
锌对1~4周龄鹅生长性能、免疫与抗氧化功能及金属硫蛋白-Ⅰ mRNA表达量的影响
2015-12-20 07:37王宝维陈苗璐王秉翰张名爱葛文华
动物营养学报 2015年4期
王宝维 陈苗璐 王秉翰 张名爱 葛文华 岳 斌
(1.国家水禽产业技术体系营养与饲料研究室,青岛 266109;2.青岛农业大学优质水禽研究所,青岛 266109;3.林肯大学,克莱斯特彻奇 85084,新西兰)
锌是动物所必需的微量元素之一,随着人们对锌认识的不断增加,锌在动物体内的转运和作用逐渐被揭开。研究表明,锌不仅对促进生长,维持免疫器官的形态、结构和功能具有重要作用,而且能够防止细胞膜氧化、减少超氧阴离子形成,有效地保护机体免受自由基损伤,改善机体健康状态[1-3]。锌还能诱导金属硫蛋白(metallothionein,MT)的合成,从而调控其基因的表达。饲粮中锌的缺乏或过量都会对动物产生不良影响,严重时甚至发病死亡。为此,开展锌适宜营养需要量研究,探讨其对机体免疫与抗氧化功能和MT-Ⅰ mRNA表达量的影响及相互关系,对科学地确定鹅饲粮中锌的需要量具有重要意义。黄艳玲等[4]通过对肉仔鸡饲粮锌适宜水平研究,推荐肉仔鸡饲粮锌的适宜水平为80 mg/kg。孙淑霞等[5]和卢昊等[6]研究表明,饲粮中添加锌可以提高动物的免疫器官指数。虞泽鹏等[7]报道,饲粮添加锌能显著提高血清铜锌超氧化物歧化酶(CuZn-SOD)和谷胱甘肽过氧化物酶(GSH-Px)活性,从而提高机体抗氧化功能。Cousins[8]发现锌诱导MT合成主要是通过调节其转录水平实现的。陈苗璐等[9]研究建议,1~4周龄鹅锌适宜水平为106.05 mg/kg。上述研究结果表明,锌对动物生长发育和免疫机能具有一定调控作用,并且饲粮中锌水平差异区间较宽。迄今为止,锌对1~4周龄鹅免疫与抗氧化功能的影响尚未进行系统全面的研究,对MT-ⅠmRNA表达量的研究处于空白。为此,本试验以1~4周龄青农灰鹅(大型鹅)为试验对象,通过探讨不同水平锌对其生长性能、免疫与抗氧化功能和MT-ⅠmRNA表达量的影响以及相关性分析,确定在最佳生长性能、免疫与抗氧化功能下饲粮中锌的适宜水平。
1 材料与方法
1.1 试验动物与试验设计
选用1日龄体重相近健康青农灰鹅(法国米朗德、阿蒂盖品种杂交配套选育而成)360只,设计6个组(Ⅰ组为对照组,Ⅱ~Ⅵ组为试验组),每组6个重复,每个重复10只鹅(公母各占1/2)。各组饲 粮锌水平分别为 31.28、81.28、131.28、181.28、231.28、281.28 mg/kg(含基础饲粮中锌含量);各组硫酸锌(ZnSO4)添加量分别为0、228.79、457.58、686.37、951.16、1 143.96 mg/kg。试验鹅由国家水禽产业技术体系育种基地高密银河润雁鹅业有限公司提供。
1.2 试验饲粮
基础饲粮参照NRC(1994)建议鹅的营养需要量[10]和《中国饲料成分及营养价值表》进行配方设计,基础饲粮组成及营养水平见表1。试验用硫酸锌购自浙江新维普添加剂有限公司,其有效成分为98%。

表1 基础饲粮组成及营养水平(风干基础)Table 1 Composition and nutrient levels of the basal diet(air-dry basis) %
1.3 饲养管理
试验前对鹅舍进行全面消毒。试验鹅全期舍饲,地面平养,自由饮水和采食,饲粮少喂勤添;随时观察鹅群的生长状况;分别于7和34日龄接种鹅副黏病毒疫苗。
1.4 样品的采集与制备
4周龄末,分别从每个重复中抽取4只,共144只(公母各占 1/2),采用颈静脉采血,3 000 r/min离心 l0 min,得浅黄色血清样品于-80℃冰箱冻存。放血处死后取部分肝脏样品,并按照组织匀浆的制备方法将肝脏制成10%的组织匀浆液(组织∶生理盐水=1∶9)。样品于 -80℃冰箱冻存备用。取部分肝脏样品迅速放入液氮中保存,用以分析肝脏中MT-Ⅰ mRNA表达量。其他试验鹅进行生长性能和屠宰性能测定。
1.5 测定指标及方法
1.5.1 免疫指标的测定
摘取试验鹅的免疫器官(胸腺、脾脏和法氏囊),用滤纸擦除血渍后称重,用以计算免疫器官指数。禽流感抗体效价的测定根据病毒的血凝(HA)和血凝抑制试验(HI)进行。
1.5.2 抗氧化指标的测定
总抗氧化能力(T-AOC)采用比色法测定;过氧化氢酶(CAT)活性采用可见光分光光度测定;GSH-Px活性采用比色法测定;CuZn-SOD活性采用黄嘌呤氧化酶法测定;丙二醛(MDA)含量采用硫代巴比妥酸反应(TBA)法测定;蛋白质含量采用考马斯亮蓝法测定。上述指标均用试剂盒测定,试剂盒均购自南京建成生物工程研究所。
1.5.3 MT-Ⅰ mRNA 表达量的测定
1.5.3.1 引物设计
MT-Ⅰ和肌动蛋白(β-actin)引物由信诺金达(北京)有限公司设计并合成,引物序列见表2。
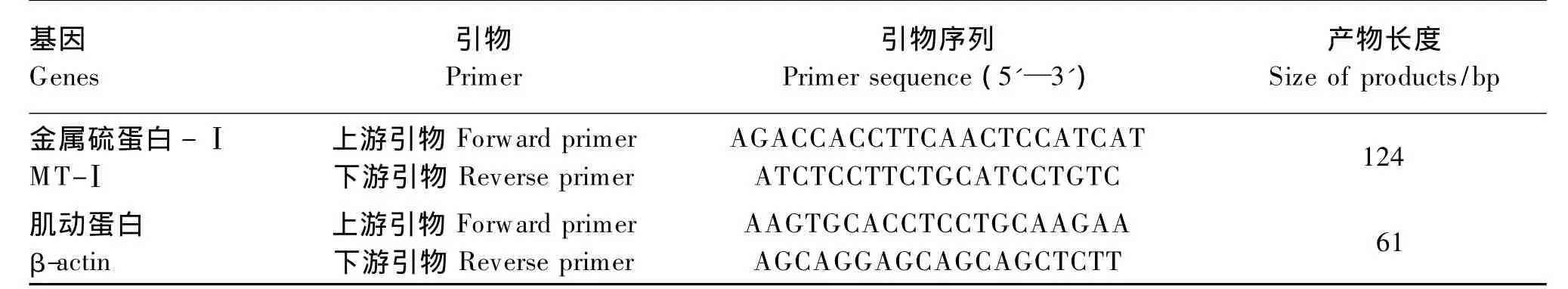
表2 MT-Ⅰ和β-actin基因引物序列Table 2 Primer sequence of MT-Ⅰ and β-actin
1.5.3.2 MT-Ⅰ mRNA 表达量的测定方法
采用Omega总RNA试剂盒提取肝脏组织中的总RNA,用紫外分光光度计测定总RNA质量浓度,并在260和280 nm下测吸光度,计算OD260/OD280的值。此值在1.7~2.0,说明提取的 RNA纯度较高,适宜进行下一步的试验。取各个样品总RNA 1μg,在20μL混合液体系统中进行反转录,合成 cDNA(65℃ 10 min,42℃ 60 min,70℃5 min)。qPCR反应体系按照10μL qPCR Mix(K0222,Fermentas),0.6 μL Forward/Reverse primer(5 μmol/L),1 μL cDNA 和 8.2 μL ddH2O进行配制,在定量PCR仪(Applied Biosystems Step One PlusTM)上进行反应。反应程序为:95℃10 min;45个循环的95℃ 15 s,60℃ 45 s;最后按照仪器默认添加熔解程序:95℃ 15 s,60℃ 20 s,95 ℃ 15 s。
1.6 统计分析
试验数据以重复为单位进行数据处理,采用SPSS 17.0软件中单因素方差分析(one-way ANOVA)中的LSD法进行,平均值的多重比较采用Duncan氏法进行。试验数据以“平均值±标准差”表示。利用二次曲线和渐近线模式对相应的数据进行拟合,以确定鹅饲粮中锌的适宜水平。采用比较Ct方法分析MT-Ⅰ mRNA表达量的数据。P<0.05和P<0.01分别为差异显著和极显著水平。
2 结果与分析
2.1 锌对鹅生长性能的影响
由表3可见,4周龄时,与Ⅰ组相比,Ⅱ组平均日增重(ADG)显著增加(P<0.05),Ⅱ、Ⅲ组的平均日采食量(ADFI)显著增加(P<0.05);料重比(F/G)、死淘率各组间差异均不显著(P>0.05);Ⅳ、Ⅴ、Ⅵ组间 ADG、ADFI均无显著差异(P>0.05),说明饲粮锌水平超过 181.28 mg/kg时,对鹅的 ADG、ADFI、F/G 均无显著影响(P >0.05)。以Ⅰ~Ⅳ组ADG、ADFI和F/G进行曲线拟合,得到如下曲线方程:

结果表明,饲粮锌水平为112.51 mg/kg时ADG最大。

结果表明,饲粮锌水平为106.05 mg/kg时F/G最低。
ADFI与锌水平间的曲线关系不明显(R2<0.700)。
结果表明,根据生长性能分析鹅饲粮锌适宜水平为 112.51 mg/kg。

表3 锌对鹅生长性能的影响Table 3 Effects of zinc on growth performance of geese
2.2 锌对鹅免疫器官指数的影响
由表4可知,胸腺指数Ⅱ组极显著高于Ⅰ组(P<0.01),其他各组均显著高于Ⅰ组(P<0.05);脾脏指数Ⅱ、Ⅲ组极显著高于Ⅰ组(P<0.01),其他各组均显著高于Ⅰ组(P<0.05);法氏囊指数各组间差异不显著(P>0.05),Ⅲ组最高。免疫器官指数与锌水平间的曲线关系不明显(R2<0.700)。
结果表明,饲粮锌水平在 81.28~131.28 mg/kg时,对胸腺指数、脾脏指数有显著影响。

表4 锌对鹅免疫器官指数的影响Table 4 Effects of zinc on immune organ indices of geese mg/g
2.3 锌对鹅抗体效价的影响
由表5可得,鹅副黏病毒疫苗免疫前鹅的抗
体效价各组间差异不显著(P>0.05);免疫后7 d,Ⅳ组抗体效价显著高于Ⅰ组(P<0.05),提高了15.76%;免疫后14 d,Ⅲ、Ⅳ组抗体效价显著高于Ⅰ组(P<0.05),均提高了15.09%。通过曲线拟合分析,发现免疫后14 d抗体效价(Y)与饲粮锌水平间(X)呈现渐近线关系,建立如下方程:Y=6.248 -1.87e -0.029X(R2=0.853,P=0.001)。由公式得出,当饲粮锌水平为157.41 mg/kg时,免疫后14 d的抗体效价最高。

表5 锌对鹅抗体效价的影响Table 5 Effects of zinc on antibody titer of geese log2
2.4 锌对鹅抗氧化指标的影响
2.4.1 锌对鹅血清及肝脏中T-AOC的影响
由表6可见,4周龄时,Ⅳ组血清T-AOC极显著高于Ⅰ组(P<0.01),Ⅲ组显著高于Ⅰ组(P<0.05);Ⅱ、Ⅲ组肝脏 T-AOC显著高于Ⅰ组(P<0.05)。T-AOC与饲粮锌水平间的曲线关系不明显(R2<0.700)。结果表明,饲粮锌水平为 81.28~131.28 mg/kg时,对血清及肝脏中T-AOC有显著影响。
2.4.2 锌对鹅血清及肝脏中CAT活性的影响
由表6可见,4周龄时,Ⅱ、Ⅲ、Ⅳ组血清CAT活性极显著高于Ⅰ组(P<0.01),Ⅴ组显著高于Ⅰ组(P<0.05);Ⅱ、Ⅲ组肝脏CAT活性极显著高于Ⅰ组(P<0.01),Ⅳ组显著高于Ⅰ组(P <0.05),以Ⅱ组活性最高。CAT活性与饲粮锌水平间的曲线关系不明显(R2<0.700)。结果表明,饲粮锌水平为 131.28~181.28 mg/kg时对血清和肝脏CAT活性有显著影响。
2.4.3 锌对鹅血清及肝脏中GSH-Px活性的影响
由表6可见,4周龄时,Ⅱ、Ⅲ、Ⅳ组血清GSHPx活性极显著高于Ⅰ组(P<0.01),Ⅴ组显著高于Ⅰ组(P<0.05),以Ⅳ组活性最高;Ⅳ、Ⅴ组肝脏GSH-Px活性极显著高于Ⅰ组(P<0.01),Ⅱ、Ⅲ组显著高于Ⅰ组(P<0.05),以Ⅳ组活性最高。通过曲线拟合分析可得,血清GSH-Px活性(Y)与饲粮锌水平(X)间呈二次曲线关系,建立的回归方程如下:Y=0.002X2+0.679X+45.497(R2=0.720,P=0.003)。结果表明,适宜饲粮水平锌能显著提高血清和肝脏中GSH-Px活性,当饲粮锌水平为160.66 mg/kg时,血清GSH-Px活性最高。
2.4.4 锌对鹅血清及肝脏中CuZn-SOD活性的影响
由表6可见,4周龄时,Ⅳ组血清 CuZn-SOD活性极显著高于Ⅰ组(P<0.01),Ⅲ组显著高于Ⅰ组(P<0.05);Ⅳ组肝脏CuZn-SOD活性极显著高于Ⅰ组(P<0.01),Ⅲ、Ⅴ组显著高于Ⅰ组(P<0.05)。CuZn-SOD活性与饲粮锌水平间的曲线关系不明显(R2<0.700)。结果表明,饲粮锌水平为131.28 ~181.28 mg/kg时对血清和肝脏 CuZn-SOD活性有显著影响。
2.4.5 锌对鹅血清及肝脏组织中MDA含量的影响
由表6可见,4周龄时,随饲粮锌水平的升高,血清MDA含量呈现先降低后升高的趋势,Ⅱ、Ⅲ组极显著低于Ⅰ组(P<0.01),Ⅳ、Ⅴ组显著低于Ⅰ组(P<0.05)。Ⅵ组肝脏MDA含量显著高于Ⅰ组(P <0.05),Ⅱ、Ⅲ组间差异显著(P <0.05),以Ⅲ组最低。
通过曲线拟合分析可得,4周龄血清MDA含量(Y)与饲粮锌水平(X)间呈二次曲线关系,建立回归方程如下:Y=2.377 ×10-5X2+0.008X+0.522(R2=0.708,P=0.003)。结果表明,当饲粮锌水平为150.90 mg/kg时,血清MDA含量最低。

表6 锌对鹅血清及肝脏中抗氧化指标的影响Table 6 Effects of zinc on antioxidant parameters in serum and liver of geese
2.5 锌对肝脏中MT-ⅠmRNA表达量的影响
由表8可知,随饲粮锌水平的增加,肝脏中MT-ⅠmRNA表达量呈先升高后降低趋势。与Ⅰ组相比,Ⅳ组显著提高(P <0.05),Ⅱ、Ⅲ组极显著提高(P<0.01),以Ⅲ组表达量最高。结果表明,饲粮适宜锌水平能够显著影响肝脏MT-ⅠmRNA表达量。

表8 锌对鹅肝脏中MT-ⅠmRNA表达量的影响Table 8 Effects of zinc on liver MT-ⅠmRNA expression of geese
2.6 肝脏中MT-ⅠmRNA表达量与抗氧化能力的相关性分析
由表9可知,鹅肝脏中MT-Ⅰ mRNA表达量与血清和肝脏中T-AOC活性呈正相关(P>0.05),与 CAT 活性呈显著正相关(P <0.05),与GSH-Px活性呈正相关(P>0.05),与 CuZn-SOD活性呈正相关(血清 P <0.05,肝脏 P >0.05),与MDA含量呈负相关(血清 P<0.05,肝脏 P>0.05)。结果表明,MT-Ⅰ mRNA表达量与抗氧化能力指标密切相关。饲粮锌水平能够影响MT-ⅠmRNA的表达量,从而调控机体抗氧化能力。

表9 肝脏中MT-ⅠmRNA表达量与抗氧化能力的相关性分析Table 9 Correlation analysis between antioxidant capacity and liver MT-ⅠmRNA expression of geese
3 讨论
3.1 鹅饲粮中锌的需要量
畜禽饲养标准中营养需要量是指动物在最适宜环境条件下,正常、健康生长或达到理想生产成绩对各种营养物质种类和数量的最低要求。NRC(1994)饲养标准中鹅锌的推荐量为:1~4周龄40 mg/kg,4周龄以后 35 mg/kg,种鹅65 mg/kg;澳大利亚饲养标准中鹅锌的推荐量为60 mg/kg;我国农业部1124公告中规定,鹅饲粮中锌的允许量为60 mg/kg;经检索上述推荐量没有相关试验研究数据做支撑,也没有相关研究报道。另外,国内外同行们普遍认为,推荐量只是一个最低维持需要量,而不是发挥最佳生长性能状态的需要量。本试验结果表明,饲粮中锌水平在 81.28~181.28 mg/kg区间内,生长性能、免疫与抗氧化功能和MT-ⅠmRNA表达量均处于高位状态,比国内外有关锌水平对家禽生长性能和屠宰性能影响的研究报道推荐量要高,分析其原因可能有以下几方面:一是与饲养品种类型有关,本试验采用的是利用大型朗德鹅选育的肝用型鹅,体型大,生长速度快;二是与试验期间夏季高温季节有关,本试验在6~9月份,平均温度在26~34℃;三是可能是机体达到最佳生长性能不一定是最佳免疫和抗氧化状态,其有关机理还有待于继续研究。另外,最佳生长性能状态下锌与微量元素间的协同和拮抗关系也需要深入研究。
3.2 锌对鹅免疫器官指数的影响
胸腺、脾脏和法氏囊是家禽主要的免疫器官。其中,胸腺和法氏囊属于中枢免疫器官,对机体的细胞免疫和体液免疫都发挥着重要作用。胸腺能够释放胸腺素,而胸腺素发挥生理作用需要锌的参与。法氏囊对B淋巴细胞的成熟有重要影响,它在家禽性成熟前达到最大,后逐渐萎缩直至消失。脾脏是机体的外周免疫器官,它对血液中的抗原物质具有滤过、清除的作用,是进行免疫应答反应的重要器官。Fraker等[11]研究表明,饲喂低锌(0.5μg/g)饲粮的成年大鼠,其胸腺明显萎缩。崔艳红等[12]研究发现,当锌添加水平达100 mg/kg时,肉用仔鸡(21、42日龄)免疫器官指数达到最高值。
本试验结果表明,胸腺、脾脏和法氏囊指数均随饲粮锌水平的升高呈现先升高后降低趋势。饲粮中加锌能显著提高胸腺和脾脏指数,以饲粮锌水平为81.28~131.28 mg/kg时,鹅的各项免疫器官指数最大,与崔艳红等[12]研究结果相近似,说明锌能有效的促进胸腺、脾脏和法氏囊的发育。
3.3 锌对鹅抗体效价的影响
免疫抗体效价是反映动物体液免疫水平的重要指标,可以直接反映家禽体内的免疫状态。对预防传染病的发生具有重要的意义。张日俊等[13]研究发现,肉仔鸡缺锌能降低体液免疫水平。本试验结果表明,鹅副黏病毒疫苗免疫后7 d,Ⅳ组抗体效价显著提高;免疫后14 d,Ⅲ、Ⅳ组抗体效价显著提高。饲粮中锌水平为157.41 mg/kg时,免疫后14 d的抗体效价最高,这也与闫素梅等[14]的研究结果相似。
3.4 锌对鹅抗氧化功能的影响
动物的健康程度与机体防御体系的抗氧化能力有着密切的联系。T-AOC是体内酶促抗氧化体系的重要组成部分,能清除机体不断产生的氧自由基,从而维持机体的代谢平衡。冯望宝等[15]报道,低锌显著降低蛋鸭T-AOC,补锌后T-AOC逐渐增加,但在高锌(1 000 mg/kg)条件下,T-AOC受到抑制。本试验结果与上述研究相似,表明锌能够显著提高动物体内T-AOC,并且随锌水平的升高T-AOC呈现先升高后降低趋势。血清T-AOC为Ⅳ组最高,肝脏T-AOC为Ⅱ组最高。
CAT是以锌为活性中心的一种酶类清除剂,它能催化过氧化氢(H2O2)分解为水和氧,消除H2O2对机体的伤害,在机体防御体系中发挥重要作用。钟映梅等[16]研究发现,随着饲粮锌添加量的增加,血清CAT活性有升高的趋势,55日龄缺锌与高锌组血清CAT活性均显著下降。本试验结果表明,锌能显著提高血清和肝脏中的CAT活性,CAT活性随锌水平的升高呈现先升高后降低趋势;饲粮锌水平在 131.28 ~181.28mg/kg时,鹅血清和肝脏中的CAT活性最高。
GSH-Px是机体内广泛存在的一种重要的含锌酶。主要参与还原型谷胱甘肽对H2O2的还原反应,能够保护细胞膜等结构和功能的完整,从而防止脂质过氧化,提高机体的抗氧化能力。CuZn-SOD是一种锌依赖酶,主要功能是使超氧自由基(O2-·)发生歧化反应,消除O2-·毒性,防止机体过氧化。蒋宗勇等[17]研究发现,饲粮添加锌显著提高黄羽肉鸡血清GSH-Px和CuZn-SOD活性,以添加120 mg/kg锌达到最高值;饲粮添加锌对肝脏中CuZn-SOD活性基本无影响。本试验结果表明,饲粮中添加锌能显著提高血清和肝脏中GSH-Px活性;血清和肝脏中CuZn-SOD活性均随饲粮锌水平的升高呈先升高后降低的趋势,血清和肝脏中CuZn-SOD活性均以Ⅳ组最高。
氧自由基能够攻击生物膜中的多不饱和脂肪酸,引起脂质过氧化反应,并形成脂质过氧化物MDA等。MDA含量能间接反映生物体内氧自由基的代谢状况以及脂质过氧化程度。Sahin等[18]报道指出,锌能降低家禽血清和肝脏中MDA含量。钟映梅等[19]的研究表明,当饲粮锌水平为110 mg/kg时,鹅血清MDA和肝脏MDA含量较低。本试验结果表明,鹅血清和肝脏中MDA含量以Ⅲ组最低,表明适宜水平锌能够有效地降低体内自由基水平。
3.5 锌对鹅肝脏中MT-Ⅰ mRNA表达量的影响
MT是一种低分子质量、半胱氨酸富集的金属结合蛋白,于1957年在马的肾脏中首次发现。现已证实,MT具有广泛的生物学功能,对机体的调节和代谢具有重要影响[20]。MT具有 MT-Ⅰ、MT-Ⅱ、MT-Ⅲ 和 MT-Ⅳ 4 种 异 构 体。其 中,MT-Ⅰ主要存在于动物的肝脏中。Wei等[21]报道,饲粮添加或注射锌均可使雏鸡组织 MT-ⅠmRNA水平升高。曹家银等[22]研究表明,饲粮中添加200 mg/kg锌可提高肉仔鸡肝脏MT mRNA水平。本研究结果表明,添加锌能使鹅肝脏中MT-ⅠmRNA表达量快速上升;饲粮锌水平高于181.28 mg/kg时,MT-Ⅰ mRNA表达量急速下降,且处于稳定状态。以131.28 mg/kg锌水平的MT-ⅠmRNA表达量最高。
3.6 肝脏中MT-ⅠmRNA表达量与抗氧化能力的相关性分析
MT是一种富含半胱氨酸(20% ~30%)的蛋白质。因为在自然状态下,半胱氨酸的巯基(-SH)多以还原状态存在,具有很强亲核性,从而使MT对O2-·、羟自由基(·OH)以及一氧化氮(NO)等自由基产生作用,达到保护机体的目的。Jourdan等[23]研究表明,锌的抗氧化作用与其诱导MT合成有关。刘湘新等[24]发现MT能够提高SOD活性,防止氧化性应激损伤。李峰等[25]报道,运动大鼠补充MT后,血清抗氧化酶活性呈上升趋势。本试验结果表明,鹅肝脏中 MT-ⅠmRNA表达量与血清和肝脏CAT活性和血清中CuZn-SOD活性呈显著正相关,与血清中MDA含量呈显著负相关。有关锌影响MT-Ⅰ mRNA调控鹅抗氧化能力的机理还有待于继续研究。
4 结论
① 饲粮锌水平为112.51 mg/kg时平均日增重最大,饲粮锌水平为106.05 mg/kg时料重比最低。
② 饲粮锌水平在81.28~181.28 mg/kg时,机体免疫和抗氧化力处于高位水平;显著提高机体免疫器官指数和鹅副黏病毒疫苗免疫后抗体效价水平;显著提高机体抗氧化功能,降低自由基含量。
③ 机体MT-Ⅰ mRNA表达量与机体抗氧化能力指标密切相关。适宜锌水平能够影响MT-ⅠmRNA的表达量,从而调控机体抗氧化能力。
④高位免疫与抗氧化力状态下锌的需要量要比最佳生长性能状态锌的需要量高。锌水平过高生长性能免疫与抗氧化功能开始下降。
[1] 韩新燕,郭勇.微量元素锌与动物营养[J].饲料博览,2000(10):26-27.
[2] 李大刚,王宏,周桂莲.微量元素锌的抗氧化作用研究[J].饲料研究,2008(11):38-41.
[3] 丁小波,文利新.微量元素锌与动物免疫[J].微量元素与健康研究,2006,23(3):55 -57.
[4] 黄艳玲,吕林,李素芬,等.0~21日龄肉仔鸡饲粮中锌适宜水平研究[J].畜牧兽医学报,2008,39(7):900-906.
[5] 孙淑霞,赵晶,李玉杰,等.蛋氨酸螯合锌对绿头野鸭免疫器官指数及血液指标的影响[J].黑龙江畜牧兽医,2008(9):42-43.
[6] 卢昊,王春维,周海,等.谷氨酸锌对肉仔鸡生长性能、血清含锌酶活性及免疫器官指数的影响[J]中国饲料,2010(1):24-26.
[7] 虞泽鹏,乐国伟,施用晖,等.不同锌源对断奶小鼠生长及机体抗氧化能力的影响[J].畜牧与兽医,2005,37(4):1 -3.
[8] COUSINS R J.Absorption,transport,and hepatic metabolism of copper and zinc:special reference to metallothionein and ceruloplasmin[J].Physiological Reviews,1985,65(2):238 -309.
[9] 陈苗璐,王宝维,张名爱,等.饲粮锌水平对鹅生长性能、血清生化指标及激素含量的影响[J].动物营养学报,2013,25(5):1105 -1112.
[10] NRC.Nutrient requirements of poultry[M].9th ed.Washington,D.C.:The National Academy Press,1994.
[11] FRAKER P J,HAAS S M,LUECKE R W.Effect of zinc deficiency on the immune response of the young adult A/J mouse[J].The Journal of Nutrition,1977,107(10):1889-1895.
[12] 崔艳红,刘保国,王艳荣,等.不同水平的蛋氨酸锌对肉用仔鸡生产性能和免疫的影响[J].饲料工业,2009,30(2):34 -36.
[13] 张日俊,周毓平,黄燕,等.锌对肉仔鸡免疫器官生长发育及免疫功能调节作用的研究[J].畜牧兽医学报,1999,30(6):504 -512.
[14] 闫素梅,郝永清,史彬林,等.日粮锌水平对肉仔鸡组织锌浓度及其生产性能与免疫机能的影响[J].饲料工业,2002,23(12):25 -27.
[15] 冯望宝,王安,艾涛.不同锌水平对笼养育成蛋鸭生长性能及总抗氧化能力的影响[J].东北农业大学学报,2007,38(5):654 -659.
[16] 钟映梅,孙刚,张俊峰,等.不同锌水平对鹅血液抗氧化酶活性的影响[J].黑龙江畜牧兽医,2007(3):44-45.
[17] 蒋宗勇,刘小雁,蒋守群,等.43~63日龄黄羽肉鸡锌需要量的研究[J].中国农业科学,2010,43(20):4295-4302.
[18] SAHIN K,SMITH M O,ONDERCI M,et al.Supplementation of zinc from organic or inorganic source improves performance and antioxidant status of heat-distressed quail[J].Poultry Science,2005,84(6):882 -887.
[19] 钟映梅,王力强,孙刚,等.锌对鹅铜锌超氧化物歧化酶和丙二醛含量影响变化的研究[J].现代畜牧兽医,2007(3):11-13.
[20] MARGOSHESM,VALLEE B L.A cadmium protein from equine kidney cortex[J].Journal of the American Chemical Society,1957,79(17):4813 -4814.
[21] WEI D Y,ANDREWS G K.Molecular cloning of chicken metallothionein.Deduction of the complete amino add sequence and analysis of expression using cloned cDNA[J].Nucleic Acids Research,1988,16(2):537-553.
[22] 曹家银,罗绪刚,DAVIS S R,等.以组织锌、金属硫蛋白及其基因表达指标评价肉仔鸡对锌源的相对生物学利用率[J].畜牧兽医学报,2003,34(3):227-231.
[23] JOURDAN E,EMONET-PICCARDI N,DIDIER C,et al.Effects of cadmium and zinc on solar-simulated light-Ⅰrradiated cells:potential role of zinc-metallothionein in zinc-Ⅰ nduced genoprotection[J].Archives of Biochemistry and Biophysics,2002,405(2):170-177.
[24] 刘湘新,李丽立,刘进辉,等.金属硫蛋白对应激猪血清中某些酶活性的影响[J].中国兽医杂志,2005,41(12):7 -8.
[25] 李峰,石辉.锌金属硫蛋白对PM2.5暴露的运动大鼠血清抗氧化酶及免疫指标的影响[J].环境科学学报,32(2):465-471.
猜你喜欢
动物营养学报(2022年8期)2022-09-01
中国饲料(2021年17期)2021-11-02
当代水产(2021年6期)2021-08-13
中成药(2019年12期)2020-01-04
广东饲料(2016年6期)2016-12-01
动物营养学报(2015年9期)2016-01-07
食品工业科技(2014年15期)2014-03-11
食品工业科技(2014年15期)2014-03-11
现代农业科技(2009年19期)2009-03-20
中学生数理化·八年级数学华师大版(2008年3期)2008-08-26
