
近交对凡纳滨对虾(Litopenaeus vannamei)生长、存活及抗逆性的影响*
2016-01-15 11:22:59袁瑞鹏刘建勇张嘉晨陈晓敏郑静静刘加慧
海洋与湖沼 2016年6期
袁瑞鹏 刘建勇① 张嘉晨 陈晓敏 郑静静 刘加慧
(1. 广东海洋大学水产学院 湛江 524088; 2. 湛江市国兴水产科技有限公司 湛江 524025)
凡纳滨对虾(Litopenaeus vannamei), 又称南美白对虾、白对虾, 原产于美洲太平洋沿岸水域, 主要分布于秘鲁北部至墨西哥湾沿岸, 以厄瓜多尔沿岸最为集中, 是世界上养殖产量最高的三大优良下种之一(王兴强等, 2004; 李世凯等, 2012)。近年来由于野生凡纳滨对虾资源的减少、海水水质的恶化、对虾病毒性和细菌性病害的频发, 国内外的相关研究机构和育种企业展开的凡纳滨对虾的遗传改良研究以适应凡纳滨对虾养殖业的发展。近交效应是对虾的遗传改良中应当考虑的因素。适当的近交有利于优良性状的稳定固化, 不恰当的近交会导致近交衰退影响遗传改良的进程(张沅, 2001)。
目前, 国内外学者也展开了近交效应对对虾遗传改良影响的研究。Moss等(2007)研究发现近交系数每上升10%, 凡纳滨对虾生长速度下降2.6%到3.9%呈显著相关性, 建议在凡纳滨对虾的遗传改良中所构建家系的近交系数不能超过10%。Moss等(2008)研究还发现凡纳滨对虾孵化率的近交衰退系数为-12.3%,无节幼体的成活率的近郊衰退系数为-11.0%, 养成存活率的近郊衰退系数为-3.8%。De Donato等(2005)研究发现在凡纳滨对虾选择育种的初期近交衰退可以避免甚至消失, 随着选育的进行, 体质量性状的变异系数由19.7%下降至11.6%。曹宝祥等(2015)研究发现凡纳滨对虾近交群体与选育群体相比, 近交系数每增加10%, 引起的收获时体质量的近交衰退系数-6.60%,存活率性状的实际衰退系数为11.3%。可见, 已有的报道主要集中在近交对凡纳滨对虾生长和存活性状的影响研究。而关于不同程度近交对凡纳滨对虾家系生长及抗逆性的影响的研究鲜有报道。
为此, 本课题组展开了不同近交程度对凡纳滨对虾生长性状、存活性状、收获时高氨氮抗逆性及收获时低溶氧抗逆性影响的研究, 旨在为凡纳滨对虾高生长高抗逆新品系的进一步良种选育工作提供依据和参考。
1 材料与方法
1.1 试验材料
2015年5月, 课题组利用ASRmel4.0软件, 对家系选择 4代的留种家系的凡纳滨对虾亲虾(部分留种家系间具有亲缘关系)设计交配组合。设计中度近交组合 GM(近交系数大于 0.25)4个家系, 分别为 G1、G2、G3、G4; 轻度近交组合 GL(近交系数为 0.25—0.0625)4个家系, 分别为Z1、Z2、Z3、Z4; 非近交组GN为非近交家系4个(近交系数为0), 分别为F1、F2、F3、F4。所构建16家系间没有亲缘关系。上述家系为采用人工授精方式, 10d内建成。
1.2 试验方法
试验构建家系苗种的培育过程中采用标准化的操作流程, 以最大限度地降低育苗操作对家系培育的影响。待试验家系仔虾养殖至PL-15期, 随机挑选900尾, 均分为3组, 转移至3个20网目2.5m2的网箱, 于1个水泥池中进行同池共同环境养殖, 保证各养殖网框饵料投喂量保持一致, 每天检查水体氨氮、溶氧等理化指标, 清扫网框中粪便残饵。试验家系共同环境养殖 120d时, 分别统计每池中各家系的成活率, 中随机抽取各家系对虾30尾, 测量体质量(BW)、体长(BL)和腹节长(ABS), 记录每尾测量对虾的性别。共同环境养殖90d时, 从各家系挑取体长相近的对虾120尾(体长为7.0—7.5cm), 设计3个平行试验,1个对照试验, 各平时试验用虾30尾进行96h高氨氮胁迫试验(预实验显示 96h高氨氮胁迫半致死浓度为110mg/L), 用氯化铵(分析纯)配置半致死浓度试验海水, 每隔24h完全更换含氯化氨氮的海水, 试验过程中试验海水溶解氧为7.0—8.0mg/L, pH 7.9—8.1, 水温 25—27℃; 各家系挑取体长相近的对虾 120尾(体长为7.00—7.5cm), 设计3个平行试验, 1个对照试验,各平行试验用虾30尾, 进行96h低溶氧胁迫试验(预实验显示 96h低溶氧胁迫半致死浓度为 0.82mg/L),试验时将对虾放入水中耗养至半致死浓度, 再通过控制供氧量使氧浓度维持在预定值水平, 试验海水pH为 7.9—8.1, 水温 25—27℃。统计高氨氮耐受存活率(SRA)和低溶氧耐受存活率(SRH), 具体试验方法如袁瑞鹏等(2015)试验研究所述。
1.3 数据处理
应用 SPSS17.0软件对不同组合家系的生长性状、存活性状、高氨氮耐受存活率、低溶氧耐受存活率的相关参数进行统计描述、单因素方差分析(ANOVA)和Duncan多重比较分析, 以P<0.05作为差异显著性水平。
成活率、家系体重变异系数的计算公式如下:
存活率(Survival rate, SR%)=试验时对虾个体数/试验开始时对虾个体数×100%家系体质量变异系数(The coefficient of variation in body weight of a family, CBW%)=家系体质量均值/家系体质量标准差
实际衰退百分比=(非近交组相关测试性状的最小二乘平均值-近交家系相关测试性状的最小二乘平均值)/非近交组相关测试性状的最小二乘平均值
应用ASReml4.0软件中的Predict指令估测试验不同家系凡纳滨对虾生长性状、存活性状、高氨氮耐受性及低溶氧耐受性的最小二乘均值。
生长性状的最小二乘均值估计模型:
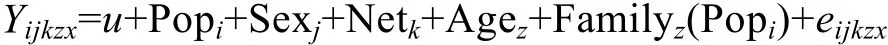
式中, Yijkzx为x尾对虾某生长性状值, u为某生长性状总体均值, Popi为第i近交处理组固定效益, Sexj表示第 j个性别固定效应, Netk为第 k个网箱固定效应,Agez为第z个家系的生长日龄作协变量, Familyz(Popi)表示第z个近交处理组内第i个家系的随机效应, eijkzx表示第x尾虾的随机残差。
存活性状、高氨氮及低溶氧抗逆性的估计模型:

式中, Yikzx为第x尾虾的存活状态(0为死亡, 1为存活),λikzx为潜在变量, 如果 λikzx>0 则 Yikzx=1, 如果 λikzxk≤0则Yikzx=0, u为某生长性状总体均值, Popi为第i近交处理组固定效益, Agez为第z个家系的生长日龄作协变量, Familyz(Popi)第i个近交处理组内第z个家系的随机效应, eikzx表示第x尾虾的随机残差。
各性状的近交衰退系数(Inbreeding depressing coefficient, IDC)和实际衰退百分比计算公式如下:

式中,为近交家系的相关测试性状的最小二乘均值,为比照组相关测试性状的最小二乘平均值,为近交家系的近交系数, 为对照组的近交系数。
2 结果
2.1 不同近交程度家系间生长性状的比较
不同近交程度凡纳滨对虾家系的近交系数、生长相关性状、体质量变异(CBW)如表 1所示。GN组所对应的家系近交系数为0, GL组所对应的家系近交系数为0.0938—0.1094, GM组所对应家系的近交系数为0.2813—0.4375。各家系BW间差异显著(P<0.05), Z2家系 BW 值最高为 8.59±3.85g, 显著大于 Z1、Z3、G1、G2、G4 家系(P<0.05); Z2、F1、F2、F3、F4、Z4、G3家系的BW值较高且两两间BW值差异不显著(P<0.05)。各家系 BL 间差异显著(P<0.05), 其中G2家系的BL最小为5.18±0.57cm, 显著小于F1、F2、F3、F4、Z1、Z2、Z4、G1、G3、G4 家系的 BL 值(P<0.05)。各家系ABS间差异显著(P<0.05), 其中 Z2家系显著大于 Z1、Z3、G2、G4家系(P<0.05); G2家系 ABS值最小, 小于除Z1、G1、G4外的其它家系。各家系中G2家系的CBW最大为49.91%, Z2家系CBW最小为20.01%, GN组家系CBW为25.11%—49.91%, GL组家系CBW为20.01%—46.24%, GM组家系CBW为25.89%—34.07%。

表1 不同近交程度凡纳滨对虾家系的生长相关性状Tab.1 The growth related traits of L. vannamei families in different degrees of inbreeding
2.2 不同近交程度家系间存活、高氨氮及低溶氧耐受性比较
不同近交程度凡纳滨对虾家系共同环境养殖90d存活(SR)、高氨氮耐受存活率(SRA)及低溶氧耐受存活率(SRH)如表 2所示。GN组家系 SR为 46.97%—63.78%, GL组 SR为 36.73%—61.73%, GM组 SR为58.74%—84.81%。不同家系SR间差异显著(P<0.05),G2家系SR最高为84.81%±10.37%, 显著高于其它家系 SR(P<0.05), Z4家系 SR最低为 36.73%±14.41%,显著低于其它家系(P<0.05)。各家系SRA间差异显著(P<0.05), 其中 G4、F3值最高, 分别为 76.92%±14.73%、76.71%±15.81%, 显著高于除 F1、Z2外的其它家系(P<0.05)。各家系 SRH间差异显著(P<0.05),其中 Z3家系 SRH最高, 为 52.63%±14.31%, 显著关于除F4家系外的其它家系(P<0.05), G1家系SRH最低, 为 15.43%±14.39%, 显著小于其它家系(P<0.05)。
2.3 不同近交处理组间生长、存活、高氨氮及低溶氧耐受性情况
不同近交程度处理组凡纳滨对虾的体质量(BW)、体质量变异系数(CBW)、共同环境养殖存活(SR)、高氨氮耐性(SRA)及低溶氧耐受性(SRH)情况如表 3所示。不同近交程度处理组 BW 间差异显著(P<0.05), 其中 GN组 BW最高, 为 7.66±2.86g, 显著高于 GM组, GL组的 BW 与其它两组间差异不显著(P>0.05)。不同近交程度处理组 CBW 间差异显著,GM组 CBW最小, 为29.30%±3.59%, 显著小于其它处理组(P<0.05)。不同近交程度处理组SR间差异不显著(P>0.05), 其中 GM组最高, 为 69.62%±16.91%;GL组最低, 为 49.44%±17.34%。不同近交程度处理组 SRA间差异不显著(P>0.05), 其中 GN组最高, 为67.63%±7.70%; GL组最低, 为58.26%±10.24%。不同近交程度处理组SRH间差异不显著(P>0.05), 其中GN组最高, 为 36.63%±8.72%; GM组最低, 为 31.25%±12.69%。
2.4 近交家系体质量和存活性状的近交衰退情况
如表4所示, 轻度近交家系Z2和中度近交家系G3的体质量最小二乘均值高于非近交组均值, 分别为8.61和7.82, 其余6个近交家系体质量最小二乘均值均小于对照组均值。与非近交组(GN)相比较, 轻度近交组(GL)和中度近交组(GM)的体质量实际衰退百分比为11.02%和20.36%, 近交系数每增加10%所引起的近交衰退系数分别为-10.03%和-5.83%。3个近交家系存活率最小二乘均值低于非近交组(GN)均值表现出近交衰退, 另有5个近交组家系未表现出近交衰退; 与非近交组(GN)相比较, 轻度近交组(GL)的存活率实际衰退百分比为9.35%, 近交系数每增加10%所引起的近交衰退系数为-6.41%, 而中度近交组(GM)的存活率未表现出近交衰退。

表2 不同近交程度凡纳滨对虾家系的存活率、高氨氮耐受性及低溶氧耐受性Tab.2 The survival rate, high-concentration ammonia nitrogen tolerance, and hypoxia tolerance of L. vannamei families in different degrees of inbreeding
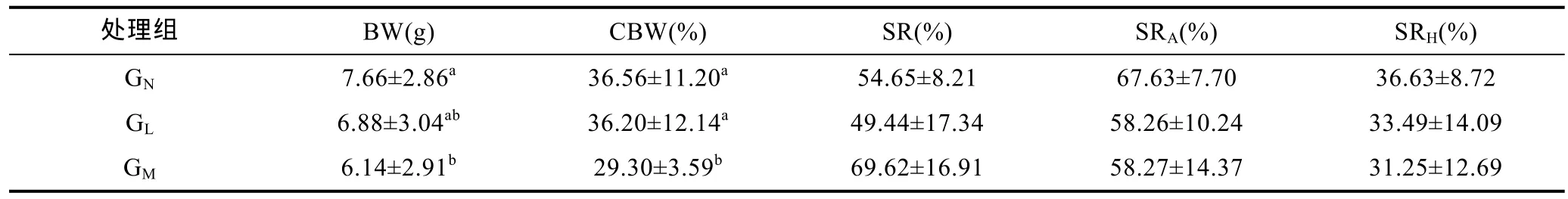
表3 不同处理组凡纳滨对虾的体质量、存活、体质量变异系数、高氨氮耐性及低溶氧耐受性Tab.3 The body weight, variation coefficient of body weight, survival rate, high-concentration ammonia nitrogen tolerance and hypoxia tolerance of L. vannamei families in different degrees of inbreeding

表4 凡纳滨对虾近交家系体质量和存活性状近交衰退情况Tab.4 Inbreeding depression of body weight and survival rate of L. vannamei families in different degrees of inbreeding
2.5 近交家系高氨氮耐受性和低溶氧耐受性的近交衰退情况
如表5所示, 轻度近交家系Z2和中度近交家系G4高氨氮耐受存活率最小二乘均值大于非近交组最小二乘均值, 未表现出高氨氮耐受近交衰退; 其余 6个近交家系高氨氮耐受存活率最小二乘均值小于非近交组最小二乘均值, 表现出现高氨氮耐受性近交衰退; 与非近交组(GN)相比较, 轻度近交组(GL)和中度近交组(GM)的高氨氮耐受性实际衰退百分比为13.95%和14.84%, 近交系数每增加10%所引起的近交衰退系数分别为-13.63%和-4.73%。轻度近交家系Z3和中度近交家系G2和 G4低溶氧耐受存活率最小二乘均值大于非近交组最小二乘均值, 未表现出低溶氧耐受近交衰退; 其余 5个近交家系低溶氧存活率最小二乘均值小于非近交组最小二乘均值, 表现出现高低溶氧耐受性近交衰退; 与非近交组(GN)相比较, 轻度近交组(GL)和中度近交组(GM)的低溶氧耐受性实际衰退百分比为 9.81%和 16.59%, 近交系数每增加 10%所引起的近交衰退系数分别为-11.84%和-4.77%。

表5 凡纳滨对虾近交家系高氨氮耐受性和低溶氧耐受性近交衰退情况Tab.5 The inbreeding depression coefficient of growth related traits, survival, high ammonia nitrogen tolerance and hypoxia tolerance of L. vannamei families in different degrees of inbreeding
3 讨论
3.1 不同程度近交对凡纳滨对虾生长性状的影响
在遗传选育的闭锁群体中, 在刚开始进行闭锁选育时, 选育工作者会有意识地避免近交, 但由于逐年淘汰, 和选育规模的受限, 选育多年后选育群体难以避免会产生不同程度的近交(郭坤德等, 1984)。本研究对闭锁繁育条件下, 产生的不同程度近交的凡纳滨对虾群体和家系的生长和存活情况进行了分析,中度近交组的体质量为 6.14±2.91g, 显著小于非近交组体质量7.66±2.86g (P<0.05), 轻度近交组近交系数每增加10%所引起体质量的近交衰退系数为-10.03%,中度近交组为-5.83%。曹宝祥等(2015)发现全同胞近交群体与选育群体体质量产生 16.68%的衰退, 近交系数每增加 10%所引起的近交衰退系数为-6.60%;Moss等(2008)在桃拉病毒和白斑病毒感染的环境中凡纳滨对虾, 近交系数每增加 10%所引起的生长的衰退为2.6%—3.9%; Keys等(2004)研究表明近交系数为0.28—0.31的日本囊对虾(Marsupenaeus japonicus)的近交衰退系数为-3.34%; 相关学者(杨翠华等,2007; 张洪玉等, 2009; 罗坤等, 2014)也发现中国明对虾(Fenneropenaeus chinensis)全同胞近交一代或者两代后, 生长性状表现出近交衰退。可见, 由于近交导致对虾生长性能衰退的现象普遍存在, 在育种工作中应避免无规划近交个体的产生。本研究中中度近交组体质量实际衰退百分比为 20.36%, 高于轻度近交组体质量实际衰退百分比 11.02%, 表明近交系数越高体质量衰退也严重。傅强等(2015)在虾夷扇贝中也发现了类似的现象, 表明在水产生产中应避免高度近交个体的产生。本研究中非近交组体质量的变异系数大于中度近交组体重变异系数, 并显著大于高度近交组体质量变异系数, 笔者认为可能是近交导致控制体质量基因多样性的降低, 提高了体质量性状的同质性。
本研究中就单个近交家系而言, 近交不一定会产生生长性能的衰退, 中度近交家系Z2、高度近交家系 G3体质量均值, 高于各对照组体质量值, 表现出一定的近交优势; 其余6个近交家系均表现出体质量近交衰退现象。可能原因是, 构建试验近交家系所用亲虾为选育留种亲虾, 具有良好的生产性能, 近交使控制快速生长的有益基因快速聚合、纯化, 使得近交家系表型出良好的生长性能。刘小军等(1993)发现一定程度的近交对猪的初生体重及生长发育有良好的作用, 建议近交程度不宜低于 12.5%, 认为合理有效地使用近交将进一步提高专门化品系的质量。郭坤德等(1984)发现闭锁繁育的星布罗肉用种鸡家系近交效果, 视其共同祖先和亲本的性能而定, 早期生长速度快的家系其后代表现较好, 半同胞交配时退化幅度较小, 有的性状, 如受精卵、饲料转换率有所提高;共同祖先和亲本早期生长速度慢的家系, 其后代表现较差。目前, 近交效应对水产动物育种的研究, 多为全同胞交配或半同胞交配所产近交群体与未近交群体的简单比较(张国范等, 2003; 刘振等, 2013; 梁健等, 2013; 喻驰方等, 2013), 这样不利于合理有效近交的发现。笔者认为, 在水产动物育种中家系选择和谱系选择相结合, 记录多代留种交配亲本的生产性能, 合理设计亲缘选配方案, 对近交产生后代实施严格淘汰, 是通过近交加快育种进程的有效途径。
3.2 不同程度近交对凡纳滨对虾存活、高氨氮及低溶氧耐受性的影响
不同的学者对近交对环境压力的影响有不同的观点。Dahlgaard等(2000)认为近交与否跟环境的适应力无必然联系。Bijlsma等(1999)与 Miller(1994)认为环境压力会增大近交效应的影响。本试验研究了近交对环境因子耐受的影响, 轻度近交组近交系数每增加 10%所引起高氨氮耐受性的近交衰退系数为-13.63%, 中度近交组近交系数每增加 10%所引起高氨氮耐受性的近交衰退系数为-4.73%; 轻度近交组近交系数每增加 10%所引起低溶氧耐受性的近交衰退系数为-11.84%, 中度近交组近交系数每增加 10%所引起高氨氮耐受性的近交衰退系数为-4.77%; 但上述性状近交组与非近交组间差异不显著(P>0.05)。张洪玉等(2009)研究发现发现, 近交中国明对虾群体与非近交群体的盐度耐受性和高温耐受性间差异不显著(P>0.05)。相关学者(Shikano et al, 2001; Nakadate et al, 2003)发现近交对孔雀鱼(Poecilia reticulata)的盐度耐受性影响显著(P<0.05), 而对其高温耐受性影响不显著(P>0.05)。可见, 近交效应对不同水产动物环境适应力影响差异较大, 即使同一近交水产动物对不同环境因子改变适应力也表现不同。本研究中25%的近交家系家系表现共同养殖存活有近交衰退,75%的家系并未出现近交衰退。Moss等(2007)也发现在近交的凡纳滨对虾存活性状未出现明显的近交衰退现象, 但在病毒感染后近交的凡纳滨对虾存活性状出现明显的近交衰退现象。De Donato等(2005)其他学者的试验发现近交会导致凡纳滨对虾存活性状出现衰退现象。笔者认为, 近交对凡纳滨对虾存活性状衰退或进展的影响, 受养殖环境和近交个体亲本遗传背景的影响, 应进一步分析养殖环境与近交对对虾存活性能的影响。
王兴强, 马 甡, 董双林, 2004. 凡纳滨对虾生物学及养殖生态学研究进展. 海洋湖沼通报, (4): 94—100
刘 振, 闫喜武, 霍忠明等, 2013. 蛤仔白斑马品系的家系选择及近交效应研究. 大连海洋大学学报, 28(4): 355—360
刘小军, 周 庆, 1993. 继代选育中猪的近交效应分析. 山西农业大学学报, 13(4): 351—353
李世凯, 江 敏, 戴习林等, 2012. 凡纳滨对虾池塘水质及对虾肌肉品质的对比分析. 上海海洋大学学报, 21(6):955—964
杨翠华, 孔 杰, 王清印等, 2007. 控制环境养殖下近交对中国对虾早期体重和抗WSSV性状的影响. 水产学报, 31(2):226—234
张 沅, 2001. 家畜育种学. 北京: 中国农业出版社, 214—219
张国范, 刘述锡, 刘 晓等, 2003. 海湾扇贝自交家系的建立和自交效应. 中国水产科学, 10(6): 441—445
张洪玉, 罗 坤, 孔 杰等, 2009. 近交对中国明对虾生长、存活及抗逆性的影响. 中国水产科学, 16(5): 744—750
罗 坤, 孔 杰, 栾 生等, 2014. 中国对虾选育群体与近交群体不同生长时期的生长性状和存活率的比较. 中国海洋大学学报, 44(7): 51—57
袁瑞鹏, 刘建勇, 张嘉晨等, 2015. 凡纳滨对虾群体杂交与自交 F1低溶氧与高氨氮耐受性比较. 中国水产科学, 22(3):410—417
郭坤德, 朱景瑞, 1984. 闭锁鸡群的近交试验. 南京农业大学学报, 7(2): 80—92
曹宝祥, 孔 杰, 罗 坤等, 2015. 凡纳滨对虾选育群体与近交群体、引进群体生长和存活性能比较. 水产学报, 39(1):42—51
梁 健, 闫喜武, 霍忠明等, 2013. 不同近交系数蛤仔橙色品系的生长和成活研究. 南方水产科学, 9(4): 87—93
喻驰方, 栾 生, 张志伟等, 2013. 中华卤虫不同近交水平家系生长性能比较. 渔业科学进展, 34(6): 75—79
傅 强, 王 师, 赵 亮等, 2015. 不同近交系数的虾夷扇贝近交衰退研究. 中国海洋大学学报(自然科学版), 45(11):43—48
Bijlsma R, Bundgaard J, Van Putten W F, 1999. Environmental dependence of inbreeding depression and purging in Drosophila melanogaster. Journal of Evolutionary Biology,12(6): 1125—1137
Dahlgaard J, Hoffmann A A, 2000. Stress resistance and environmental dependency of inbreeding depression in Drosophila melanogaster. Conservation Biology, 14(4):1187—1192
De Donato M, Manrique R, Ramirez R et al, 2005. Mass selection and inbreeding effects on a cultivated strain of Penaeus (Litopenaeus) vannamei in Venezuela. Aquaculture,247(1—4): 159—167
Keys S J, Crocos P J, Burridge C Y et al, 2004. Comparative growth and survival of inbred and outbred Penaeus(marsupenaeus) japonicus, reared under controlled environment conditions: indications of inbreeding depression. Aquaculture, 241(1—4): 151—168
Miller P S, 1994. Is inbreeding depression more severe in a stressful environment? Zoo Biology, 13(3): 195—208
Moss D R, Arce S M, Otoshi C A et al, 2007. Effects of inbreeding on survival and growth of Pacific white shrimp Penaeus (Litopenaeus) vannamei. Aquaculture, 272(S1):S30—S37
Moss D R, Arce S M, Otoshi C A et al, 2008. Inbreeding effects on hatchery and growout performance of Pacific white shrimp, Penaeus (Litopenaeus) vannamei. Journal of the World Aquaculture Society, 39(4): 467—476
Nakadate M, Shikano T, Taniguchi N, 2003. Inbreeding depression and heterosis in various quantitative traits of the guppy, Poecilia reticulata. Aquaculture, 220(1—4): 219—226
Shikano T, Chiyokubo T, Taniguchi N, 2001. Effect of inbreeding on salinity tolerance in the guppy (Poecilia reticulata).Aquaculture, 202(1—2): 45—55
猜你喜欢
园林科技(2020年2期)2020-01-18 03:28:18
当代水产(2019年7期)2019-09-03 01:02:26
当代水产(2019年1期)2019-05-16 02:42:00
福建质量管理(2019年2期)2019-01-22 03:04:56
农家之友(2018年8期)2018-08-24 08:22:32
大电机技术(2017年3期)2017-06-05 09:36:02
天津科技大学学报(2016年1期)2016-02-28 16:59:44
广东海洋大学学报(2015年4期)2016-01-13 08:39:32
中国医学科学院学报(2015年5期)2015-03-01 04:03:37
中国医药科学(2015年15期)2015-02-27 12:32:28
