
不同动物高免血清对猪肺炎支原体的代谢抑制作用研究
2016-02-07 08:22:41孙亚波魏晶晶刘富东王芳宁宜宝孙晔王凤霞沈青春
中国兽药杂志 2016年12期
孙亚波,魏晶晶,刘富东,王芳,宁宜宝,孙晔,,王凤霞,沈青春,*
(1.北京中海生物科技有限公司,北京 100081;2.中国兽医药品监察所,北京 100081)
不同动物高免血清对猪肺炎支原体的代谢抑制作用研究
孙亚波1,魏晶晶1,刘富东1,王芳2,宁宜宝2,孙晔1,2,王凤霞1,沈青春1,2*
(1.北京中海生物科技有限公司,北京 100081;2.中国兽医药品监察所,北京 100081)
选用猪、兔、鸡、小鼠和豚鼠5种试验动物分别制备猪肺炎支原体高免阳性血清,并测定了其对猪肺炎支原体的代谢抑制效价。再从兔高免血清中纯化特异性抗体,比较抗体纯化前后的代谢抑制效价是否发生变化。结果表明,5种试验动物高免血清对猪肺炎支原体的代谢抑制价分别为0、104、103、105、102,纯化后的抗体仍可抑制猪肺炎支原体的生长,代谢抑制价没有明显变化。由此可见,不同动物来源的高免血清对猪肺炎支原体的代谢抑制价不同,代谢抑制作用的主要作用物质为特异性抗体;首次证明病原靶动物——猪的高免抗血清对猪肺炎支原体没有代谢抑制作用,其他支原体是否也表现为对靶动物(或人)抗血清的耐受性还有待进一步研究。
猪肺炎支原体;代谢抑制作用;高免血清
猪肺炎支原体(Mycoplasmahyopneumoniae,Mhp)是引起猪地方流行性肺炎(Swine enzootic pneumoniae,SEP)的病原,国内称为“猪喘气病”。该病广泛流行于世界各地,是严重危害全球养猪业健康发展的慢性疾病之一[1]。
支原体具有类似于病毒中和作用的代谢抑制(Metabolic inhibition, MI)特性,是细菌所不具有的特殊性质[2]。该特性具有很强的种属特异性,即抗一种支原体的高免血清只能抑制该种支原体的生长繁殖,而对其他种类的支原体不具有抑制作用,因而以该特性为基础的代谢抑制试验和生长抑制试验已成为支原体鉴定的金标准[3-5]。本实验室研究工作中发现,猪抗Mhp的高免血清对Mhp似乎不具有代谢抑制作用,而兔抗Mhp的高免血清则抑制作用较强,有关不同动物的抗血清对支原体的代谢抑制作用的相关问题,在国内外还没有任何报道,甚至代谢抑制相关研究也仅有早期的少量报道。为了调查几种常见试验动物抗Mhp高免血清的代谢抑制作用情况,本实验选用了5种试验动物进行了免疫和代谢抑制效价测定及相关研究。
1 材料和方法
1.1 材料
1.1.1 菌毒种 猪肺炎支原体232株,来自中国兽医药品监察所。
1.1.2 试剂和培养基 PPLO、猪血清、马血清等,购自GIBCO公司;Lps-5支原体液体培养基(含PPLO、葡萄糖、MEM等)和猪肺炎支原体间接血凝(IHA)抗原(批号:201501),北京中海生物科技有限公司自制。IMS 1313水性佐剂,法国赛比克公司提供。
1.1.3 试验动物 BalB/C小鼠、豚鼠和清洁级健康家兔,来自北京维通利华实验动物技术有限公司。SPF鸡购自梅里亚。30~40日龄健康仔猪,购自保定某商品代猪场,猪肺炎支原体抗原、抗体阴性,猪瘟、猪伪狂犬、猪蓝耳病抗原阴性。
1.2 抗原的制备 开启猪肺炎支原体232株冻干菌种,使用Lps-5支原体液体培养基,按1/10的接种比例扩大繁殖至5000 mL菌液,8000 r/min离心30 min后,使用PBS(pH 7.2±0.1)100 mL重悬,灭活后1313水性佐剂100 mL,低剪切力混合5~10 min,按照现行《中国兽药典》附录要求进行无菌检验后,置2~8 ℃避光保存备用。
1.3 阳性血清的制备与检测 BalB/C小鼠、豚鼠、家兔、SPF鸡和仔猪5种动物的数量分别为10只、5只、3只、3只和2头,单次免疫剂量分别为0.2 mL/只、0.5 mL/只、1 mL/只、1 mL/只和2 mL/头,首次免疫14日后,每隔一周使用相同剂量加强免疫一次,5次免疫后10日采血分离血清,进行琼扩效价和IHA效价测定。
1.4 活菌滴度测定 取新鲜培养的猪肺炎支原体232株培养物,按照CCU50测定方法进行活菌浓度测定[6],用于代谢抑制试验的培养物CCU50滴度应不低于10-8.0/mL,置-70 ℃以下冻存不超过3个月。
2.活跃课堂气氛,改变陈旧的教学理念和模式,培养学生自觉思维能力。特别在课堂上要形成师生之间、生生之间的彼此交流、互相质疑,让学生的创新意识、创新思维能够互相影响、互相激励,产生链式反应。
1.5 代谢抑制试验 使用Lps-5支原体液体培养基对经CCU50测定的Mhp 232株培养物进行10倍比稀释至10-8,在10-6、10-7、10-8三个稀释度各取1 mL,向其中分别加入1 mL含10%待测抗Mhp阳性血清的液体培养基,充分混匀,取10-6、10-7、10-8三个稀释度各2 mL作为对照管。另设2 mL液体培养基作空白对照组,于37℃培养14日后判定结果。空白对照不变色,对照组3个滴度都变黄,试验组至少10-7、10-8两个滴度不变色即可判定为猪肺炎支原体可被待测阳性血清所抑制。
1.6 代谢抑制效价测定 采用1.5的方法将Mhp 232株培养物10倍比稀释至10-8,取10-1~10-8七个稀释度各取1 mL,向其中分别加入1 mL含10%待测抗Mhp阳性血清的液体培养基,充分混匀,取10-6、10-7、10-8三个稀释度各2 mL作为对照。另设2 mL液体培养基作空白对照组,于37 ℃培养14 d后判定结果。空白对照不变色,3支对照管都变黄,记录试验组最后一支变色小管的滴度,并计算出待测阳性血清的代谢抑制价,如10-1~10-3小管变色,10-4~10-7小管不变色,而测定菌液的CCU为108/mL,则共抑制了5个滴度,即待测阳性血清的代谢抑制价为105。
1.7 代谢抑制作用的机理初步研究
1.7.1 猪高免阳性血清中加入兔阴性血清后的代谢抑制价测定 将兔阴性血清与猪抗Mhp阳性血清按1∶1混合,再按照1.5项的方法进行代谢抑制试验,以确定兔血清中是否存在某种特殊因子有助于抗体对Mhp的代谢抑制作用。
1.7.2 纯化兔特异性抗体的代谢抑制效价测定 家兔高免血清表现出较强的代谢抑制作用,为了确定代谢抑制作用是由特异性抗体的结合引起,还是与其他因子共同作用的结果,本试验将20 mL家兔高免血清经硫酸铵沉淀后用PBS复溶,经分子筛过柱后,获得纯化抗体10 mL(浓度10.4 mg/mL)。将原倍和倍比稀释物分别替代高免血清按1.6项方法进行代谢抑制效价测定。
2 结 果
2.1 抗原制备 用于抗原制备的猪肺炎支原体232株培养物共计5000 mL,取10 mL分装成1 mL/支小管,共10支,置-70 ℃以下冻存,用于代谢抑制试验。其余培养物用于制备抗原,共获得抗原195 mL,无菌检验结果均符合规定。培养物的CCU50测定结果为1.72 × 108CCU50/mL。
2.2 阳性血清的制备与检测结果 对制备的5种动物的Mhp高免阳性血清进行琼扩效价测定,同时按照现行《中国兽药典》进行猪支原体肺炎间接血凝试验,结果表明5种高免血清的效价均高于1∶10,即均为猪肺炎支原体阳性,其IHA效价和琼扩效价见表1。
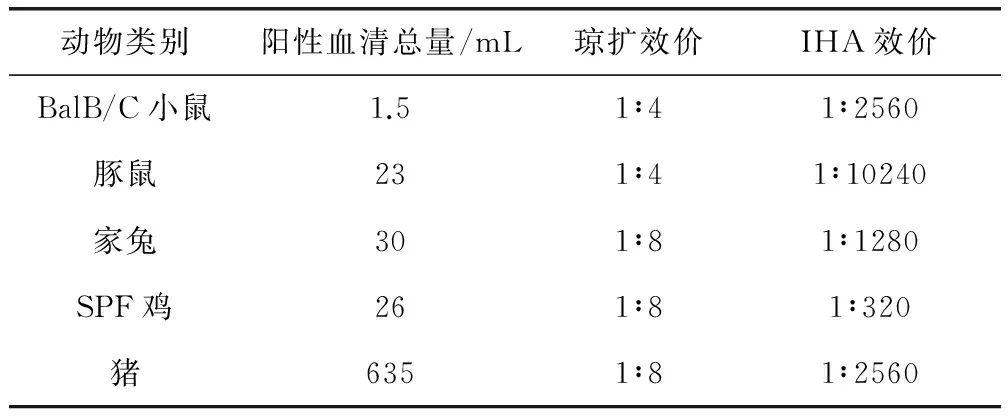
表1 5种动物的高免阳性血清制备结果
2.3 代谢抑制试验 代谢抑制试验结果表明BalB/C小鼠、豚鼠、家兔和SPF鸡的高免阳性血清都可以不同程度地抑制猪肺炎支原体的生长,而仔猪高免血清的代谢抑制结果为三个稀释度为均不变色,表现为不抑制,结果见表2。

表2 5种动物的高免阳性血清代谢抑制试验结果
2.4 代谢抑制价的测定 为进一步确定上述各动物高免血清的代谢抑制能力,对其分别进行代谢抑制价测定,结果见表3。
结果表明,小鼠的高免血清具有很强的代谢抑制特性,代谢抑制价最高,达到105,其次是家兔,可达104,IHA抗体效价最高的豚鼠阳性血清的代谢抑制价则比较低,而IHA抗体效价次高的仔猪的代谢抑制价结果为0,即无抑制。

表3 5种动物的高免阳性血清代谢抑制价测定结果
上述结果表明,不同动物的高免阳性血清对Mhp的代谢抑制能力不同,琼扩效价和IHA效价均不能很好地反映血清的代谢抑制能力,而本动物——猪高免血清没有表现出代谢抑制能力。出现上述结果是否与支原体的免疫逃避机制[7-8]有关需要进一步的研究。
2.5 代谢抑制作用的机理初步研究
2.5.1 混合血清的代谢抑制效价测定 家兔高免血清表现出较强的代谢抑制作用,而猪的高免血清尽管琼扩效价和IHA效价均高,但代谢抑制价测定结果为0,是否说明代谢抑制作用不仅仅与抗体有关,还与存在于兔血清中的其他因子参与其中,本实验将猪阳性血清等比例混合兔阴性血清后进行代谢抑制价测定,结果见表4。

表4 猪、兔混合血清的代谢抑制价测定结果
结果表明,兔阴性血清并没有改变仔猪高免阳性血清的代谢抑制效价,由此可见兔阴性血清中不含与代谢抑制有关的成分。2.6 纯化的兔特异性抗体的代谢抑制效价测定 本试验共获得纯化抗体10 mL(浓度10.4 mg/mL)。将原倍和倍比稀释物的纯化家兔特异性抗体分别替代高免血清按1.6项方法进行代谢抑制效价测定,结果见表5。

表5 兔纯化抗体的代谢抑制价测定结果
结果表明,兔抗猪肺炎支原体抗体可独立抑制猪肺炎支原体的生长,抑制作用的强弱与抗体浓度直接相关,抗体浓度降低一半后,代谢抑制作用下降了2个滴度,由此可见二者似乎不呈线性关系。
3 讨论与结论
支原体是自然中能独立生长繁殖的最小生物,其结构简单,无细胞壁。其代谢抑制特性是在人类发现病毒的抗体中和作用之后发现的,美国科学家1958年首次发现用柯赛基病毒B组抗体可抑制该病毒的感染,即病毒中和作用[9]。1966年,Purcell R H等首次发现兔抗支原体高免血清对相应支原体具有显著抑制其生长繁殖的作用[2,10]。
本研究需要对不同动物高免血清对支原体的代谢抑制作用的强弱进行评价,首次引入了“代谢抑制效价”的概念及其测定方法,而此前国内外还没有任何支原体抗血清的代谢抑制强弱及其评价方法的报道,实验证明“代谢抑制效价”能很好地评价各血清样品的代谢抑制作用强弱,而且测定方法较为简单。一直以来国内外用于代谢抑制试验所用的抗血清多采用兔高免血清,而这可能与家兔容易饲养,并能得到足够量的血清有关,本研究结果表明在所制备的猪肺炎支原体的高免血清中,鼠高免血清相比兔具有更高的代谢抑制效价,但相比家兔,小鼠血清制备较为困难,单只所得血清量少。
病毒的中和试验基本原理是特异性抗体主要通过封闭病毒与细胞的结合受体使其失去感染性[11]。支原体的代谢抑制特性是否与病毒的中和特性相似?本研究采用了纯化后的兔源特异性抗体进行试验,发现其可以抑制猪肺炎支原体的生长,但为何猪的高免血清对Mhp又不具有代谢抑制作用?是因为不同动物抗体在种属特异性结构上的差异,还是其他原因造成这样的结果,还需要进一步的实验研究予以解答。
Mhp对其靶动物——猪的抗Mhp高免血清几乎没有代谢抑制作用的结论,就引出了一个新问题,猪支原体肺炎当前最主要的防治手段是免疫预防,即接种弱毒活疫苗和灭活疫苗,弱毒活疫苗能产生细胞免疫和体液免疫,获得很好的免疫保护力容易理解,而灭活疫苗则主要产生体液免疫,即产生针对Mhp的抗体。病毒疫苗的体液免疫主要通过中和作用实现抗感染,而从本文结论可知猪支原体肺炎疫苗似乎不太可能是通过特异性抗体的代谢抑制作用实现抗感染。Mhp灭活疫苗是当前全球用于预防猪支原体肺炎最主要的工具之一,事实证明可以使猪体产生良好的免疫保护力[12-13],其产生抗感染的作用的产生可能与代谢抑制没有直接关系,而与细菌疫苗的抗感染作用相似。
猪阳性血清对Mhp不具有代谢抑制作用,将对Mhp的培养具有很好的应用参考价值,一些文献要求配制用于培养猪肺炎支原体的培养基所使用的猪血清为Mhp抗体阴性[14],而Mhp抗体阴性的成年猪极难找到,得到大量阴性血清非常困难[15]。本研究的结果显示试验Mhp抗体阳性的猪血清对Mhp不具有代谢抑制作用,因而不影响Mhp的生长繁殖,即用于Mhp培养的猪血清无需要求Mhp抗体阴性。
本研究首次报到了Mhp的靶动物——猪的抗Mhp高免血清对Mhp不具有代谢抑制作用,兔抗Mhp特异性抗体可独立抑制Mhp的生长。其他支原体是否也对靶动物(或人)的高免抗血清具有耐受性,该特性是否与代谢抑制作用的机理有关,还有待进一步的研究。
[1] 沈青春, 宁宜宝, 覃青松, 猪肺炎支原体的研究进展[J]. 中国兽药杂志,2003, 37(6):26-30.
[2] Purcell R H, Wong D, Chanock R M,etal, Significance of antibody to mycoplasma as measured by metabolic-inhibition techniques[J]. Ann N Y Acad Sci,1967,143:664-675.
[3] Poveda J B, Nicholas R. Serological identification of mycoplasmas by growth and metabolic inhibition tests[J]. Methods Mol Biol,1998,104:105-111.
[4] Poveda J B. Biochemical characteristics in mycoplasma identification[J]. Methods Mol Biol,1998,104:69-78.
[5] Assuncao P, Diaz R, Comas J,etal. Evaluation ofMycoplasmahyopneumoniaegrowth by flow cytometry[J]. J Appl Microbiol,2005,98:1047-1054.
[6] 沈青春, 李聪研, 冯倩倩, 等.用半数变色单位法精确测定支原体活菌滴度[J]. 微生物学报,2013, 53(12):1347-1352.
[7] Bogema D R, Deutscher A T, Woolley L K,etal. Characterization of cleavage events in the multifunctional cilium adhesin Mhp684 (P146) reveals a mechanism by whichMycoplasmahyopneumoniaeregulates surface topography[J]. mBio,2012, 3(2):1-11.
[8] Deutscher A T, Jenkins C, Minion F C,etal. Repeat regions R1 and R2 in the P97 paralogue Mhp271 ofMycoplasmahyopneumoniaebind heparin, fibronectin and porcine cilia[J]. Mol Microbiol,2010,78:444-458.
[9] Rogers N G, Bankhead A S, Crawford I P,etal. Metabolic inhibition test for determination of antibodies to group B Coxsackie viruses[J]. Proc Soc Exp Biol Med,1958,98:227-231.
[10]Purcell R H, Taylor-Robinson D, Wong D C,etal. A color test for the measurement of antibody to the non-acid-forming humanMycoplasmaspecies[J]. Am J Epidemiol,1966,84:51-66.
[11]Tebruegge M, Curtis N. Enterovirus infections in neonates[J]. Semin Fetal Neonatal Med,2009,14:222-227.
[12]Wilson S, Van Brussel L, Saunders G,etal. Vaccination of piglets at 1 week of age with an inactivatedMycoplasmahyopneumoniaevaccine reduces lung lesions and improves average daily gain in body weight[J]. Vaccine,2012, 30(52):7625-7629.
[13]Simionatto S, Marchioro S B, Maes D,etal.Mycoplasmahyopneumoniae: from disease to vaccine development[J]. Vet Microbiol,2013,165:234-242.
[14]Marois C, Le Carrou J, Kobisch M,etal. Isolation ofMycoplasmahyopneumoniaefrom different sampling sites in experimentally infected and contact SPF piglets[J]. Vet Microbiol,2007,120:96-104.
[15]Xiong Q, Wei Y, Feng Z,etal. Protective efficacy of a live attenuatedMycoplasmahyopneumoniaevaccine with an ISCOM-matrix adjuvant in pigs[J]. Vet J,2014,199:268-274.
(编辑:李文平)
Metabolic Inhibition Studies on Different Animal Hyperimmune Antiserum againstMycoplasmahyopneumoniae
SUN Ya-bo1,WEI Jing-jing1,LIU Fu-dong1,WANG Fang2,NING Yi-bao2,SUN Ye1,2,WANG Feng-xia1, SHEN Qing-chun1,2*
(1.BeijingZhonghaiBiotechCo.Ltd.,Beijing100081,China; 2.ChinaInstituteofVeterinaryDrugControl,Beijing100081,China)
Five kinds of experimental animals, including pig, rabbit, chicken, mice and guinea pig were immuned repeatedly to prepare hyperimmune serum againstMycoplasmahyopneumoniae(Mhp), and the metabolic inhibition potency of the sera were determined. The purified specific antibody from rabbit hyperimmune sera was used for the potency determination,too.The results showed that the metabolic inhibitory potency to Mhp of the hyperimmune sera from five kinds of animals were 0,104,103,105and 102, respectively.The purified specific antibody could inhibit the growth of Mhp, and the potency was 104. Therefore, the metabolic inhibitory potency against Mhp of the hyperimmune serum from different animals was different. The main effective material of metabolic inhibition was the specific antibody. The agent target animal —— pig’s hyperimmune antiserum against Mhp could not inhibit the growth of Mhp. Whether or not the other mycoplasma has the tolerance to antiserum from target animals (or human) needs to be further studied.
Mycoplasmahyopneumoniae; metabolic inhibition; hyperimmune antiserum
孙亚波,本科,从事兽用生物制品的研制工作。
2016-09-27
A
1002-1280 (2016) 12-0007-05
S858.28
猜你喜欢
心理学探新(2022年1期)2022-06-07 09:16:02
今日农业(2021年17期)2021-11-26 23:38:44
兽医导刊(2019年1期)2019-02-21 01:14:26
兽医导刊(2019年1期)2019-02-21 01:13:58
猪业科学(2018年5期)2018-07-17 05:55:18
妈妈宝宝(2017年3期)2017-02-21 01:22:14
兽医导刊(2016年12期)2016-05-17 03:51:45
兽医导刊(2015年9期)2016-01-04 12:00:06
兽医导刊(2015年7期)2016-01-04 11:59:58
中国当代医药(2015年8期)2015-03-01 02:01:55
