
氮磷钾施用量对冬油菜几种酶活性的影响
2015-12-25 12:21韩顺昌李学才
甘肃农业科技 2015年6期
韩顺昌, 李学才
(1.甘肃农业大学农学院,甘肃 兰州 730070;2.甘肃省永昌县清河现代农业循环经济产业园区管委会,甘肃 永昌 737200)
油菜是需肥较多的作物,要实现高产、优质,必须满足油菜对养分的需要。其中以氮、磷、钾的作用尤其突出,氮能促进有机体生长发育,增加分枝[1],磷主要是对能量传递体系起介质的作用[2],钾对油菜体内多种重要的酶类起催化剂的作用[3]。因此,如何施用氮、磷、钾肥,一直是人们广泛关注的问题。有研究表明,单施氮肥可促进冬油菜的生长,而从越冬情况来看,冬前单施氮肥对冬油菜越冬不利;磷、钾配施可使冬油菜成熟期推迟,全生育期延长;氮、磷、钾肥配施对与冬油菜抗性有关的超氧化物歧化酶、脯氨酸、可溶性蛋白、可溶性糖等都有显著的影响[4-6]。氮、磷、钾配施比单施一种肥料明显改善生长状况并提高产量和抗性[7],主要是因为氮、磷、钾肥配施能够促进油菜对养分的均衡吸收。我们研究不同氮、磷、钾配比对冬油菜酶活性的影响,以期为指导油菜合理施肥提供参考。
1 材料与方法
1.1 试验材料
氮肥为尿素(N≥46.4%),刘家峡化工集团生产;磷肥为颗粒状普通过磷酸钙(P2O5≥12%),白银天丰磷复肥有限公司生产;钾肥为硫酸钾(K2O=50%),俄罗斯进口。指示油菜品种为陇油7号。
1.2 试验地概况
试验设在兰州市秦王川试验基地,位于北纬36°13′~36°53′,东经 103°27′~104°21′,海拔1 900 m。年日照时数2 660~2 768 h,夏季平均日照时数8~9 h,年总辐射量543.4~557.3 kJ/cm2,年平均气温5.9℃,年平均无霜期125~175 d,最高气温33.3℃,最低气温-23.5℃,≥0℃的年平均积温为3 324.5℃,≥10℃的年平均积温为2 798.3℃。年降水量260~290 mm,年蒸发量1 950 mm。土壤为大白土和砂壤土相间。土壤中性偏碱,其中有机质9.7 g/kg、全氮0.714 g/kg、全磷0.924 g/kg,pH 8.7。前茬作物为小麦。
1.3 试验方法
采用“3414”试验设计,以不施肥为对照(CK),共14个处理,小区面积15 m2(2.5 m×6.0 m),行距20 cm,株距10 cm,不设重复,播种方式为开沟点播。2013年8月20日播种。
酶液提取参照Zhu等的方法[8-9],并稍作改动。称取不同处理的样品叶片0.5 g,在冰浴条件下置于预冷过的研钵中,加入50 mmol/L的pH 7.8磷酸缓冲液(内含1%聚乙烯吡咯烷酮)5 mL研磨至匀浆,转入离心管,在4℃、12 000 r/min条件下离心15 min,上清液即为酶液。
超氧化物歧化酶(SOD)活性测定参照Huang等的方法[10],并稍作改动。配制反应混合液(50 mmol/L的pH 7.8磷酸缓冲液15 mL、0.1 mmol/L的EDTA 3 mL、0.75 mmol/L的氮蓝四唑3 mL、130 mmol/L的甲硫氨酸3 mL、0.02 mmol/L的核黄素3 mL及蒸馏水2.5 mL),吸取3 mL反应混合液,加入20μL酶液,在4 000 Lx条件下光照30 min,之后在560 nm处测定吸光度,其中SOD活性以抑制氮蓝四唑50%为1个酶活性单位。
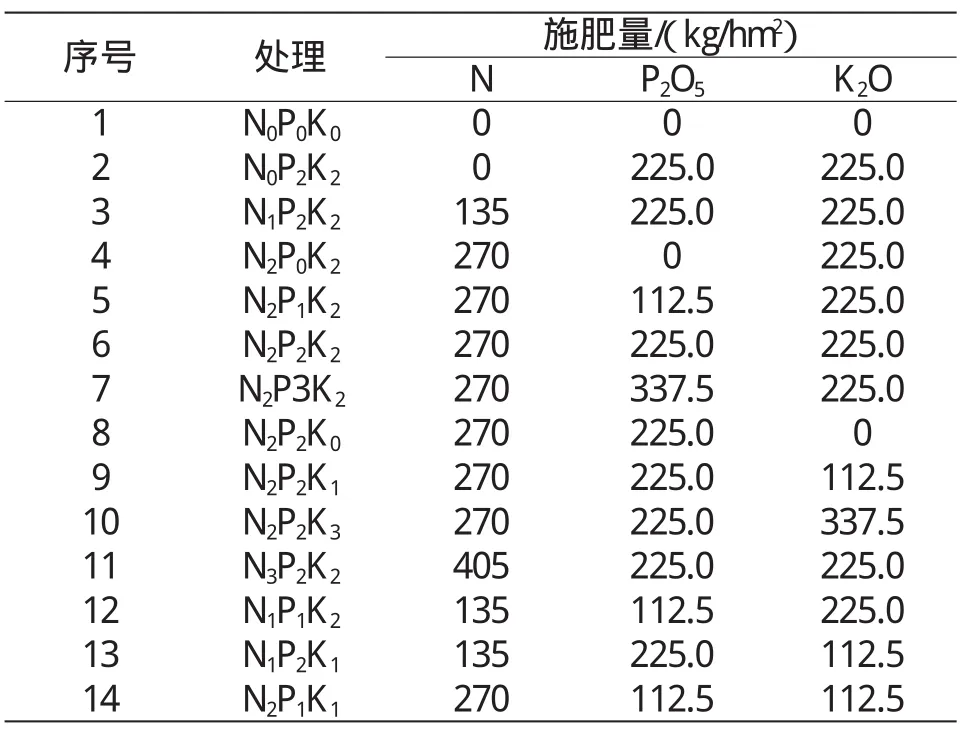
表1 不同处理的施肥量
过氧化物酶(POD)活性测定参照Shi等的方法[11],并稍作改动。配制反应混合液(0.1 mol/L的pH 6.0磷酸缓冲液50 mL、愈创木酚28μL和30%H2O219μL),吸取3 mL反应混合液加入20μL酶液,在470 nm处测定吸光度,每隔30 s读数1次,以30 s吸光度的增加值来表示酶活性的大小。
过氧化氢酶(CAT)活性测定参照Aebi的方法[12],并稍作改动。配制反应混合液(0.1 mol/L的pH 7.0磷酸缓冲液20 mL和0.1 mol/L的H2O25 mL),吸取3 mL反应混合液,加入100μL酶液,在240 nm处测定吸光度,每隔1 min读数1次,以1 min内1μmol H2O2的分解量来表示酶活性的大小。
抗坏血酸过氧化物酶(APX)活性测定参照Nakano和 Asada的方法[13]。配制反应混合液(50 mmol/L pH 7.0的PBS缓冲液 1.8 mL、15 mmol/L AsA 0.1 mL、0.3 mmol/L H2O21 mL),吸取3 mL反应液,加入0.1 mL酶液(以不加酶液为对照),在290 nm处测定吸光度,每隔1 min读数1次,以1 min OD290变化0.01为1个酶活性单位。

图1 不同施肥水平对冬油菜SOD活性的影响
2 结果与分析
2.1 氮、磷、钾不同施用量对冬油菜SOD活性的影响
氮、磷、钾不同施用量对冬油菜SOD活性的影响不同。由图1 A可以看出,处理2(N0P2K2)、处理3(N1P2K2)、处理6(N2P2K2)和处理11(N3P2K2)的SOD活性随施氮量的增加呈现上升趋势,当氮肥水平达到最高时,即在处理11(N3P2K2)下,冬油菜SOD活性比对照升高26%,因此可知,当磷肥和钾肥处于比较高的水平时,配施氮肥对冬油菜SOD活性有影响。由图1 B可以看出,处理8(N2P2K0)、处理9(N2P2K1)、处理6(N2P2K2)和处理10(N2P2K3)的SOD酶活性随施钾量的增加呈现先上升后下降而后再上升的变化趋势,在处理10(N2P2K3)下,冬油菜的SOD活性比对照增加了52.0%,因此可知,当氮肥和磷肥处于比较高的水平时,配施钾肥对冬油菜SOD活性有明显的影响。由图 1 C可以看出,处理4(N2P0K2)、处理5(N2P1K2)、处理6(N2P2K2)和处理7(N2P3K2)的 SOD酶活性随施磷量的增加呈现先下降后上升再下降的变化趋势,而且当磷肥水平达到最高时,即在处理7(N2P3K2)下,冬油菜SOD酶活性比对照(N0P0K0)下降了20%,由此可知,当氮肥和钾肥处于比较高的水平时,配施磷肥对冬油菜SOD酶的活性有抑制作用。综上所述,钾肥对冬油菜SOD活性的影响最明显,而且在N2P2K3的处理下,SOD酶活性达到最高,明显高于对照和其它处理。
2.2 氮、磷、钾不同施用量对冬油菜POD活性的影响
由图2A可以看出,处理2(N0P2K2)、处理3(N1P2K2)和处理6(N2P2K2)的冬油菜 POD活性随施氮量的增加呈现先下降后升高再下降的趋势,当氮肥的水平达到最高时,即处理11(N3P2K2)下,冬油菜POD活性反而比对照下降40.8%,因此可知,当磷肥和钾肥处于比较高的水平时,配施氮肥对冬油菜POD活性有抑制作用。由图2B可以看出,处理8(N2P2K0)、处理9(N2P2K1)、处理6(N2P2K2)和处理10(N2P2K3)的冬油菜POD活性随施钾量的增加呈现出先下降后升高的趋势,尤其是在处理10(N2P2K3)下,冬油菜的POD活性比对照增加了47.2%,因此可知,当氮肥和磷肥处于比较高的水平时,配施钾肥对冬油菜POD活性影响明显。由图 2C可以看出,处理4(N2P0K2)、处理5(N2P1K2)和处理6(N2P2K2)的冬油菜POD活性随施磷量的增加呈现先下降后升高而后下降的趋势,而且当磷肥水平达到最高时,即在处理7(N2P3K2)下,冬油菜POD活性比对照(N0P0K0)下降了20.4%,由此可知,当氮肥和钾肥处于比较高的水平时,配施磷肥对冬油菜POD的活性有抑制。综上所述,钾肥对冬油菜POD活性的影响最明显,而且在N2P2K3的处理下,冬油菜POD活性达到最高,明显高于对照和其它处理。
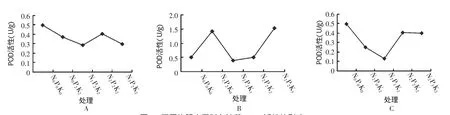
图2 不同施肥水平对冬油菜POD活性的影响

图3 不同施肥水平对冬油菜CAT活性的影响
2.3 氮、磷、钾不同施用量对冬油菜CAT活性的影响
由图 3A可以看出,处理2(N0P2K2)、3(N1P2K2)和6(N2P2K2)的CAT活性随施氮量的增加,呈先下降后升高的变化趋势,当氮肥的水平达到最高时,即在处理11(N3P2K2)下,冬油菜的CAT活性比对照升高32.3%,因此可知,当磷肥和钾肥处于比较高的水平时,配施氮肥对CAT活性的影响不明显。由图 3B可以看出,处理8(N2P2K0)、处理 9(N2P2K1)、处理6(N2P2K2)和处理10(N2P2K3)的 CAT活性随施钾量的增加呈现先升高后下降而后升高的变化趋势,尤其是在处理10(N2P2K3)下,冬油菜的CAT活性比对照增加了41.9%,因此可知,当氮肥和磷肥处于比较高的水平时,配施钾肥对冬油菜的CAT活性影响明显。图3C可以看出,处理4(N2P0K2)、处理 5(N2P1K2)和处理6(N2P2K2)的CAT活性随施磷量的增加呈现缓慢上升的趋势,当磷肥水平达到最高时,即在处理7(N2P3K2)下,冬油菜CAT活性比对照(N0P0K0)上升了30.2%,由此可知,当氮肥和钾肥处于比较高的水平时,配施磷肥对冬油菜CAT活性的影响不明显。综上所述,钾肥对冬油菜CAT活性的影响最明显,而且在N2P2K3处理下,CAT活性达到最高,明显高于对照和其它处理。

图4 不同施肥水平对冬油菜APX活性的影响
2.4 氮、磷、钾不同施用量对冬油菜APX活性的影响
由图4A可以看出,处理2(N0P2K2)、处理3(N1P2K2)和处理6(N2P2K2)的冬油菜APX活性随施氮量的增加变化不明显,当氮肥的水平达到最高时,即在处理11(N3P2K2)下,冬油菜APX活性比对照升高87.4%,因此可知,当磷肥和钾肥处于比较高的水平时,配施氮肥对APX活性的影响不明显。由图 4B可以看出,处理8(N2P2K0)、处理9(N2P2K1)、处理6(N2P2K2)和处理10(N2P2K3)的 APX活性与对照(N0P0K0)相比有明显的增加,尤其在处理10(N2P2K3)下,冬油菜的APX活性比对照增加了97.4%,因此可知,当氮肥和磷肥处于比较高的水平时,配施钾肥对冬油菜APX活性影响明显。由图 4C可以看出,处理4(N2P0K2)、处理 5(N2P1K2)和处理6(N2P2K2)的冬油菜APX活性与对照(N0P0K0)相比,变化不明显,但当磷肥水平达到最高时,即在处理7(N2P3K2)下,冬油菜APX活性比对照(N0P0K0)上升了86.7%,由此可知,当氮肥和钾肥处于比较高的水平时,配施磷肥对冬油菜APX活性的影响不显著。综上所述,钾肥对冬油菜APX活性的影响最明显,而且在N2P2K3的处理下,APX活性达到最高,明显高于对照和其它处理。
3 小结与讨论
试验结果表明,氮肥和磷肥对冬油菜体内超氧化物歧化酶活性、过氧化物酶活性、过氧化氢酶活性、抗坏血酸过氧化物酶活性影响不是很明显;而钾肥对这4种酶活性影响明显,随着施钾肥量的增加,4种酶的活性开始显著升高,并均在N2P2K3的处理下达到最高,即在N、P2O5、K2O施用量分别为270.0、225.0、337.5 kg/hm2时,APX、POD、CAT及SOD活性最高,分别比对照增加了97.4%、47.2%、41.9%和52.0%;可见,钾肥对冬油菜体内各种酶活性有显著的影响作用,这与前人钾对油菜体内多种重要的酶类起催化剂作用的研究一致[3]。
[1] 李志玉,郭庆元,廖 星.不同氮水平对双低油菜中双9号产量和品质的影响[J].中国油料作物学报,2007,29(2):78-82.
[2] 段海燕,王运华.不同甘蓝型油菜品种磷营养效率的差异研究[J]. 华中农业大学学报,2001,20(3):241-242.
[3] ZHAO F J.Sulfur and oilseed rape production in the U-nited Kingdom[J].Sulfur.in Agric.,1991,15:13-16.
[4] 魏文慧,孙万仓.氮磷钾肥对西北寒旱区冬油菜越冬率、产置及经济性状的影响[J].西北农业学报,2009,18(2):122-125;130.
[5] 郭庆元,李志玉,涂学文.我国南方红黄壤地区优质油菜营养特性与施肥研究Ⅱ.红壤区稻田双低油菜中双7号高产施肥[J].中国油料作物学报,2001,23(1):51-55.
[6] 韩燕来,介晓磊,谭金芳.超高产冬小麦氮磷钾吸收、分配与运转规律的研究[J].作物学报,1998,24(6):908-914.
[7] 刘志荣,杨滢智.氮、磷、钾、硼肥不同配方对油菜产量的影响[J].耕作与栽培,2006(4):36-37.
[8] ZHU Z B, LIANG Z S,HAN R L.Saikosaponin accumulation and antioxidative protection in drought-stressed Bupleurum chinense DC.Plants[J].Environ.Exp.Bot,2009,66:326-333.
[9] AN Y Y, LIANG Z S.Drought tolerance of Periploca sepium during seed germination:antioxidant defense and compatible solutes accumulation[J].Acta Physiol Plant,2013,35(3):959-967.
[10] HUANG X S,LIU JH,CHEN X J.Overexpression of PtrABF gene,a bZIP transcription factor isolated from Poncirus trifoliata,enhances dehydration and drought tolerance in tobacco via scavenging ROSand modulating expression of stress-responsive genes[J].BMC Plant Biol,2010,10:230.
[11] SHI S,FU X Z,PENG T,et al.Spermine pretreatment confers dehydration tolerance of citrus in vitro plants via modulation of antioxidative capacity and stomatal response[J].Tree Physiol,2010,30:914-922.
[12] AEBIH.Catalase in vitro[J].Methods Enzymol,1984,105:121-126.
[13] NAKANO Y,ASADA K.Hydrogen peroxide is scavenged by ascorabate specific peroxidase in spinach chloroplasts[J].Plant Cell Physiol,1981,22:867-880.
猜你喜欢
炼油与化工(2022年6期)2023-01-10
今日农业(2022年1期)2022-11-16
电脑迷(2022年10期)2022-11-08
发明与创新(2022年27期)2022-09-16
中国土壤与肥料(2021年5期)2021-12-02
西藏农业科技(2018年4期)2018-04-25
浙江农业科学(2016年11期)2016-05-04
大豆科技(2016年6期)2016-03-11
医学研究杂志(2015年12期)2015-06-10
新疆农垦科技(2014年7期)2014-02-28
