树种分别占总数的40.83%和58.33%。样地中重要值>10.00的物种共有25种,占总种数的20.8%,占总重要值的28.4%;样带中属于单属、单种的科有17种,占总科数的35.4%,而含有2个属以下的科共有34个,占总科数的70.8%;种数大于5的科有樟科(Lauraceae)、芸香科(Rutaceae)、蔷薇科(Rosaceae)、茜草科(Rubiaceae)和大戟科(Euphorbiaceae)5个科。种数最多的科为大戟科(14种)。
由表2可知,洼地中多以小果厚壳桂(Cryptocaryaaustrokweichouensis)、栀子皮 (Itoaorientalis)为主,岩石裸露率高的样方以灰岩棒柄花(Cleidionbracteosum)、长管越南茜(Rubovietnamiaaristata)、罗伞(Brassaiopsisglomerulata)为主,其次还有密榴木 (Miliusachunii);随海拔上升,陆续出现广西密花树(Rapaneakwangsiensis)、化香树(Platycaryastrobilacea)、密花树 (Rapaneaneriifolia)、罗城鹅耳枥(Carpinusluochengensis),山顶则以清香木(Pistaciaweinmannifolia)、化香树、密花树和米念芭(Tirpitziaovoidea)为主。

表2 各样地重要值大于5的物种
不同分类等级之间的数量比(图2) 基本上反映了该样带不同植物群落类型在物种组成方面的差异。种数/科数、种数/属数、属数/科数的比值是对植物区系分化强度的反映,其值低则表示寡种属多,区系的古老和残遗性强;其值高,则表明多种属比例大,反映区系的物种分化强烈。由图2可知各比例的最大值出现在y11样方(海拔502.1 m)。
2.1.2 群落生活型
从图 3可以看出,针叶树种仅出现在13和14样方,这可能与较高海拔的低温有关。总体来看,在整个海拔梯度上,常绿树种的重要值比重始终远高于落叶树种,表明该区域植物群落构成以常绿树种为主。
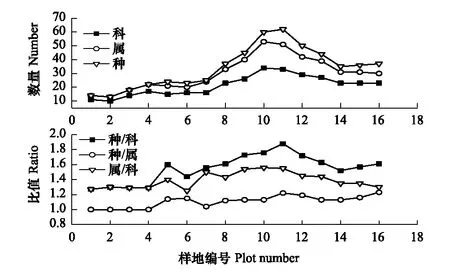
图2 不同系统等级分类单元的多样性梯度Fig.2 Altitudinal gradient of diversity of units at different taxonomic levels

图3 不同生活型植物重要值百分比沿海拔梯度的变化Fig.3 The altitudinal patterns of IV percentage in different life forms
2.1.3 群落分类与排序
综合等级聚类分析和NMS排序结果,16个样地被明显地分为了3个类型:第Ⅰ类,是以小果厚壳桂和灰岩棒柄花为主要优势种的群落,包括1、2、3、4、5、6号样地。第II类,是以长管越南茜和罗伞为主要优势种的群落、包括7、8、9、10、11号样地;第III类,是以化香树和密花树为主要优势种的群落、包括12、13、14、15、16号样地。等级聚类分析和NMS排序方法得出结果的一致,说明分类结果的合理性。另外,数量分类的结果与现地植被类型的分布格局相符,同样印证了分类结果的合理性。
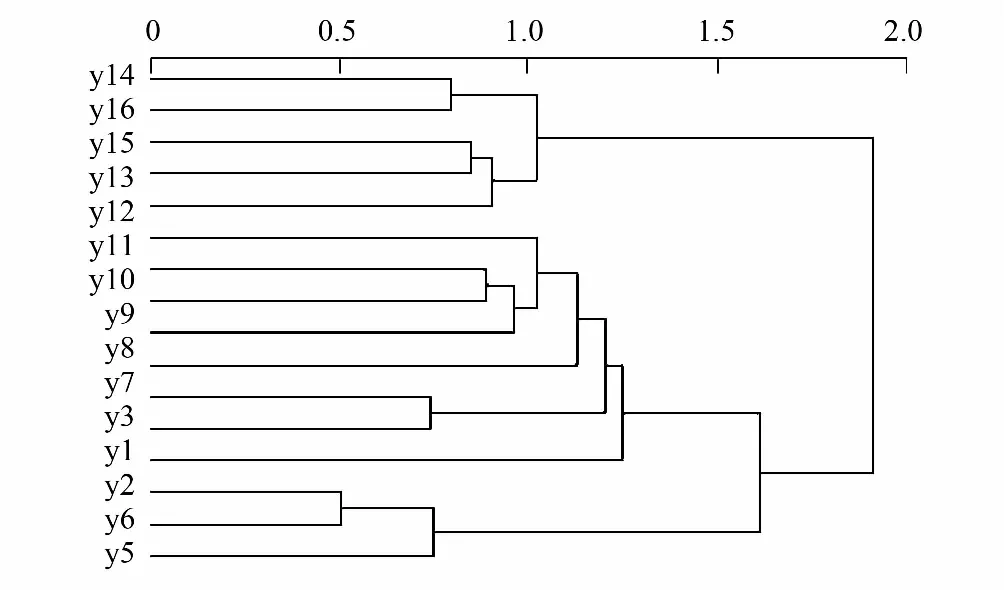
图4 群落样方的聚类分析Fig.4 Cluster analysis classification of the community

图5 非度量多维标度(NMS)排序Fig.5 Non-metric multidimensional scaling (NMS) of the species × attributes matrix
2.2 物种丰富度
图6表示了物种丰富度沿海拔梯度的变化趋势。可以看出,在低海拔地区,随海拔升高,木本植物的丰富度先缓慢增加,从7号样方开始迅速增大,在11号样方达到峰值后开始下降。也就是说,11号样方的分类群丰富度最高。表明该区域物种总数在中等海拔最大,符合“中间高度膨胀(mid-altitude bulge)”理论。
2.3 群落多样性
2.3.1 α多样性
Shannon-Wiener多样性指数与Simpson优势度指数沿海拔梯度表现出一致的趋势(图 7),随着海拔升高,基本呈现出“低-高-低”的单峰分布格局,并均在12号样方达到峰值。
由图8可知,Pielou均匀度指数在研究区域内总体趋势也是随着海拔梯度先升高后逐渐降低,峰值同样出现在12样方。低海拔区域内,随着海拔梯度的升高,外界干扰活动逐渐减少,均匀度开始逐渐升高,在中高海拔区域内,由于生境的异质性和特殊性,只有特定的物种才能生存,物种类型较为单一,均匀度也随之下降。
3个指数都反映出基本一致的变化趋势:即随着海拔的升高先增加后减少,而3个指数均在 6号样方突然减少,可能是由于该样方岩石裸露率高,藤本覆盖面积较大导致木本植物数量较少,且分支较多所致。

图6 丰富度随海拔梯度变化趋势图Fig.6 Changes in species richness along an altitudinal gradient
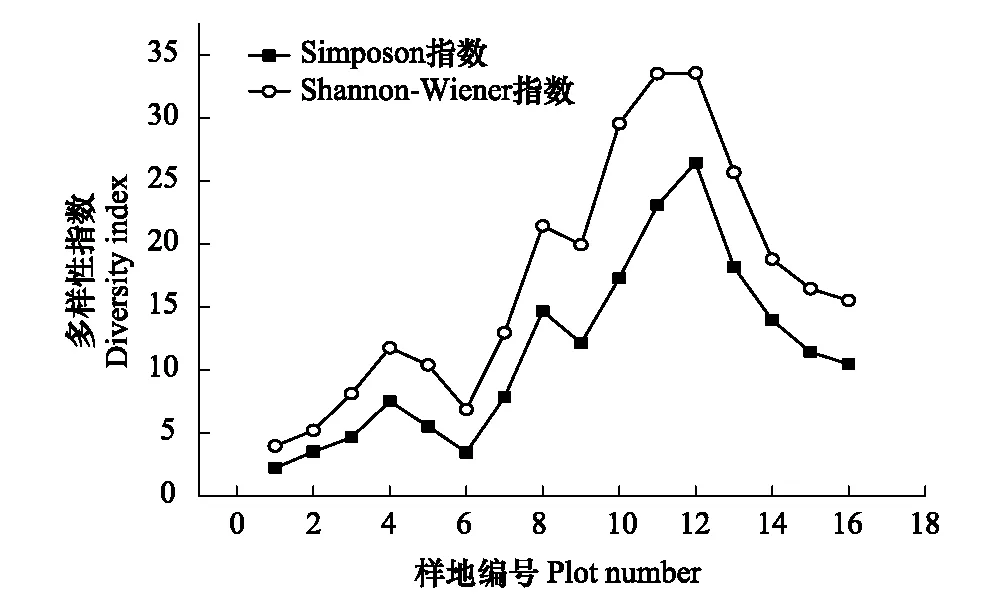
图7 多样性指数随海拔梯度变化趋势图Fig.7 Species diversity along an altitudinal gradient
2.3.2 β多样性
Bray-Curtis指数属于相似性测度,不同地段共有种越多,其β多样性越低,从图9可以看出,1号样地与其它不同海拔样地乔木层的Bray-Curtis指数在海拔梯度上大致呈现先升高后下降再升高的趋势,表明随着海拔落差的增加,其他样方与1号样方的共有物种先增加后减少再增加,β多样性随着海拔的升高减小。出现这种趋势的原因可能是随着海拔升高,植物群落进一步发育,群落物种组成差异性不断增加,群落之间的生态距离逐渐变远,到达一定高度后干扰减少,群落所处的局部小环境更加稳定。

图8 均匀度指数随海拔梯度变化趋势图Fig.8 Pielou′s index along an altitudinal gradient
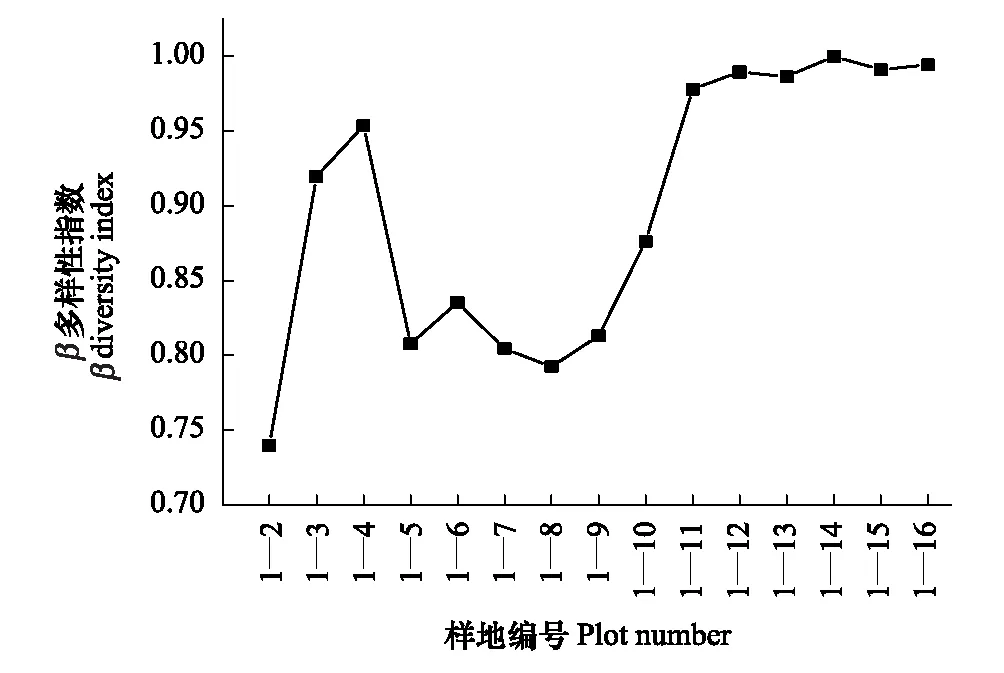
图9 1 号样地与其它样地不同尺度乔木层β样多样性Fig.9 The first quadrat′s beta diversity of tree layer vs that of each of the other quadrats′
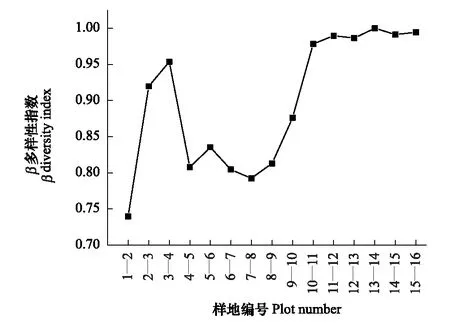
图10 相邻样地乔木层β多样性 Fig.10 The beta diversity in tree layer′ of each two adjacent quadrats
图10显示了沿海拔梯度相邻样地间的β多样性变化趋势,结果表明,相邻样地间种类组成随海拔梯度变化差异比较明显,1—2、4—5及6—7样地间种类组成差异较小,相似性较高,3—4和8—9样地间差异较大,相似性较低。
3 讨论
喀斯特峰丛洼地景观异质性强,岩石裸露率达80%以上,土层浅薄,土壤稀少而干燥,植物多生长在岩石上或石缝中,既要有石生性、喜钙性和耐旱性的特点,又必须具有发达而强壮的根系才能扎根和生长[12]。木论喀斯特森林群落物种组成复杂多样,根据本文研究结果,在16个样地0.64 hm2内统计到48科90属120种植物,与宋同清等[12]在该保护区0.8 hm2内调查到的43科91属123种木本植物的结果基本一致。相比而言:远高于长白山25 hm2阔叶红松林样地的52种[24]和东灵山20 hm2样地的58种[25],低于古田山24 hm2样地[26]、鼎湖山[27]和西双版纳[28]20 hm2样地分别有的159、210和468种。
植物多样性与海拔梯度的关系错综复杂,与群落类型、水热条件、所处的土壤基质和取样尺度等不同有关[1]。先前的研究一般认为:物种多样性具有多种海拔梯度分布格局,其中最普遍的观点是物种多样性的海拔梯度格局与纬度格局相似,即随着海拔上升物种多样性降低[15,29- 30],但也有研究表明随着海拔升高,物种多样性呈现单峰分布格局,即物种多样性随着海拔上升先增加、后减少[31- 33]或呈现出更为复杂的变化规律[34- 35]。
相对海拔是影响喀斯特山地水热条件分布的主要因素,它通过影响光照、水分及土壤资源的再分配进而影响植物群丛的分布及结构[36]。本研究所选样带的α多样性与常见的随海拔升高而下降的模式[30]不同,表现出与佛坪自然保护区[37]和五鹿山国家级自然保护区物种多样性海拔格局[38]一致的“单峰格局”。主要原因可能是:由于样地选址位于喀斯特峰丛洼地,处于亚热带区域,海拔最高不超过700 m,随着海拔升高,温度下降不明显,仍然属于适合植物生长的范围内,喀斯特山地地形复杂、坡位变化及其它环境资源因子分配差异大,低海拔洼地,土层较深厚、湿度条件较好,但太阳直射光照时间不长,一般仅限于某些喜荫和耐荫性强的种类,因而较相对于光照充足的相邻的中等海拔山坡多样性和物种种类组成都低。而在高海拔地段山体陡峭、少土、日照时间长导致土壤蒸发强烈,造成严重干旱缺水等不利植物定居的恶劣环境,因而多样性随海拔变化而呈现下降趋势。另外海拔梯度实际上在一定程度反映了水热状况的梯度变化,因此海拔的影响实际反映了气候因子的综合影响[39]。很多情况下,在低海拔地段,植物生长季节时所需热量充足而水分常常不足;在高海拔地段,水分充足而热量可能成为植物分布的限制因子。相对于低海拔和高海拔,在中海拔地段可能是热量和水分组合最好的生境,从而使得资源的可利用率较高[32]。6号样方出现的多样性突然降低是由于该样地的特殊地形,因此在以后的研究中应该更多的考虑局部小环境的影响。
β多样性表示沿某一环境梯度物种替代速率,能够较直观地反映不同群落间物种的组成差异和不同生境间的梯度变化[40]。在对β多样性随海拔梯度变化格局的研究中,数量数据测度和二元属性数据法各有利弊,不同学者也得出了不同结论:鼎湖山群落β多样性随海拔呈无规律变化[16];南岭国家级自然保护区森林群落β多样性随海拔升高而下降[41];长白山北坡植物群落β多样性随群落间海拔差的增加而增大[42]。但生态位理论认为群落相似性的降低是由于环境异质性的增加[33],中性理论则认为扩散限制产生了β多样性[30]。所以本研究区域的森林群落β多样性符合随海拔升高而减小的常见格局[16]。
本研究选取的木论保护区属于典型的喀斯特峰丛洼地地貌,从洼地到山顶的绝对高度不大,水热条件差异大,植被垂直变化明显。从低海拔往高海拔16个样地被明显地分为了3个类型:第一类是洼地及其边缘,由于光照时间不长,比较荫蔽,湿度条件较好,故分布着以小果厚壳桂和灰岩棒柄花为主要优势种的喜湿和耐荫性强的植物群落。第二类,是山坡山腰地段,这里因太阳直射时间比洼地长,土壤较少,土层较浅薄,保水性能较差,湿度条件不如洼地,群落主要优势种为长管越南茜和罗伞;第三类,是山顶及其周围,这里环境条件恶劣,全天受到太阳直射,温度高,土壤极少,岩石裸露,加之山顶上风大、保水性能和湿度条件最差,群落主要优势种为化香树和密花树等耐旱植物。因此,在喀斯特山地植被恢复时,要根据植物群落物种丰富度的海拔梯度规律,科学、合理地进行立地类型划分,选择适宜栽种物种,制订相应技术和管理措施,开展高效的植物群落营建和封育管护,才能达到费省效宏的效果。
致谢:样地调查得到木论国家级自然保护区管理局工作人员和中南林业科技大学2012级生态学专业实习同学的大力支持,特此致谢。
[1] 方精云. 探索中国山地植物多样性的分布规律. 生物多样性, 2004, 12(1): 1- 4.
[2] Brown J H. Mammals on mountainsides: elevational patterns of diversity. Global Ecology and Biogeography, 2001, 10(1): 101- 109.
[3] Hegazy A K, El-Demerdash M A, Hosni H A. Vegetation, species diversity and floristic relations along an altitudinal gradient in south-west Saudi Arabia. Journal of Arid Environments, 1998, 38(1): 3- 13.
[4] Küchler A W. Review: vegetation of the Earth. Ecology, 1980, 61(1): 206- 206.
[5] O′Brien E M. Climatic gradients in woody plant species richness: towards an explanation based on an analysis of southern Africa′s woody flora. Journal of Biogeography, 1993, 20(2): 181- 198.
[6] 唐志尧, 方精云. 植物物种多样性的垂直分布格局. 生物多样性, 2004, 12(1): 20- 28.
[7] 朱源, 康慕谊, 江源, 刘全儒. 贺兰山木本植物群落物种多样性的海拔格局. 植物生态学报, 2008, 32(3): 574- 581.
[8] 苏宗明, 李先琨. 广西岩溶植被类型及其分类系统. 广西植物, 2003, 23(4): 289- 293.
[9] Hou M F, López-Pujol J, Qin H N, Wang L S, Liu Y. Distribution pattern and conservation priorities for vascular plants in Southern China: Guangxi Province as a case study. Botanical Studies, 2010, 51(3): 377- 386.
[10] 郑颖吾. 木论喀斯特林区概论. 北京: 科学出版社, 1999: 36-38.
[11] 蒋忠诚, 李先琨, 曾馥平. 岩溶峰丛洼地生态重建. 北京: 地质出版社, 2007: 16-17.
[12] 宋同清, 彭晚霞, 曾馥平, 王克林, 曹洪麟, 李先琨, 覃文更, 谭卫宁, 刘璐. 喀斯特峰丛洼地不同类型森林群落的组成与生物多样性特征. 生物多样性, 2010, 18(4): 355- 364.
[13] 沈泽昊, 胡会峰, 周宇, 方精云. 神农架南坡植物群落多样性的海拔梯度格局. 生物多样性, 2004, 12(1): 99- 107.
[14] 陶晶, 臧润国, 余昌元. 云南哈巴雪山植物群落和植物多样性海拔梯度分布格局. 林业科学, 2011, 47(7): 1- 6.
[15] 唐志尧, 方精云, 张玲. 秦岭太白山木本植物物种多样性的梯度格局及环境解释. 生物多样性, 2004, 12(1): 115- 122.
[16] 林国俊, 黄忠良, 竺琳, 欧阳学军. 鼎湖山森林群落β多样性. 生态学报, 2010, 30(18): 4875- 4880.
[17] 李林, 周小勇, 黄忠良, 魏识广, 史军辉. 鼎湖山植物群落α多样性与环境的关系. 生态学报, 2006, 26(7): 2301- 2307.
[18] 吴裕鹏, 许涵, 李意德, 骆土寿, 陈德祥, 林明献. 海南尖峰岭热带林乔灌木层物种多样性沿海拔梯度分布格局. 林业科学, 2013, 49(4): 16- 23.
[19] 蒋国芳. 木论喀斯特森林区生物多样性初报. 生物多样性, 1995, 3(2): 91- 94.
[20] 宋同清, 彭晚霞, 曾馥平, 王克林, 覃文更, 谭卫宁, 刘璐, 杜虎, 鹿士杨. 木论喀斯特峰丛洼地森林群落空间格局及环境解释. 植物生态学报, 2010, 34(3): 298- 308.
[21] 韩文衡, 向悟生, 叶铎, 吕仕洪, 丁涛, 李先琨. 广西木论保护区喀斯特常绿落叶阔叶混交林优势种空间格局及其相关性. 应用生态学报, 2010, 21(11): 2769- 2776.
[22] 彭晚霞, 宋同清, 曾馥平, 王克林, 刘璐. 喀斯特常绿落叶阔叶混交林植被的空间异质性. 西北植物学报, 2011, 31(4): 815- 822.
[23] 宋同清. 西南喀斯特植物与环境. 北京: 科学出版社, 2015:115-123.
[24] 郝占庆, 李步杭, 张健, 王绪高, 叶吉, 姚晓琳. 长白山阔叶红松林样地(CBS): 群落组成与结构. 植物生态学报, 2008, 32(2): 238- 250.
[25] 刘海丰, 李亮, 桑卫国. 东灵山暖温带落叶阔叶次生林动态监测样地: 物种组成与群落结构. 生物多样性, 2011, 19(2): 232- 242.
[26] 祝燕, 赵谷风, 张俪文, 沈国春, 米湘成, 任海保, 于明坚, 陈建华, 陈声文, 方腾, 马克平. 古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构. 植物生态学报, 2008, 32(2): 262- 273.
[27] 叶万辉, 曹洪麟, 黄忠良, 练琚愉, 王志高, 李林, 魏识广, 王章明. 鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究. 植物生态学报, 2008, 32(2): 274- 286.
[28] 兰国玉, 胡跃华, 曹敏, 朱华, 王洪, 周仕顺, 邓晓保, 崔景云, 黄建国, 刘林云, 许海龙, 宋军平, 何有才. 西双版纳热带森林动态监测样地—树种组成与空间分布格局. 植物生态学报, 2008, 32(2): 287- 298.
[29] 郝占庆, 于德永, 杨晓明, 丁之慧. 长白山北坡植物群落α多样性及其随海拔梯度的变化. 应用生态学报, 2002, 13(7): 786- 789.
[30] Vázquez G J A, Givnish T J. Altitudinal gradients in tropical forest composition, structure, and diversity in the Sierra de Manantlán. Journal of Ecology, 1998, 86(6): 999- 1020.
[31] Rahbek C. The role of spatial scale and the perception of large-scale species-richness patterns. Ecology Letters, 2005, 8(2): 224- 239.
[32] 王国宏. 祁连山北坡中段植物群落多样性的垂直分布格局. 生物多样性, 2002, 10(1): 7- 14.
[33] Wang G H, Zhou G S, Yang L M, Li Z Q. Distribution, species diversity and life-form spectra of plant communities along an altitudinal gradient in the northern slopes of Qilianshan Mountains, Gansu, China. Plant Ecology, 2003, 165(2): 169- 181.
[34] 张华, 郑楠, 武晶, 祝业平, 李剑源, 张宝财, 孙卫东. 辽东山地老秃顶子北坡植物物种多样性的垂直分布格局. 辽宁师范大学学报: 自然科学版, 2009, 32(4): 484- 489.
[35] Doležal J,tek M. Altitudinal changes in composition and structure of mountain-temperate vegetation: a case study from the Western Carpathians. Plant Ecology, 2002, 158(2): 201- 221.
[36] 袁道先, 章程. 岩溶动力学的理论探索与实践. 地球学报, 2008, 29(3): 355- 365.
[37] 王宇超, 王得祥. 佛坪自然保护区植物群落物种多样性和复杂性的海拔格局研究. 西北植物学报, 2013, 33(1): 169- 176.
[38] 何艳华, 闫明, 张钦弟, 张直峰, 苗艳明, 毕润成. 五鹿山国家级自然保护区物种多样性海拔格局. 生态学报, 2013, 33(8): 2452- 2462.
[39] Whittaker R J, Willis K J, Field R. Scale and species richness: towards a general, hierarchical theory of species diversity. Journal of Biogeography, 2001, 28(4): 453- 470.
[40] 马克平, 刘灿然, 刘玉明. 生物群落多样性的测度方法——Ⅱβ多样性的测度方法. 生物多样性, 1995, 3(1): 38- 43.
[41] 张璐, 苏志尧, 李镇魁. 南岭国家级自然保护区森林群落β多样性随海拔梯度的变化. 热带亚热带植物学报, 2007, 15(6): 506- 512.
[42] 郝占庆, 于德永, 吴钢, 邓红兵, 姜萍, 王庆礼. 长白山北坡植物群落β多样性分析. 生态学报, 2001, 21(12): 2018- 20
Altitudinal pattern of woody plant species diversity in the karst forest in Mulun, China
LAN Sian1,2,SONG Min1,2,3,ZENG Fuping1,2,SONG Tongqing1,2,*,PENG Wanxia1,2,DU Hu1,2,HAN Chang1,2,4, CHEN Li1,2,4,QIN Wengeng5
1KeyLaboratoryofAgro-ecologicalProcessesinSubtropicalRegion,InstituteofSubtropicalAgriculture,ChineseAcademyofSciences,Changsha410125,China2HuanjiangObservationandResearchStationofKarstEcosystem,ChineseAcademyofSciences,Huanjiang547100,China3CollegeofBioscienceandBiotechnology,HunanAgriculturalUniversity,Changsha410128,China4CollegeofAgriculture,HunanAgriculturalUniversity,Changsha410128,China5AdministrativeBureauofMulunNationalNatureReserve,GuangxiZhuangAutonomousRegion,Huanjiang547200,China
Variation in species diversity along ecological gradients is the main focus in recent biodiversity studies. Among multiple interacting resource gradients, altitude gradient may be a decisive factor determining spatial patterns of species diversity. Forest community classification, ordination, and species diversity patterns along altitudinal gradients can reveal relationships between species distribution and environmental factors, and provide an important basis for understanding the function of mountain ecosystems. For studying the species diversity along ecological gradients in karst ecosystems, we conducted a study in Mulun National Nature Reserve (MNNR), northwest of Huanjiang Maonan Autonomous County in Guangxi, China. This reserve and Maolan National Nature Reserve together possesses the largest and most typical karst forests in the world, and play an important role in plant biodiversity research due to their unique geography, climate, and vegetation conditions. The climax community in this region is an evergreen and deciduous broadleaf mixed forest on limestone in the central subtropical zone in MNNR. The area of this forest is 89.69 km2with 94.8% of vegetation coverage. The soil type is calcareous lithosols (limestone soil). Sixteen sample plots (20 m × 20 m) were established in a typical depression landscape between karst hills on a southwestern slope in MNNR. All woody plants [DBH (diameter at breast height) ≥ 1 cm] from the 16 plots were investigated. We analyzed the pattern of woody plant species diversity along an altitudinal gradient in MNNR by measuring the indices of species richness, diversity, and evenness. The results showed that: (1) there were 5089 individuals in the studied plots, which belonged to 120 species, 48 families and 90 genera; (2) there was a distinct altitudinal pattern of woody plant species in the karst forest communities. Hierarchical cluster analysis and Non-metric Multidimensional Scaling analysis showed that the forest communities could be classified into three types, including those in lower slope, middle slope and upper slope respectively. In the lower slope, the dominant species wereCryptocaryaaustrokweichouensisandCleidionbracteosumare; in the middle slope, the dominant species wereRubovietnamiaaristataandBrassaiopsisglomerulata;PlatycaryastrobilaceaandRapaneaneriifoliaarewere the dominant species in the upper slope. As the altitude increases, the temperature and moisture also changed. Aridity and low temperature respectively limited plant growth at low and high altitude; the precipitation and sunshine were plentiful in middle elevations, which supports a high diversity of plant species; (3) among the woody species, there was a greater proportion of evergreen species than deciduous broadleaf species. In addition, coniferous species only grew on the top of the mountain; (4) Species richness and the Shannon-Wiener index, along the altitudinal gradient, exhibited a pattern of unimodal distribution, with the peak appearing in the mid slope; (5) theβdiversity index fluctuated considerably, and decreased with rising elevation in the ecosystem. Our results will be useful for developing strategies of vegetation restoration in the karst region.
altitudinal gradient;αdiversity;βdiversity; woody species; karst
中国科学院科技服务网络计划STS资助项目(KFJ-EW-STS-092); 国家自然科学基金资助项目(31370485, 31370623, 31400412, 31460135); 广西科技惠民资助项目(桂科转1599001- 6);广西特聘专家资助项目; 湖南省研究生科研创新资助项目(CX2015B231)
2016- 01- 08;
2016- 05- 06
10.5846/stxb201601080055
*通讯作者Corresponding author.E-mail: songtongq@isa.ac.cn
兰斯安, 宋敏, 曾馥平, 宋同清, 彭晚霞, 杜虎, 韩畅, 陈莉, 覃文更.木论喀斯特森林木本植物多样性垂直格局.生态学报,2016,36(22):7374- 7383.
Lan S A,Song M,Zeng F P,Song T Q,Peng W X,Du H,Han C, Chen L,Qin W G.Altitudinal pattern of woody plant species diversity in the karst forest in Mulun, China.Acta Ecologica Sinica,2016,36(22):7374- 7383.
猜你喜欢
林业勘查设计(2022年1期)2022-02-15
现代园艺(2021年23期)2021-12-01
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
林业勘查设计(2020年1期)2021-01-18
新农业(2020年18期)2021-01-07
林业调查规划(2020年3期)2020-06-03
乡村地理(2019年2期)2019-11-16
————水溶蚀岩石的奇观
家教世界(2019年4期)2019-02-26
文化月刊·下旬刊(2014年6期)2014-08-28
