
干旱胁迫对白木通光合生理特性的影响
2015-12-21 08:16吴玲利雷小林柯镔峰李建安
中南林业科技大学学报 2015年11期
吴玲利,雷小林 ,龚 春 ,柯镔峰 ,熊 利,李建安
(1.中南林业科技大学 经济林培育与保护省部共建教育部重点实验室,湖南 长沙 410004;2.江西省林业科学院,江西 南昌330013)
干旱胁迫对白木通光合生理特性的影响
吴玲利1,雷小林2,龚 春2,柯镔峰1,熊 利1,李建安1
(1.中南林业科技大学 经济林培育与保护省部共建教育部重点实验室,湖南 长沙 410004;2.江西省林业科学院,江西 南昌330013)
采用盆栽试验, 研究干旱胁迫对白木通光合生理指标及叶绿素荧光参数的影响, 揭示干旱胁迫对白木通光合作用及生理指标之间的关系, 为白木通水肥管理及栽培提供参考依据。结果表明:轻度干旱(LS)提高了白木通幼苗的水分利用效率(P<0.05), 对其它光合生理指标无明显影响(P>0.05); 中度干旱(MS)使白木通叶片叶绿素含量、净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、电子传递速率(ETR)、最大光化学效率(Fv/Fm)、PSII的潜在活性(Fv/Fo)及光化学猝灭系数(qP)显著下降(P<0.05), 而对RuBP羧化酶活性、气孔导度(Gs)、实际光化学量子效率(ΦPSII) 及暗下最大荧光(Fm)影响不显著(P>0.05); 重度干旱(SS)导致白木通叶绿素含量、RuBP羧化酶活性、净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、电子传递速率(ETR)、最大光化学效率(Fv/Fm)、实际光化学量子效率(ΦPSII)、PSII的潜在活性(Fv/Fo)、暗下最大荧光(Fm)及光化学猝灭系数(qP)显著下降(P<0.05), 胞间CO2浓度(Ci)、水分利用效率(WUE)、初始荧光(Fo)、非光化学猝灭系数(CNPQ)显著升高(P<0.05); 在中度干旱胁迫下白木通幼苗光合作用的降低主要是由气孔关闭引起的,而重度干旱胁迫下光合作用的下降主要是由光合机构活性降低的非气孔因素引起的。
白木通;干旱胁迫;光合特性;RuBP羧化酶;叶绿素荧光参数
在陆地生态系统中,水分是限制植物分布和生长的最主要因素[1]。近年来,随着全球气候与环境的变化,南方大部分地区在盛夏会出现高温干旱天气,使得土壤有效水分含量减少,对植物的生长发育产生了严重影响。研究表明,干旱胁迫对作物产量的影响超过了其他因素造成减产的总和[2]。研究表明,干旱胁迫降低了香樟幼苗的净光合速率,妨碍了有机物的积累,植物在干旱胁迫下光合效率的降低受到气孔因素和非气孔因素共同作用,首要的表现为气孔关闭的影响[3-4]。曹慧等人[5]在苹果中研究表明,轻度干旱胁迫下,植物的自我调节使得光合速率保持稳定,但在中度和重度干旱胁迫下,植物的光合速率会明显降低。
白木通Akebia trifoliata (Thunb.)为木通科木通属藤本植物,为三叶木通的南部亚种。味苦,性微寒,具有清心泻火、疏肝益肾、通经散瘀、除烦利尿之功效,主要分布于江苏、浙江、江西、广西、湖南、湖北、山西、四川等地[6-9]。野生白木通果肉含粗脂肪4.03%、粗蛋白2.45%、粗纤维3.86%、灰分6.38%、总糖50.32% ,并含有丰富的VC和 Na、Mg、Fe、K、Cu、Zn、Co 等矿质元素,同时至少含有17种氨基酸,其中7种为必需氨基酸[10]。光合作用是植物最基本的生命活动,它反映了植物的遗传特性和对环境中水肥气热的适应能力,是对水分胁迫最敏感的生理过程之一,可以作为判断植物生长和抗逆性的重要指标[11]。叶绿素荧光参数对研究植物叶片光合作用过程中的光能吸收、传递、转换、耗散、分配等具有重要意义[12]。目前,关于干旱胁迫对植物光合生理的影响在苹果[13],葡萄[14],沙枣[15]等树种中进行了大量的研究。而干旱胁迫对木通光合生理方面的研究还未见报道。为此,本文使用盆栽试验,研究不同土壤相对含水量对白木通幼苗叶片叶绿素含量、光合特性、叶绿素荧光等参数的影响,通过量化木通光合生理指标以期为抗旱栽培提供理论依据。
1 材料与方法
1.1 供试材料
供试材料白木通Akebia trifoliate var. australis由江西省林业科学研究院提供,为2年生扦插基质苗,试验在中南林业科技大学校内进行。
1.2 试验设计
试验于2014年6月~9月在中南林业科技大学校内进行,选取长势一致的木通扦插苗,移栽到15 cm×15 cm×12 cm装有基质的塑料盆中,田间持水量为26.3%,有机质含量11.1 g·kg-1,全氮0.5 g·kg-1,全磷 3.7 g·kg-1,速效钾 100.2 mg·kg-1。试验设4个处理,分别为正常供水CK、轻度干旱LS、中度干旱MS和重度干旱SS,其土壤含水量分别为土壤田间最大持水量的90%~85%、75%~70%、55%~50%、35%~30%,每个处理9盆,总共36盆[16],水分控制通过称重法,试验在第一天下午充分浇透土壤,晚上过夜使多余的水分充分流干,第二天早上的盆重为田间最大持水量时的重量,用此时的盆重减去盆子土壤干重为土壤的含水量(W100),4个处理分别为W100×(90%~85%、75%~70%、55%~50%、35%~30%),为防止水分大量蒸发,盆子表面铺一层塑料薄膜,每天早上通过称重法进行补水[17]。处理一个月后测定叶绿素含量、光合参数和叶绿素荧光参数等各项指标。
1.3 光合生理指标的测定
1.3.1 叶绿素含量的测定
叶片叶绿素含量参照张宪政丙酮乙醇混合液法提取[18],用打孔器切取0.1 dm2鲜叶剪成细丝,浸泡在10 mL丙酮乙醇 (V∶V=1∶1) 溶液中暗处提取24 h,按照Arnon方法用分辨率较高的分光光度计测定663 nm和645 nm处读光密度(OD值)[19],计算叶绿素a、叶绿素b及叶绿素总量,每个处理重复测定3次。
1.3.2 光合酶活性的测定
光合作用测定完成后第2天上午9:00~11:00采集样品,各处理选取生长正常且长势长相基本相同的叶片,称取0.5 g鲜叶用锡箔纸包好做标记后立即置入液氮中速冻 30 min,放入-80℃冰箱中保存,每个处理采集6份样品备用,在取样结束一周后,统一测定其酶活性,重复3次。RuBP羧化酶的提取、纯化参照马冬云等的方法[20],将叶片冰浴研磨,匀浆于4℃下以 15 000×g 离心20 min,取上清液置0℃保存待测。RuBP羧化酶活性的测定按Racker分光光度法略加修改[21]。
1.3.3 光合参数的测定
光合指标测定于晴朗上午8:30~11:00自顶端向下选取成熟的功能叶片,用LI-6400XT便携式光合仪( LI-COR,USA)测定叶片的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)及蒸腾速率(Tr)等参数。水分利用效率(WUE)=净光合速率(Pn)/蒸腾速率(Tr);测定时用红蓝光源叶室,光照强度设为 1 200 μmol·m-2s-1,气温为30~35 ℃,CO2气体由小钢瓶提供,控制浓度为 400 μmol·m-2s-1,每个测定重复 3 次。
1.4 叶绿素荧光的测定
叶绿素荧光参数的测定用LI-6400xt光合仪测定,参考LI-6400xt操作手册,经过充分暗适应的叶片在凌晨破晓前照射检测光测定Fo,然后施加饱和脉冲为 7 200 μmol·m-2s-1的光强下 0.8 s,测得暗适应下最大荧光Fm,荧光参数的计算参照Rohácˇek的方法[22]。天亮后用叶室内活化光活化30 min以上,直接测定PSII有效光化学量子产量Fv′/Fm′、实际光化学量子效率ΦPSII、光化学猝灭系数qP及非光化学猝灭系数NPQ等参数。
1.5 数据统计与分析
用Excel 2007对数据进行处理作图,用SPSS 17.0软件进行相关性分析及其差异显著性。
2 结果与分析
2.1 干旱胁迫对白木通叶绿素含量的影响
由表1可知,随着干旱胁迫的加重,白木通叶片的叶绿素a、叶绿素b及总叶绿素含量均呈降低趋势。与对照(CK)相比,轻度干旱(LS)对叶绿素a、叶绿素b及总叶绿素含量的影响不显著(P>0.05);中度干旱使叶绿素a、叶绿素b及总叶绿素含量分别降低了17.24%、21.33%、18.35%,差异显著(P<0.05);重度干旱使叶绿素a、叶绿素b及总叶绿素含量分别降低了40.39%、38.67%、39.93%,差异及显著(P<0.05)。
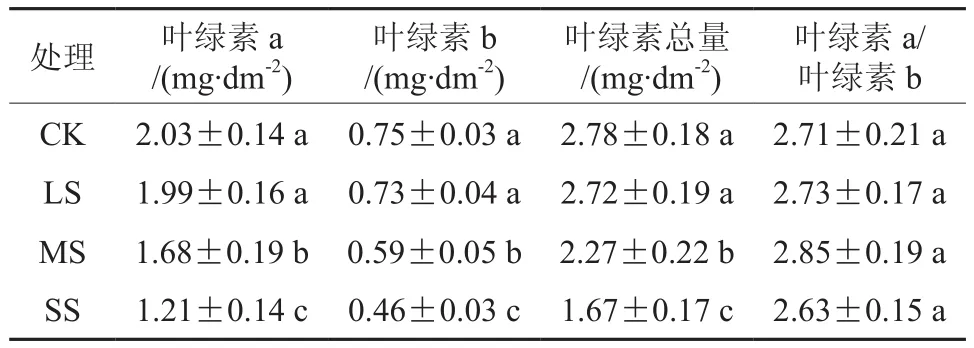
表1 干旱胁迫对白木通叶绿素含量的影响†Table 1 Effects of drought stress on chlorophyll content of A.trifoliate
2.2 干旱胁迫对白木通光合酶活性的影响
RuBP羧化酶活性的高低直接影响植物的净光合速率。由图1可知,重度干旱(SS)使白木通RuBP羧化酶活性降低了39.39%,差异显著(P<0.05);在轻度干旱(LS)与中度干旱(MS)下,RuBP羧化酶活性与对照差异不显著(P>0.05)。说明轻度干旱对白木通光合酶活性的影响较小,重度干旱严重影响了白木通的光合酶活性,从而抑制植物的正常生长。
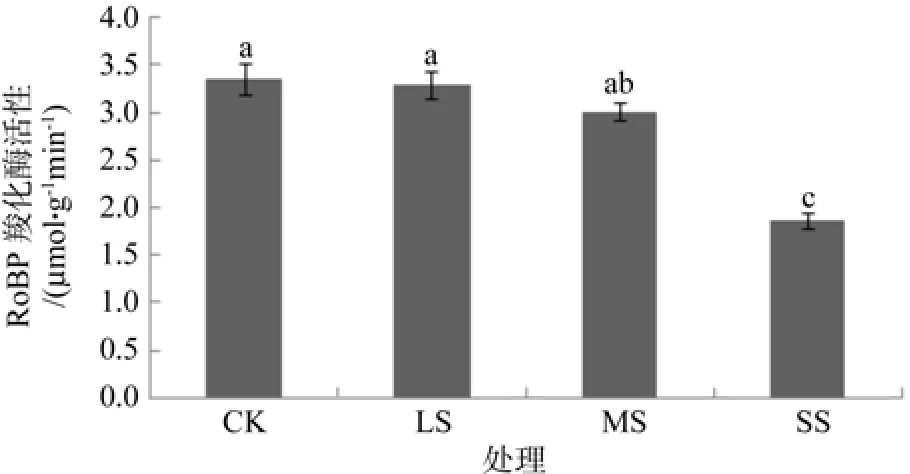
图1 干旱胁迫对白木通RuBP羧化酶活性的影响Fig.1 Effects of drought stress on RuBP carboxylase activity of A.trifoliate
2.3 干旱胁迫对白木通光合特征参数的影响
由表2所示,与对照(CK)相比,随着干旱胁迫的加重,白木通的净光合速率(Pn)、蒸腾速率(Tr)及气孔导度(Gs)依次呈降低趋势,水分利用效率(WUE)逐渐提高,胞间CO2浓度先降低后升高。轻度干旱(LS)对白木通的光合作用影响不明显;中度干旱(MS)使白木通的Pn、Tr、Gs分别降低了17.96%、25.97%、14.29%,水分利用效率提高了10.93%;重度干旱(SS)使白木通的Pn、Tr、Gs分别降低了48.73%、55.84%、35.71%,差异显著(P<0.05),重度干旱(SS)使白木通的水分利用效率提高了16.27%,说明干旱胁迫有利于白木通的水分利用效率。

表2 干旱胁迫对白木通幼苗叶片光合参数的影响Table 2 Effects of drought stress on the photosynthetic parameters of A.trifoliate
2.4 干旱胁迫对白木通叶绿素荧光参数的影响
2.4.1 干旱胁迫对白木通叶片Fo、Fm、Fv/Fm和Fv/Fo的影响
Fo表示初始荧光、暗荧光,Fm表示暗下最大荧光,Fv/Fo代表了PSII的潜在光化学效率,Fv/Fm表示PSII的最大光化学效率,Fv/Fm常用来判断植物是否受到胁迫[23]。由图2可知,随着干旱胁迫的加重,白木通的初始荧光Fo逐渐升高,Fm、Fv/Fm及Fv/Fo逐渐降低,轻度干旱(LS)对白木通Fo、Fm、Fv/Fm及Fv/Fo的影响不显著;中度干旱(MS)使Fo升高了8.57%,Fm、Fv/Fm及Fv/Fo分别降低了4.09%、4.93%、13.95%;重度干旱(SS)使Fo升高了19.43%,Fm、Fv/Fm及Fv/Fo分别降低了13.81%、8.64%、34.88%,差异显著(P<0.05)。

图2 干旱胁迫对白木通叶片Fo、Fm、Fv/Fm和Fv/Fo的影响Fig.2 Effects of drought stress on Fo, Fm, Fv/Fm and Fv/Fo of A.trifoliate
2.4.2 干旱胁迫对白木通ETR、ΦPSII、qP和NPQ的影响
ΦPSII表示PSII的实际光化学效率,它的大小反映了吸收的光子供给PSII反应中心的效率[24]。qP为光化学猝灭系数,它反映的是PSII天线色素吸收的光能用于光化学电子传递的份额。NPQ反映的是天线色素吸收的光能不能用于光合电子传递而以热等的形式耗散掉的光能部分[25]。由图3可知,白木通的ETR、ΦPSII、qP随着干旱胁迫的加重逐渐降低,轻度干旱对白木通的ETR、ΦPSII及qP的影响不显著;中度干旱使白木通的ETR及qP的值分别降低了21.6%、14.29%,差异显著(P<0.05);重度干旱使白木通的ETR、ΦPSII及qP分别降低了34.52%、41.94%、28.57%,差异显著(P<0.05)。
3 结论与讨论
水分胁迫对白木通的光合生理参数均有影响,但不同处理差异显著,通过研究发现,轻度干旱(LS)对白木通幼苗的生长无明显影响,中度干旱(MS)及重度干旱(SS)均降低了白木通叶片的叶绿素含量,由此可知,水分胁迫不利于白木通幼苗光合色素的合成,其原因可能是在水分胁迫下植物体内的叶绿素酶活性及活性氧使光合色素分解所致,这与宋丽萍等在刺五加中的研究结果相一致[26]。RuBP羧化酶是植物进行光合作用过程中碳固定的关键酶,在水分胁迫下,RuBP羧化酶的活性也降低,但在重度干旱(SS)胁迫下RuBP羧化酶活性降低的程度要大于中度干旱(MS)胁迫,同时,净光合速率,气孔导度及电子传递速率均有所降低,这说明白木通幼苗光合速率的降低与叶绿素含量、光合酶活性的降低及气孔因素均有关系。土壤水分胁迫对植物生长和代谢的影响是多方面的,其中对光合作用的影响尤为突出[27]。前人研究表明,在外界环境胁迫下,光合作用的限制因子可分为气孔因素和非气孔因素,如果气孔导度与胞间CO2浓度同时下降时,Pn下降主要是由气孔限制引起的,如果 Pn的降低伴随着胞间CO2浓度升高,光合作用的主要限制因素则是非气孔因素[28]。本研究发现,在中度干旱(MS)条件下,净光合速率降低的同时,气孔导度及胞间CO2浓度均降低,说明在中度干旱(MS)胁迫下,白木通幼苗净光合速率的降低是由于气孔关闭引起的,这也是植物在长期进化过程中的形成的一种保护机制。在重度干旱(SS)胁迫下发现,净光合速率降低,气孔导度也伴随降低,但是胞间CO2浓度升高,说明在重度干旱(SS)胁迫下已经破坏了白木通幼苗叶片的光合酶活性及光合色素含量而引起的。
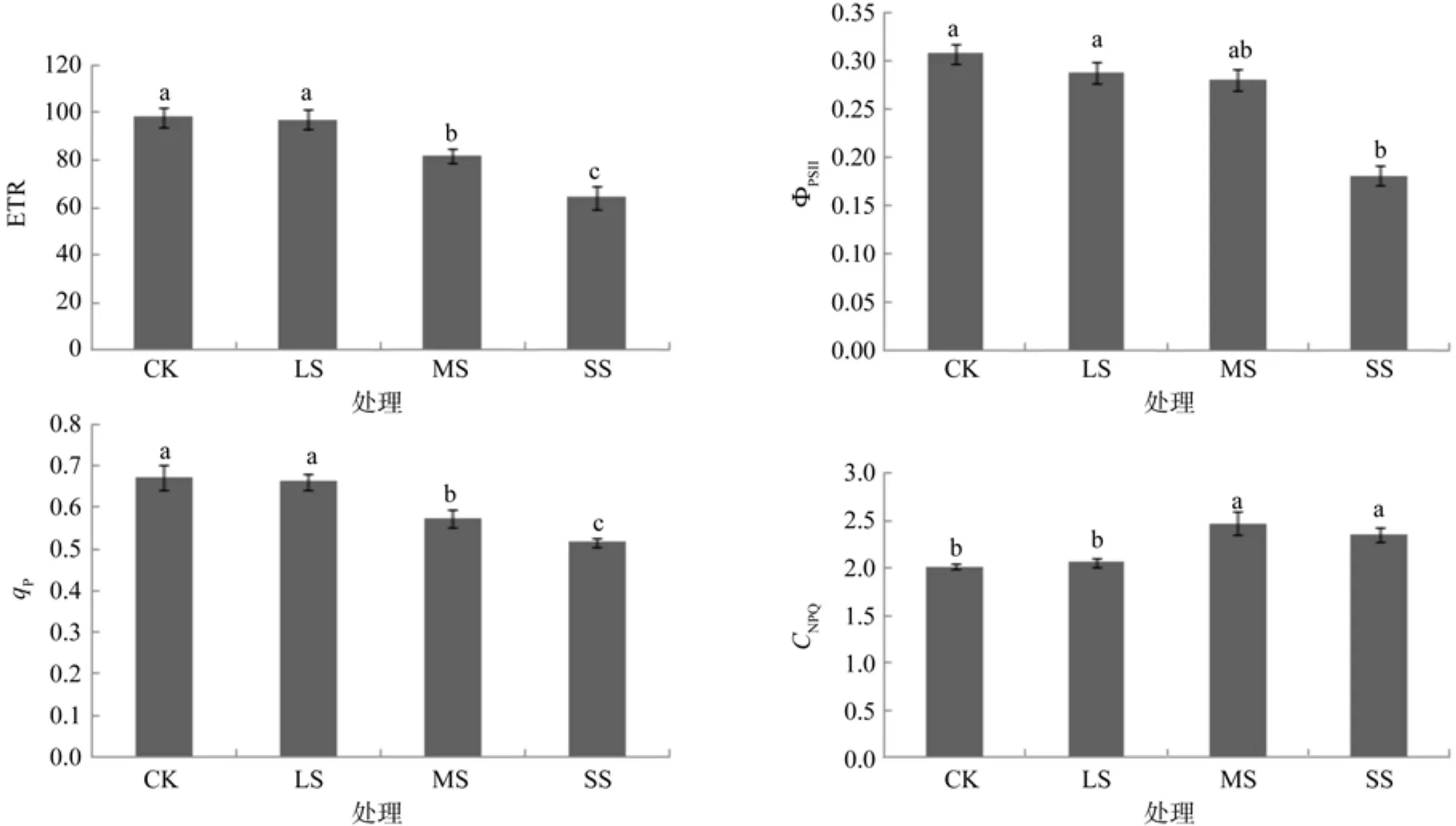
图3 不同供镁水平对油桐幼苗叶片RETR、ΦPSII、qP和CNPQ的影响Fig.3 Effects of drought stress on RETR, ΦPSII, qP and CNPQ of A. trifoliate
一般情况下,叶绿素吸收的光能主要通过光合电子传递、叶绿素荧光和热耗散3种途径消化掉,这3种途径间存在着此消彼长的关系,光合作用和热耗散的变化会引起荧光发射的相应变化,因此荧光变化可以反映光合作用和热耗散的情况[29]。在重度干旱胁迫下白木通幼苗叶片的电子传递速率ETR、Fv/Fm及Fv/Fo均显著降低,说明干旱胁迫使白木通PSII的活性降低,电子传递速率受阻,最终导致光合效率降低。本研究主要从光合生理指标方面探讨了干旱胁迫对白木通幼苗的影响,轻度干旱并未对白木通的生长产生任何影响,说明轻度干旱有可能提高了土壤透气度,使得根系能够更好地呼吸。中度干旱和重度干旱均对白木通的生长产生影响,且重度干旱对白木通的影响要明显大于中度干旱,由以上分析可知,中度干旱只是由于气孔关闭引起的光合效率降低引起的,在复水后可以恢复到正常的生长状态,而重度干旱是破坏了白木通叶片PSII光合机构活性的非气孔因素引起的,对白木通的生长有显著影响。本文通过盆栽试验探讨了干旱胁迫对白木通光合生理指标的影响,对大田试验还需以后进一步的研究。
[1]Bailey RG. Ecosystem Geography with Separate Maps of the Oceans and Continents at 1:8000000 [M].New York: Springer-Verlag, 1996.
[2]陈 珂, 蒋 祺, 类延宝,等. 植物对干旱胁迫的生理响应[J].安徽农业科学, 2009, 37 (5): 1907-1908.
[3]胡 义, 胡庭兴, 胡红玲,等. 干旱胁迫对香樟幼树生长及光合特性的影响[J].应用与环境生物学报, 2014,20(4):675-682.
[4]Levitt J. Responses of plants to environmental stresses. Volume II. water, radiation, salt and other stresses[M]. Academic Press,1980, 607.
[5]曹 慧, 许雪峰, 韩振海,等. 水分胁迫下抗旱性不同的两种苹果属植物光合特性的变化[J].园艺学报,2004,31(3):285-290.
[6]国家药典委员会. 中华人民共和国药典一部[M].北京化学工业出版社, 2005.
[7]李 丽, 陈绪中, 姚小洪, 等. 三种木通属植物的地理分布与资源调查[J].武汉植物学研究, 2010, 28 (4): 497-506.
[8]高慧敏, 王智民. 白木通中一个新的三萜皂苷类化合物[J].药学学报, 2006, 41(9): 835-839.
[9]柳 方. 关木通与白木通的来源与鉴别[J].新疆中医药, 2004,22(2): 35-36.
[10]鄢帮有, 张时煌, 刘梅影,等. 新型藤本油料植物白木通的开发与利用[J].江西农业学报, 2014, 26(9): 20-22.
[11]刘金花, 李 佳, 张永清. 黄芩植株光合特性初步研究[J].中草药, 2009, 40(6): 961-964.
[12]李中勇, 张 媛, 韩龙慧,等. 氮钙互作对设施栽培油桃叶片光合特性及叶绿素荧光参数的影响[J].植物营养与肥料学报,2013, 19(4):893-900.
[13]张丽军, 赵领军, 赵善仓.干旱胁迫对苹果光合特性的影响[J].河北果树,2007, (4):3-5.
[14]王振兴, 陈 丽, 艾 军,等.不同干旱胁迫对山葡萄的光合作用和光系统II活性的影响[J].植物生理学报, 2014, 50(8):1171-1176.
[15]孙景宽, 李 田, 夏江宝, 等.干旱胁迫对沙枣幼苗根茎叶生长及光合色素的影响[J].水土保持通报, 2011, 31(1):68-71.
[16]王智威, 牟思维, 闫丽丽, 等. 水分胁迫对春播玉米苗期生长及其生理生化特性的影响[J].西北植物学报, 2013, 33(2):0343-0352.
[17]Mauro Centritto1, Federico Brilli, Roberta Fodale, et al. Different sensitivity of isoprene emission, respiration and photosynthesis to high growth temperature coupled with drought stress in black poplar (Populus nigra) saplings [J]. Tree Physiology .2011, 31,275-286.
[18]张宪政.植物叶绿素含量测定-丙酮乙醇混合液法[J].辽宁农业科学, 1986, (3): 26-28.
[19]Arnon DI. Copper enzymes in isolated chloroplasts.Polyphenoloxidase in Beta vulgaris[J]. Plant Physiol., 1949, 24:1-15.
[20]马冬云, 郭天财, 宋 晓, 等. 施氮对冬小麦旗叶RuBP羧化酶活性及叶绿素荧光参数的影响[J].西北植物学报, 30(11):2197-2202.
[22]Roháček K. Chlorophyll fluorescence parameters: the definitions photosynthetic meaning and mutual relationships[J].Photosynthetica, 2002, 40(1): 13-29.
[23]吴雪霞, 杨晓春, 朱宗文, 等. 外源6-BA对低温胁迫下茄子幼苗光合作用、叶绿素荧光参数及光能分配的影响[J].植物生理学报, 2013,49 (11): 1181-1188.
[24]Bilger W, Bjorkman O. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis[J]. Photosynthesis Research, 1990,5(3):173-185.
[25]张守仁. 叶绿素荧光动力学参数的意义及讨论[J].植物学通报, 1999, 16(4): 444-448.
[26]宋丽萍, 蔡体久, 喻晓丽. 水分胁迫对刺五加幼苗光合生理特性的影响[J].中国水土保持科学, 2007, 5(2): 91-95.
[27]付秋实, 李红岭, 崔 健, 等. 水分胁迫对辣椒光合作用及相关生理特性的影响[J].中国农业科学,2009,42(5):1859-1866.
[28]Farquhar G D. Sharkey T D. Stomatal conductance and photosynthesis. Annual Review of Physiology, 1982,33:317-345.
[29]王旭军,潘百红,程 勇,等.红榉不同种源光合特性的比较[J].中南林业科技大学学报,2013,33(6):37-42.
[30]蒋 燚,李志辉,朱积余,等.不同红锥种源幼林光合特性日变化测定与分析[J].中南林业科技大学学报,2013,33(6):43-47.
[31]Peterson R B, Aivak M N, Walker D A. Relationship between steady-state fluorescence yield and photosynthetic efficiency in spinach leaf issue[J]. Plant Physiology, 1998, 88: 158-163.
Effects of drought stress on photosynthetic physiological characteristics of Akebia trifoliate
WU Ling-li1, LEI Xiao-lin2, GONG Chun2, KE Bin-feng1, XIONG Li1, LI Jian-an1
(1. Key Lab. of Cultivation and Protection for Non-Wood Forest Trees Co-constructed by China Education Ministry and Central South University of Forestry and Technology, Changsha 410004, Hunan, China; 2. Jiangxi Province Academy of Forestry, Nanchang 330013,Jiangxi, China)
In order to provide theoretical basis for different water and fertilizer managements and drought resistance cultivation, potted culture experiments were conducted to study the physiological and biochemical responses and chlorophyll fluorescence parameters to reveal the effect of drought stress on photosynthesis and associated physiological characters of Akebia trifoliate. The results show that there was no significant effect on A.trifoliate besides that the water use efficiency(RWUE) was increased under light drought stress;When under modest drought stress, the chlorophyll content, net photosynthetic rate(Pn), transpiration rate(Tr), intercellular CO2concentration(Ci), electron transfer rate(RETR), the maximum photochemical efficiency(Fv/Fm), Fv/Foand qPdecreased significantly(P < 0.05) while RuBP carboxylase activity, stomatal conductance(Gs), ΦPSIIand Fm had no significant changes; When under severe drought stress, the chlorophyll content, RuBP carboxylase activity, net photosynthetic rate(Pn), transpiration rate(Tr), stomatal conductance(Gs), electron transfer rate(RETR), the maximum photochemical efficiency(Fv/Fm), ΦPSII, Fv/Fo, Fm, and qPdecreased significantly (P<0.05), while intercellular CO2concentration(Ci), water use efficiency(RWUE), Foand CNPQincreased significantly(P<0.05). Under moderate drought stress, the photosynthesis reduce of A.trifoliate seedlings was primarily caused by stomatal closure while the decrease of net photosynthetic rate was considered as a result of non-stomatal factors under severe drought stress condition.
Akebia trifoliate var. australis; drought stress; photosynthetic characteristics; RuBP carboxylase; chlorophyll fluorescence
S718.43
A
1673-923X(2015)11-0068-06
10.14067/j.cnki.1673-923x.2015.11.013
2015-01-10
国家林业公益性行业科研专项 (201304802)
吴玲利,硕士研究生
李建安,教授,博士;E-mail:lja0731@126.com
吴玲利,雷小林,龚 春,等.干旱胁迫对白木通光合生理特性的影响[J].中南林业科技大学学报, 2015, 35(11): 68-73.
[本文编校:吴 毅]
猜你喜欢
贵州农业科学(2022年4期)2022-05-06
中草药(2022年7期)2022-04-01
干旱地区农业研究(2022年1期)2022-01-28
现代盐化工(2021年3期)2021-08-27
食品与生物技术学报(2020年4期)2020-01-06
装备制造技术(2019年12期)2019-12-25
科技创新与品牌(2019年12期)2019-02-06
现代职业教育·中职中专(2017年12期)2017-08-13
新课程·下旬(2015年10期)2015-10-21
中国民族民间医药·下半月(2014年5期)2014-12-02
