
低温层积对纳罗克非洲狗尾草种子酶活性及其萌发的影响
2016-02-05 02:56:18张桓溥罗富成杜凤麒李博伦
草原与草坪 2016年6期
张桓溥,彭 健,罗富成,韩 博,何 超,杜凤麒,李博伦
(1.云南农业大学 动物科学技术学院草业科学系,云南 昆明 650201; 2.贵州省六盘水市烟草专卖局(公司),贵州 六盘水 553000)
低温层积对纳罗克非洲狗尾草种子酶活性及其萌发的影响
张桓溥1,彭 健2,罗富成1,韩 博1,何 超1,杜凤麒1,李博伦1
(1.云南农业大学 动物科学技术学院草业科学系,云南 昆明 650201; 2.贵州省六盘水市烟草专卖局(公司),贵州 六盘水 553000)
为探究低温层积对纳罗克非洲狗尾草(Setaria sphacelata cv.Narok)种子酶活性及其萌发的影响,将一年龄纳罗克非洲狗尾草生产的种子进行低温(4℃)层积不同时间处理,并测定在不同处理下种子的酶活性、发芽率及活力水平。结果表明:纳罗克非洲狗尾草种子在低温层积处理最佳时间为30 d,在此处理条件下,种子休眠已完全解除,此时种子的发芽率和各项活力指数分别达到75%和0.0263,是对照(低温层积处理0d)的2.08倍和2.27倍,差异达到极显著水平(P<0.01);种子超氧化物歧化酶(SOD)活性,呈先下降后升高并逐渐稳定的趋势,低温层积 60 d降至最低水平650.800 U/g FW·h,较对照降低了9.07倍;过氧化物酶(POD)活性随时间的推移波动变化,但变化不明显(P>0.05);过氧化氢酶(CAT)活性在层积处理90 d前无显著变化(P>0.05),之后逐渐下降。
纳罗克非洲狗尾草种子;低温层积;酶活性;种子活力
纳罗克非洲狗尾草(Setariasphacelatacv.Narok)原产于肯尼亚的阿尔德地区,1963年引入澳大利亚昆士兰州,1969年通过澳大利亚牧草品种审定,1983年从澳大利亚引入我国云南省,并于1997年通过国家品种审定委员会审定登记为“引进品种”。纳罗克已广泛应用于世界热带和亚热带地区的草地建设、公路护坡、岩溶石漠化治理及矿区植被恢复[1-5],具有牧草产量高[6]、品质佳、水土保持效果好等优点。但其种子产量低,休眠程度深,严重制约了在我国的推广应用[7]。大面积生产条件下,其种子产量在45.0~60.0 kg/hm2,发芽率仅为10%[8],休眠率高达34%~50%[9];致使南方地区纳罗克非洲狗尾草种子供不应求。低温层积是常用的一种模拟自然条件打破种子休眠的方法[10],具有软化种皮,增强种子透性,降低种子ABA 含量,增加GA含量及其对GA的敏感性等优点,进而诱导休眠种子萌发[11]。超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)具有控制植物体内活性氧积累,防御活性氧毒害具有重要作用[12]。
1 材料和方法
1.1 试验材料
材料为云南农业大学草学基地种植第1年(即一年龄植株)收获的纳罗克非洲狗尾草净种子。其发芽率为36%,千粒质量为1.318 3 g、生活力为78.3%、休眠率为42.3%。
1.2 种子低温层积处理
分别取适量的净种子,用1.5%硫酸铜溶液浸种10 min消毒,用清水反复冲洗后与高温灭菌的湿沙(手握成团、松手即散)混匀,装入磁盘,表层再覆盖1 cm厚的湿沙,置于4℃ 冰箱内贮藏(即低温层积)。视干湿情况,每隔3 d喷水1 次,确保湿度。分别于层积后0(CK)、15、30、45、60、75、90、105、120 d随机取出部分种子,洗净晾干后,检测其发芽率、活力及抗氧化酶活性。
1.3 种子发芽率及活力水平测定
参考云南省地方标准“牧草种子检验章程”[13],采用纸上发芽法,将种子置于(30±1)℃,24 h光照的人工气候箱中发芽,3 次重复,每重复100粒。逐日统计发芽粒数,并将发霉种子挑出,第7 d统计发芽势,第14 d统计发芽率。发芽试验结束后,测定幼苗干重,并计算种子发芽指数和活力指数[14]。
1.4 种子酶活性测定
参照文献[15]提供的方法测定种子SOD和POD活性;参照文献[16]提供的方法测定种子CAT活性。
1.5 数据处理
数据整理和作图用Excel完成,用SPSS 17.0软件进行数据方差分析。
2 结果与分析
2.1 低温层积对纳罗克非洲狗尾草种子酶活性的影响
2.1.1 低温层积对纳罗克非洲狗尾草种子SOD活性的影响 经低温层积,纳罗克非洲狗尾草种子的SOD活性随处理时间的延长表现为先下降后升高并逐渐稳定的趋势。层积30 d后SOD活性开始下降(P<0.05),并于层积60 d后降至最低水平,仅为650.800 U/(g FW·h),较对照降低了9.07倍;层积75 d后SOD活性开始有所回升,并在层积90 d后稳定在4 100.000 U/(g FW·h)(图1)。
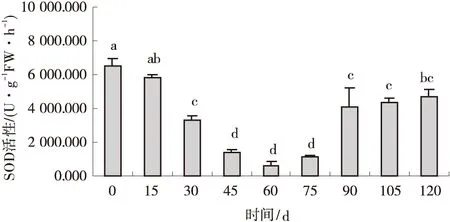
图1 低温层积下纳罗克非洲狗尾草种子的SOD活性Fig.1 Effect of cold stratification treatment on SOD activity of Narok seeds注:不同小写字母表示不同层积时间之间差异显著(P<0.05),下同
2.1.2 低温层积对纳罗克非洲狗尾草种子POD活性的影响 经低温层积后,纳罗克非洲狗尾草种子的POD活性变化不明显(P>0.05),稳定在约6.500 U/(gFW·h)(图2)。
2.1.3 低温层积对纳罗克非洲狗尾草种子的CAT活性影响 纳罗克非洲狗尾草种子的CAT活性在其层积90 d内无显著变化(P>0.05),90 d后开始下降,至120 d降至最低,仅为41.94 U/(g·min),较对照降低了2.66倍(P<0.05)(图3)。

图2 低温层积下纳罗克非洲狗尾草种子的POD活性Fig.2 Effect of cold stratification treatment on POD activity of Narok seeds
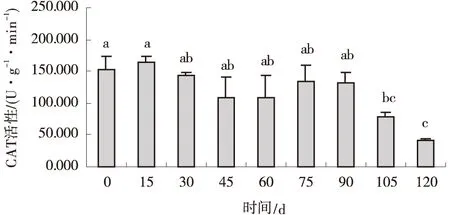
图3 低温层积下纳罗克非洲狗尾草种子的CAT活性Fig.3 Effect of cold stratification treatment on CAT activity of Narok seeds
2.2 低温层积对纳罗克非洲狗尾草种子发芽率的影响
低温层积处理后,纳罗克非洲狗尾草种子的发芽率随层积时间的推移表现为先快速增加,后缓慢下降的趋势。低温层积15 d,种子的发芽率达到73%,显著高于对照(低温层积0 d)(P<0.05),较种子本身的生活力(78.3%)已无显著差异(P>0.05),休眠已基本解除(图4);低温层积30 d后其发芽率超过75%,种子休眠已完全解除。但随着层积时间的进一步延长,种子的发芽率又呈现出下降的趋势,层积第60 d,种子的发芽率较层积第15~30 d已有显著降低(P<0.05),至层积第120 d 发芽率仅为40%,较对照几无差异(P>0.05)(表1)。
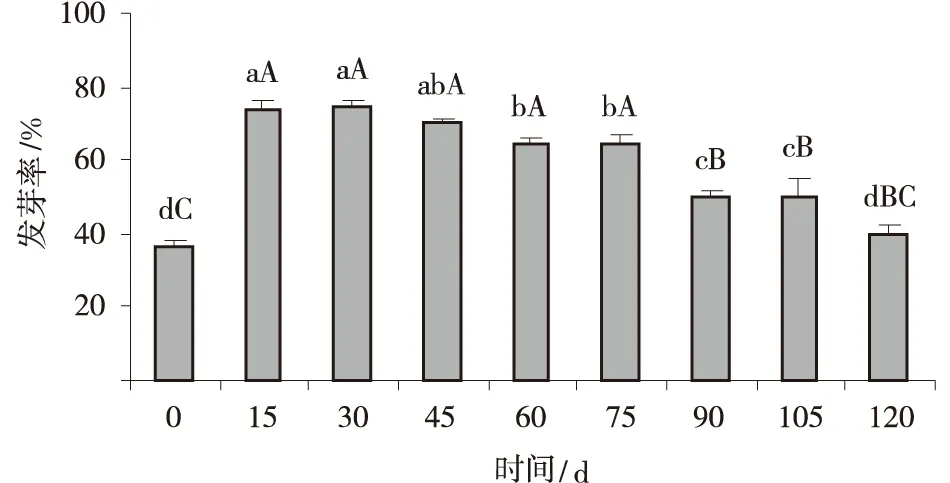
图4 低温层积下纳罗克非洲狗尾草种子的发芽率Fig.4 Effect of cold stratification treatment on seeds germination rate of Setaria sphacelata cv.Narok seeds注:不同小写字母表示不同层积时间之间差异显著(P<0.05),不同大写字母表示差异极显著(P<0.01),下同
2.3 低温层积对纳罗克非洲狗尾草种子活力的影响
随着层积时间的延长,纳罗克非洲狗尾草种子的各项活力指标(包括发芽势、发芽指数和活力指数)与发芽率的变化趋势一致,也均表现出先增加后下降的趋势。低温层积30 d,种子的发芽势、发芽指数和活力指数均达到最高值,分别为74%,23.15和0.026 3,分别是对照的2.55,2.35和2.27倍(P<0.01)。其中,发芽势与层积15 d、45 d没有显著差异(P>0.05),但与其他层积时期均有显著差异(P<0.01);发芽指数与层积15 d、45 d、60 d、75 d没有显著差异(P>0.05),但与其他层积时期均有显著差异(P<0.01);活力指数较层积15 d差异显著(P<0.01),较层积45、60、75 d差异不显著(P>0.05),说明层积30~75 d,种子活力处于高峰期;随后各项活力指标呈下降趋势,至120 d降至最低,分别为38%、9.73 和 0.010 5,此时发芽指数与活力指数与对照的水平相当(P>0.05),但发芽势仍显著高于对照(P<0.01)。

表1 低温层积处理下纳罗克非洲狗尾草种子活力
3 讨论
SOD的作用是将植物细胞内产生的超氧化物O2-歧化为H2O2和O2,使植物免受活性氧的毒害。研究结果表明,纳罗克非洲狗尾草种子SOD酶活性的变化恰好与种子活力水平的变化趋势相反,种子活力伴随着SOD酶活性的降低而升高,SOD酶活性与植物所处环境及自身因素密切相关,处于逆境时活性氧含量较高,SOD酶活性较高[17],故纳罗克非洲狗尾草种子休眠未解除前其内部生理生化反应受到抑制,产生大量活性氧,低温层积处理后,抑制被解除,活性氧含量下降,种子活力上升。这与前人[22-24]的低温层积伴随种子内源激素和酶活性的变化,有利于抑制物质的降解、大分子物质转化为小分子物质,进而解除种子休眠,提高种子的发芽率和生活力的研究结果相吻合。SOD的反应产物H2O2在POD和CAT的共同催化下,歧化为O2和H2O[17]。试验表明,POD的活性呈波动变化但变化不明显(P>0.05);CAT活性前期变化不明显,90 d后则呈现下降趋势,说明低温层积后期种子活力伴随CAT活性的降低而降低,这可能与种子休眠已解除却不能获得萌发的基本条件,种子发生劣变导致种子内部生理代谢受到抑制有关[18-20]。
在120 d的低温层积过程中,纳罗克非洲狗尾草种子的发芽率与活力随低温层积时间的延长均表现为先增加后降低的趋势。起初,发芽率与活力水平快速增加,这可能与层积处理可使种皮物理结构发生变化,提高种皮透性,促使种胚完成后熟有关。试验表明,种子的发芽率及各项活力指标均于种子层积30 d后达到最大值,此时种子发芽率为75%,已接近种子本身的生活力水平(78.3%,P<0.05)。80% 的具有生活力的种子可萌发则视为种子休眠已解除[21-22],故低温层积30 d纳罗克非洲狗尾草种子的休眠已完全解除。
4 结论
纳罗克非洲狗尾草种子低温(4℃)层积的最佳时间是30 d。低温层积30 d,种子休眠已完全解除,其发芽率和活力水平分别是对照的2.08和2.27倍。在低温层积过程中,SOD活性的高低对种子萌发及其活力表达均具有较高的决定作用。SOD活性降低,种子休眠得以解除,活力水平得以提升。
[1] 全国牧草品种审定委员会.中国牧草登记品种集[M].北京:中国农业大学出版社,1999.
[2] Mizumachi S,Shinjo T,Kawamoto Y.Effect of harvesting interval on dry matter productivity of guineagrass(Panicummaximum) and setaria(Setariasphacelata)[J].Japanese Journal of Grassland Science,2009,55(3):199-205.
[3] Mushtaque M,Ishaque M,Bakhush M A A H A.Growth and herbage yield ofSetariasphacelatagrass in response to varying clipping stages[J].The Journal of Animal and Plant Science,2010,20(4):261-265.
[4] Jones R J.Liveweight gain from rotationally and continuously grazed pastures of Narok setaria and Samford rhodesgrass fertilized with-nitrogen in southeast Queensland[J].Tropical grasslands,2004,38(3):135-138.
[5] Ivory D A,Whiteman P C.Effects of environmental and plant factors on foliar freezing resistance in tropical grasses[J].Australian Journal of Agricultural Research,2004,41(2):243-259.
[6] 袁福锦,奎嘉祥,谢有标,等.云南南亚热带多雨区引进禾本科牧草的适应性及评价[J].草原与草坪,2005(6):21-24.
[7] 崔阁英,邓菊芬,尹俊.不同脱粒方法对纳罗克非洲狗尾草种子产量和质量的影响[J].云南农业大学学报,2009,24(3):369-373.
[8] 何超,郭轶敏,彭健,等.稀土溶液浸种对纳罗克非洲狗尾草种子休眠的破除效果[J].种子,2014,33(4):18-20.
[9] 罗富成,郭轶敏,彭健,等.外源激素对纳罗克非洲狗尾草种子休眠的破除效果[J].草业科学,2015,33(3):406-412.
[10] 段春华,张德罡,张靖,等.物理、化学处理方法对矮生嵩草和线叶嵩草种子萌发的影响[J].草原与草坪,2013,33(1):40-44.
[11] 金正律,洪权春.低温层积对决明子种子发芽及幼苗生长的影响[J].种子,2012,31(3):107-109.
[12] 高俊杰,秦爱国,于贤昌.低温胁迫下嫁接对黄瓜叶片SOD和CAT基因表达与活性变化的影响[J].应用生态学报,2009,20(1):213-217.
[13] 云南省技术监督局.DB/5300B20001-91牧草种子检验规程[S].昆明:1991.
[14] 颜启传.种子学[M].北京:中国农业出版社,2001:420-433.
[15] 孙群,胡景江.植物生理学研究技术[M].杨凌:西北农林科技大学出版社,2005.
[16] 蔡庆生.植物生理学实验[M].北京:中国农业大学出版社,2013.
[17] 李璇,岳红,王升,等.影响植物抗氧化酶活性的因素及其研究热点和现状[J].中国中药杂志,2013,38(7):973-978.
[18] Chin H L,Ling Y L,Meno J T.The Effect of Stratification and Thidiazuron Treatment on Germination and Protein Synthesis ofPyrusserotinaRehd cv.Niauli[J].Annals of Botany,1994,73:515-523.
[19] Susan E,Meyer,Rosemary L,Pendleton.Genetic Regulation of Seed Dormancy in Purshia tridentate(Rosaceae)[J].Annals of Botany,2000,85:521-529.
[20] 马天晓,王艳梅,李世荣,等.硫酸和低温处理对酸角种子发芽率的影响[J].河南农业大学学报,2012,46(2):152-155.
[21] 于天祥,段乃斌,王建成,等.低温储藏对高羊茅种子萌发特性的影响[J].山东农业科学,2009(9):86-88.
[22] 叶常丰,戴心维.种子学[M].北京:中国农业出版社,1994:180-185.
Effect of cold stratification on seed germination and enzyme activity of Setaria sphacelata cv.Narok
ZHANG Huan-pu1,PENG Jian2,LUO Fu-cheng1,HAN Bo1,HE Chao1, DU Feng-qi1,LI Bo-lun1,
(1.DepartmentofGrasslandScience,CollegeofAnimalScienceandTechnology,YunnanAgriculturalUniversity,Kunming650201,China; 2.TobaccocompanyofLiupanshui,Liupanshui553000,China)
Setariasphacelatacv.Narok is one of excellent tropical and subtropical forages.However,the deep seeds dormancy restricts the popularization and utilization of the forage.Used one year oldSetariasphacelatacv.Narok under cold stratification treatment and determined its enzyme activity,germination rate and vigor,the results showed that after the cold stratification 30 d,seed dormancy has been completely removed,the seed germination rate and vigor index were 75% and 0.026 3,respectively,which were 2.08 times and 2.27 times compared with the control and had a significant difference (P<0.01).The SOD activity of seed decreased and then increased to the stable level.After cold stratification 60 d,the SOD activity reached lowest and reduced 9.07 times compared with the control.The POD activity fluctuated along with time change,but the change was not obvious (P>0.05).The CAT activity had no changes before cold stratification 90 d (P>0.05)and then decreased. Key words:one year oldSetariasphacelatacv.Narok;cold stratification;seeds vigor;enzyme activity
2016-01-18;
2016-07-07
国家自然科学基金项目“纳罗克非洲狗尾草种子休眠机理及解除方法研究”(31260582)资助
张桓溥(1992-),男,吉林白山人,在读硕士。 E-mail:854623468@qq.com 罗富成为通讯作者。
S 544
A
1009-5500(2016)06-0035-05
猜你喜欢
植物资源与环境学报(2022年6期)2022-12-16 11:35:12
防护林科技(2020年11期)2020-12-30 03:55:36
潍坊学院学报(2020年6期)2020-11-22 08:04:08
作文周刊·小学一年级版(2020年40期)2020-10-19 04:42:20
中国食品学报(2019年3期)2019-01-13 01:47:39
许昌学院学报(2018年8期)2018-09-05 02:05:18
小学生作文选刊·低年级版(2016年6期)2016-06-24 08:40:46
蔬菜(2016年10期)2016-03-27 12:35:11
武夷学院学报(2015年3期)2015-07-18 11:03:47
云南中医学院学报(2014年3期)2014-07-31 18:58:03
