
云南高原湖泊湿地蚊类多样性空间分布特征及与重要环境因素关系*
2015-11-12 01:32臧颖惠龚正达连宏宇边长玲琚俊科岳仁萍张丽云周红宁
寄生虫与医学昆虫学报 2015年4期
臧颖惠 龚正达,** 连宏宇 边长玲 琚俊科 李 栋 岳仁萍 张丽云 周红宁
(1. 大理大学公共卫生学院,云南大理 671000; 2. 云南省地方病防治所, 云南大理 671000;3. 云南省寄生虫病防治所,云南普洱 665000)
湿地是介于水域生态系统和陆地生态系统之间的一类特殊生态系统,同时也是自然界最富生物多样性的生态景观和人类最重要的生存环境之一(崔保山等,1999)。由于湿地具有稳定环境、物种基因保护及资源利用的功能,被誉为“自然之肾”、“生物基因库”和“人类摇篮”(孙广友,2000)。根据我国湿地资源现状以及《湿地公约》对湿地的分类系统,我国的湿地共分沼泽湿地、湖泊湿地、河流湿地、滨海湿地、人工湿地5大类(赵学敏,2005)。目前,国外对湿地昆虫群落的研究比较全面而深入,开展了区系组成、生物学、生态学以及环境评价等诸多方面的研究(Lathrop,2000; Lawleretal., 2005;Munca,etal., 2005;Stefanescu, 2005; Kati, 2006),而我国在这方面的研究尚处于起步阶段。因此,开展和研究不同地理环境条件中湿地动物地理区系和物种多样性的空间分布状况、特征与规律,对于揭示生物多样性与地理环境之间关系、实施湿地生物多样性的监测和保护都有重要意义。
蚊类(Culicidae)属于双翅目昆虫,幼虫各期孳生于各类型自然或人工水体。成虫不仅侵扰人畜,刺叮吸血,而且是多种虫媒疾病(Vector-borne diseases) 如疟疾、淋巴丝虫病、日本乙型脑炎、登革热等的重要媒介(瞿逢伊,2002;边长玲和龚正达,2009)。近年,国外开展了蚊类物种多样性沿环境梯度的变化规律及其与疾病和重要环境因素等关系的相关研究(Denkeetal.,1996;Kittayapong,etal.,Devietal.,2004;Zhouetal.,2004)。国内有我国西南“三江并流”区域的澜沧江、怒江和金沙江流域蚊类多样性大尺度空间分布格局与环境因素(葛军旗等,2008;张菊仙等,2008;李栋等,2011),和同期进行与虫媒病毒关系等研究(翟本刚等,2008;Sunetal., 2009;陈斌等, 2011;Maetal., 2011)。
根据国家环保部2008年4月公布的全国湖泊湿地动物名录中蚊类只有2亚科4属14种,其中云南仅记载大理洱海湿地分布有按蚊属3种和伊蚊属2种(臧颖慧等, 2009a)。而在生物多样性研究领域中,有关湖泊湿地尤其是高原湖泊湿地蚊类群落生态学及多样性的研究少见报道。鉴于此,我们近年陆续对滇南和滇西高原湖泊部分湿地蚊类多样性开展了一些研究(岳仁萍等,2009;臧颖慧等,2009b)。
本研究以云南省为调查研究的空间,对当地不同环境梯度(纬度和海拔梯度)的重要高原湖泊湿地夜间活动的蚊类多样性进行调查取样和统计分析,在此基础上并对高原湖泊湿地蚊类多样性大尺度空间分布格局的基本规律、特征及其与重要环境因素关系等问题进行研究和探讨。
1 材料与方法
1.1 研究区域
云南省地处中国西南边陲, 地势北高南低。海拔最高点在滇西北滇藏交界的梅里雪山主峰卡格博峰(6 740 m);而最低点是滇南与越南交界的元江出境处(仅为76.4 m),相对高差达6663.6 m,南北跨越约9个纬度(21°N~30°N)。由于地理位置不同和高差大, 由南向北或者由低到高可依次出现亚热带、暖温带和寒温带多种不同的气候带。目前,云南现有蚊类3亚科20属31亚属近300种(亚种)(董学书等,2010),其数量分别约占我国已知属和种类的95%和76%以上,接近世界所知蚊种数的10%,为我国蚊类区系和物种分布和分化的中心(张菊仙和龚正达, 2008)。云南省面积超过 1 km2以上的高原湖泊湿地有30余个(含永久性湖泊湿地23个)。它们主要分布于滇中、滇东南和滇西北地区。因其所处的地貌和海拔高度不同,又分为高原低地中山河谷盆地、亚高山和高山等湖泊湿地类型。它们多是云南省著名和重要的风景旅游区或经济建设区。
以云南省北纬21°08′~ 29°01′;东经97°31′~ 106°11′和海拔500~3 500 m之间的区域作为研究的空间范围,于2007~2009三年间,在雨季、气温和蚊类密度高峰期的7~9月份进行。根据主要天然湖泊的分布情况,从南到北共选择21个天然高原湖泊湿地(永久性湿地)为调查样区,它们是:大屯海、长桥海、异龙湖、普者黑、杞麓湖、星云湖、抚仙湖、滇池、阳宗海、洱海、青海湖、西湖、茈碧湖、剑湖、草海、程海、文笔海、拉市海、泸沽湖、纳帕海、大山包(大海子)。其中,位于滇西北的纳帕海海拔分布最高(3 260 m),而位于滇东南的大屯海和长桥海海拔最低(约1 300 m),各湖泊的地理位置和分布情况详见(图1)。根据云南21个湖泊调查样区所分布和跨越的水平和海拔(约北纬23°~28°N、海拔1 000~3 500 m之间)分布范围,按每纬度(60′)划分水平梯度带,调查区域可划分为5个带, 以Ⅰ-Ⅴ表示,分别代表北纬23°~24°(Ⅰ)、24°~25°(Ⅱ)、25°~26°(Ⅲ)、26°~27°(Ⅳ)、27°~28°(Ⅴ)的取样区间和研究单元;按每500 m划分垂直梯度带, 调查区域可划分为5个带,以A~E表示,分别代表海拔1 000~1 500 m(A)、1 500~2 000 m(B)、2 000~2 500 m(C)、2 500~3 000 m(D)、3 000~3 500 m(E)的取样和数据统计的研究单元。

图1 云南省21个主要高原湖泊湿地的地理位置及分布Fig.1 21 lakes studied in Yunnan Province1大屯海 Datun Lake;2长桥海 Changqiao Lake;3异龙湖 Yilong Lake;4普者黑 Puzhehei Lake;5杞麓湖 Gilu Lake;6星云湖 Xingyun Lake;7抚仙湖 Fuxian Lake;8滇池 Dianchi Lake;9阳宗海 Yangzong Lake;10青海湖 Qinghai Lake; 11洱海 Erhai Lake;12西湖 Xihu Lake;13茈碧湖 Cibi Lake;14 剑湖 Jianhu Lake;15 草海 Caohai Lake;16程海 Chenghai Lake; 17文笔海 Wenbi Lake;18拉市海 Lashi Lake;19 泸沽湖 Lugu Lake;20 纳帕海 Napa Lake;21 大山包 Dashanbao Lake。
1.2 调查与取样方法
在每个湖泊湿地样区周边各选择2、3个乡镇及多个村庄为调查点,采用灯诱捕法(使用改装后的功夫小帅牌(220v/50Hz,24w)光催化捕蚊器-武汉吉星环保科技有限责任公司生产)在湖畔临水域畜圈、禽舍、鱼棚内夜间吸血活动的成蚊进行通宵取样(晚19:30至次日晨7:30),每湖泊连续布灯诱捕3~4晚,每晚布灯4(5)个。所获标本低温冷冻致死后,应用40倍普通体解剖镜进行蚊种的分类鉴定,计数和记录。同期收集当地相关环境因素(如海拔高度、气候环境、纬度、平均气温和降水量等)等信息。
1.3 数据处理及分析方法
以各湖泊湿地(或纬度和海拔梯度带)内出现的物种数表示物种丰富度; 用α多样性指数(Shannon-Wiener指数和Pielou指数)测度和比较局域(湖泊、纬度、海拔梯度)的物种多样性; 用γ多样性描述和比较物种丰富度沿环境梯度(纬度、海拔带)的变化规律(方精云等,2004);用Sorenson相似性指数表示不同环境梯度(纬度、海拔梯度带)间物种组成的相似程度。
(1)物种丰富度(Species richness):S=各纬度(海拔)梯度带的物种数
(2)物种相似性系数
Sorenson指数:SI=2c/(a+b)
(3)α多样性指数
Shannon-Wiener指数:
Pielou指数:E=H′/lnS
其中,S为各个样地(湖泊、或环境梯度带)的物种数,a和b分别为各湖泊或环境梯度带内各自的物种数,c为两湖泊或环境梯度带的共有物种数,g(H)为沿两湖泊或环境梯度带增加的物种数,l(H)为沿两湖泊或环境梯度带减少的物种数,Pi为调查群落(湖泊)中各梯度带内第i个种的个体比例。
(4)用生态优势度分析群落或样区优势种的集中程度,优势度分析采用Simpson(1949)的公式(赵志模和郭依泉,1990):
式中,C=生态优势度,ni=每一种个体数,N=总个体。
(5)蚊类密度(Density):密度(D)=只/灯·晚。
(6)物种多度(Abundance): 物种多度(A)=ni/N。
(7)数据分析:聚类分析相关分析采用SPSS16.0完成。
2 结果
2.1 云南高原湖泊湿地蚊类的组成及分布
通过21个湖泊湿地的调查,共捕获蚊类506 974只, 经鉴定隶属于2亚科6属34种。其中,按蚊属Anopheles7种、库蚊属Culex18种、伊蚊属Aedes6种、阿蚊属Armigeres、曼蚊属Mansonia和脉毛蚊属Culiseta各1种。各湖泊蚊类的组成情况详见表1。
在所获34种蚊类中, 三带喙库蚊Cx.tritaeniorhynchus, 致倦库蚊Cx.pipiensquinguefasciatus和希氏库蚊Cx.theileri在各个湖泊均有分布,分布广泛;中华按蚊An.sinensis在20个、环带库蚊Cx.annulus和骚扰阿蚊Ar.subalbatus在16个湖泊有分布,分布亦较广;而其他蚊种分布范围相对较窄。
2.2 云南省高原湖泊湿地蚊类物种及其多度的空间分布特征
从表 2中看出,在所获34种蚊类种,不同蚊种对环境和气候条件的适应性和分布状况显然不同。中华按蚊、致倦库蚊、希氏库蚊、三带喙库蚊、刺扰伊蚊Ae.vexans和骚扰阿蚊在调查区域内北纬23°~28°和海拔1 000~3 500 m之间的各梯度带均有分布,显示了它们具有较强的气候和环境适应和分布的能力。此外, 昆明按蚊An.kunmingensis、常型曼蚊Ma.uniformis和环带库蚊在上述大多数纬度和海拔梯度带有分布,也反映较强的分布能力,但比较而言,昆明按蚊主要分布于纬度和海拔相对较高的区域,而后两者的分布区域则相反。余下的其他蚊种分布能力相对较窄或很窄。
数据统计结果显示,所获蚊类物种之间的相对多度(构成比)明显不同,其中:三带喙库蚊数量占明显优势(86.26%), 可以认为它是高原湖泊湿地蚊类数量上的优势种;中华按蚊和致倦库蚊分别占捕获蚊类总数的9.36%和2.81%, 数量上较为常见;其他23种蚊类的数量均少于捕获总数的1%, 在本区域的数量相对较少。
2.3 云南高原湖泊湿地蚊类γ多样性(物种丰富度)的空间分布格局
从图2和表 3中看出,蚊类物种丰富度随着纬度的升高, 呈双峰分布格局,两高峰的具体位置处于Ⅱ带(24°N)和Ⅳ带(26°N), 蚊类物种数分别达17和25种。比较而言,在这两个梯度带内,所分布的湖泊数量均相对较多(6和7个),反之,位于Ⅲ带(北纬25°)的湖泊仅有2个(表2),物种丰富度也最底。它反映出蚊类物种丰富度的高低,与梯度带所分布湖泊湿的数量多少有一定关系。
从图2和表4也可看出,蚊类的物种丰富度随海拔的升高其丰富度呈现先增高然后缓慢降低又下降近弧线型的宽峰分布格局, 峰值出现在B-D带(1 500~2 500 m), 发现蚊类在20~23种之间,显示了中海拔梯度蚊类物种丰富度相对较高的特点。 而随着海拔的升高,物种逐渐减少,在高海拔地区E带(3 000 m以上), 蚊种组成相对简单, 仅获13种。显示了云南高原湖泊湿地蚊类物种丰富度在海拔梯度的分布中,总体呈现了中间高两头低的变化趋势。

图2 云南高原湖泊湿地蚊类物种丰富度沿纬度和海拔梯度的变化Fig.2 Species richness of mosquitoes along a latitudinal and altitudinal gradient in the Lake Wetlands of Yunnan1:23°,1 000 m;2:24 °, 1 500 m;3:25°,2 000 m;4:26°,2 500 m;5:27°,3 000 m.
2.4 云南高原湖泊湿地蚊类α多样性的空间分布格局
从表3和图3中看出,在不同纬度梯度带,蚊类物种多样性、均匀度和优势度指数不尽相同,相对而言,在北纬26°和27°多样性指数(0.8391和2.0190))和均匀度指数(0.2607和0.7126)较高而优势度指数(0.6433和0.1715)较低,他们的密度也明显较低。它反映了云南高原湖泊湿地蚊类物种多样性与其物种的丰富度、均匀度、优势度与密度之间在不同纬度梯度上的变化状况及其相互关系。
根据表4、图3看出,在不同海拔梯度带,蚊类物种多样性、均匀度和优势度指数也不尽相同,相对而言,随着海拔的升高,蚊类物种多样性指数和均匀度指数总体呈现了逐步升高的趋势,而蚊类密度呈逐步下降的趋势,多样性指数(1.5740)和均匀度指数(0.6137)最高位于E带(3 000 m),而优势度指数(0.2402)和密度(29.44)是最低的。
上述蚊类物种多样性(α多样性)的空间分布格局,整体上呈现了沿着纬度和海拔的提升成逐步升高,而蚊类密度和物种优势度总体呈下降的趋势和特征。
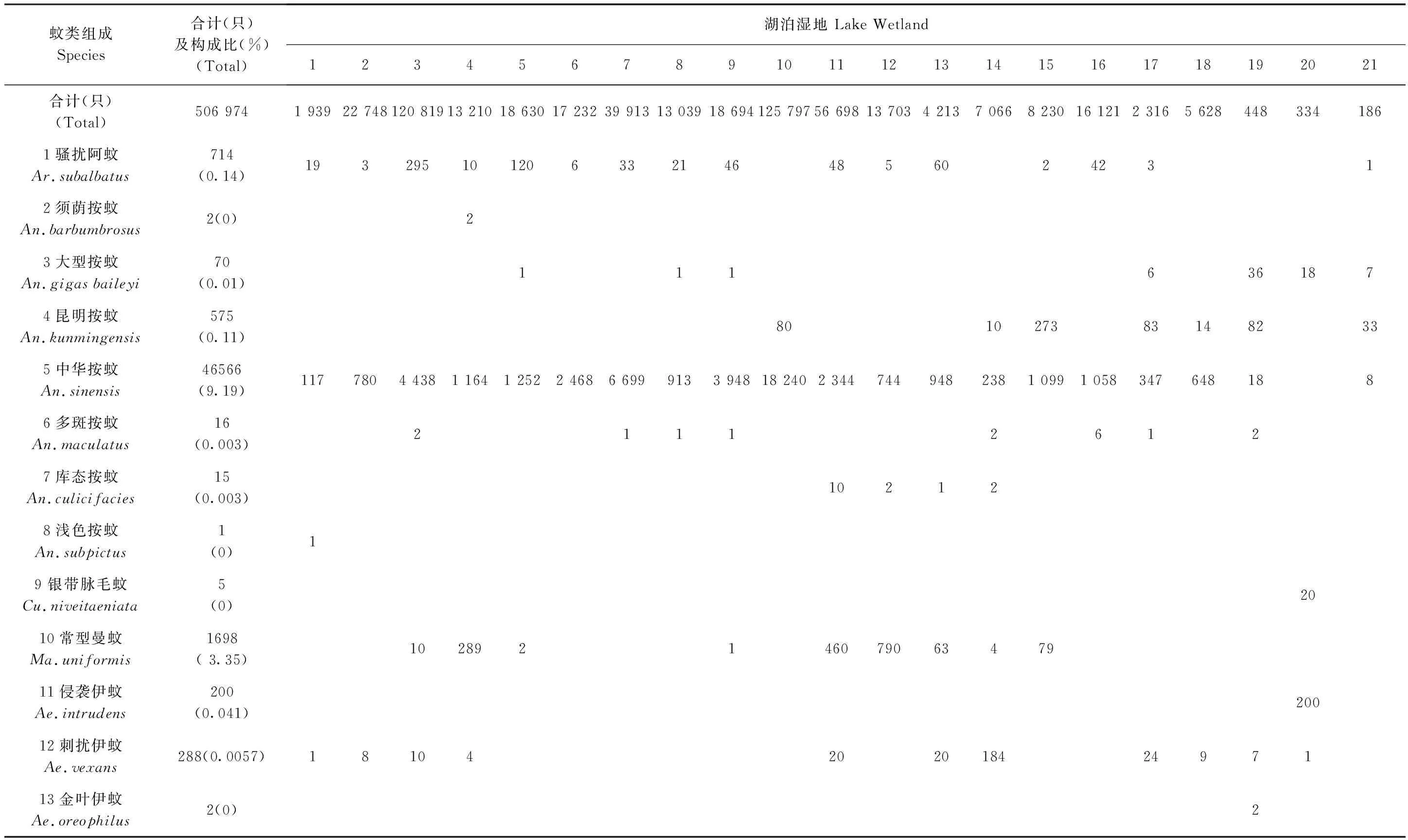
表1 云南省21个高原湖泊湿地蚊类的组成与分布(由南向北)Tab.1 Mosqutioes composition and distribution in the 21 Lake Wetlands of Yunnan Plateau
续表1
Tab.lContinued

蚊类组成Species合计(只)及构成比(%) (Total)湖泊湿地 Lake Wetland12345678910111213141516171819202114白蚊伊蚊Ae.albopictus1(0)115云南伊蚊Ae.yunnanensis4(0)416北部伊蚊Ae.tonkinensis4(0)1617环带库蚊Cx.annulus752(0.149)426529103818164162503745624083718二带喙库蚊Cx.bitaeniorhynchus40(0.008)171212132219棕头库蚊 Cx.fuscocephala2(0)220致倦库蚊Cx.pipiens quinquefascietus14 244(2.81)28641881 4481 5841 2021 9462 8142055351 166752130862751 43221665 993621拟态库蚊Cx.mimeticus3(0)1222伪杂鳞库蚊Cx.pseudovishnui87(0.018)21231147212723中华库蚊Cx. Sinensis3(0)2124希氏库蚊Cx. theileri3252(0.64)42813570219215377311258142118796116102341006767325三带喙库蚊 Cx.tritaeniorhynchus437 308(86.26)1768218631158161027215534134743097790521411410644352198112032828568161671351413784754132459526薛氏库蚊Cx.shebbearei36(0.007)213327褐尾库蚊Cx.fuscanus18(0.004)125622
续表1
Tab.lContinued

蚊类组成Species合计(只)及构成比(%) (Total)湖泊湿地 Lake Wetland12345678910111213141516171819202128贪食库蚊Cx.halifaxia5(0)112129棕盾库蚊Cx.jacksoni3(0)11130黄氏库蚊Cx.huangae138(0.029)206104831天坪库蚊Cx.tianpingensis4(0)1332小拟态库蚊 Cx.mimulus4(0)533类拟态库蚊Cx.murrelli6(0)221134角管库蚊 Cx.hutchinsoni2(0)11物种丰富度(S)34871212977119911121314121314101298
1:大屯海Datun Lake;2:长桥海Changqiao Lake;3:异龙湖Yilong Lake;4:普者黑Puzhehei Lake;5:杞麓湖Gilu Lake;6:星云湖Xingyun Lake;7:抚仙湖Fuxian Lake;8:滇池Dianchi Lake;9:阳宗海Yangzong Lake;10:青海湖Qinghai Lake;11:洱海Erhai Lake;12西湖Xihu Lake;13:茈碧湖Cibi Lake;14: 剑湖Jianhu Lake;15: 草海Caohai Lake;16:程海Chenghai Lake;17:文笔海Wenbi Lake;18:拉市海Lashi Lake;19: 泸沽湖Lugu Lake;20: 纳帕海Napa Lake;21: 大山包Dashanbao Lake.

表2 云南省高原湖泊湿地蚊类组成及其物种多度的空间分布Tab.2 Spatial distribution and abundance of mosquitoes species in the 21 Lake Wetlands of Yunnan Plateau
续表2Tab.2Continued

蚊类组成(Species)合计(只、%)(Total、%)纬度梯度(N) Latitudinal zone海拔梯度(M) Altitudinal zone23°-24°-25°-26°-27°-1 000-1 500-2 000-2500-3 000-24希氏库蚊Cx. theileri3252(0.64)610245691 516137141 5951 2902837025三带喙库蚊Cx.tritaeniorhynchus437308(86.26)139 44793 423158 64145 525272149 719255 30621 09011 05314026薛氏库蚊Cx.shebbearei36(0.007)234213327褐尾库蚊 Cx.fuscanus18(0.00)351037828贪食库蚊Cx.halifaxia5(0.00)2311329棕盾库蚊Cx.jacksoni3(0.00)1211130黄氏库蚊Cx.huangae138(0.03)2011828610431天坪库蚊Cx.tianpingensis4(0.00)131332小拟态库蚊Cx.mimulus5(0.00)5533类拟态库蚊Cx.murrelli6(0.00)2224234角管库蚊Cx.hutchinsoni2(0.00)211合计(只)(Total)506 974145 507120 717182 49557 277978158 716306 12427 29814 306530密度(D)6 924.141 886.2010 735.001 004.8633.725 119.873 779.31827.21572.2429.44物种丰富度(S)3514171425161723212013

表3 云南高原湖泊湿地蚊类物种多样性沿纬度梯度的变化Tab.3 Mosquitoes diversity along latitudinal gradients in the 21 Lake Wetlands of Yunnan

表4 云南高原湖泊湿地蚊类物种多样性沿海拔梯度的变化Tab.4 Mosquito diversity along altitudinal gradients in the 21 Lake Wetlands of Yunnan
2.5 云南高原湖泊湿地蚊类物种多样性、密度和优势种多度的空间分布及其关系
从图4中看出,高原湖泊湿地蚊类多物种样性与总蚊密度和三带喙库蚊多度两者沿纬度梯度的空间分布格局明显呈负相关关系,而总蚊密度和三带喙库蚊多度两者间整体呈现了正相关系的变化趋势(表6)。

图3 云南高原湖泊湿地蚊类α多样性随纬度和海拔梯度的变化Fig.3 Mosquito α diversity index along latitudinal and altitudinal gradients in the 21 Lake Wetlands of Yunnan1: 23°,1 000 m;2: 24°,1 500 m;3: 25°,2 000 m;4: 26°,2 500 m;5: 27°,3 000 m.

图4 云南高原湖泊湿地蚊类物种多样性和密度随纬度梯度的变化Fig.4 Mosquito density and α diversity index along latitudinal gradients in the 21 Lake Wetlands of YunnanⅠ:23°-;Ⅱ:24 °-;Ⅲ:25°-;Ⅳ:26°-;Ⅴ:27°-.

图5 云南高原湖泊湿地蚊类物种多样性、密度随海拔梯度的变化Fig.5 Mosquito density and α diversity index a altitudinal gradients in the 21 Lake Wetlands of YunnanA:1 000 m-; B:1 500 m-; C:2 000 m-; D:2 500 m-; E:3 000 m-.
从图5中看出,高原湖泊湿地蚊类物种多样性随着海拔的升高基本呈升高的趋势,而蚊类密度呈现了明显下降的趋势,两者之间的空间分布格局明显呈负相关关系。此外,它们与三带喙库蚊多度分布格局也有一定的关系(表6)。
上述结果总体上显示了湖泊湿地蚊类物种多样性、密度和优势种三带喙库蚊多度之间,沿纬度和海拔梯度上的协同变化趋势及其关系。
2.6 云南高原湖泊湿地蚊类空间分布的聚类分析与区系分异
根据不同纬度梯度带蚊类组成的聚类结果(图6),看出云南高原湖泊湿地蚊类可划分为3个主要地域或区系类型。 第一类仅含Ⅰ-Ⅲ带(北纬23°~25°N), 该梯度带由于地处热带气候边缘, 属于东洋区系;第二类为Ⅳ带(北纬26°N), 地处北亚热带至暖温带区间,属于东洋区系与古北区系的过渡地带,特点是两区系物种分布混杂。第三类为Ⅴ带(北纬27°N),该区域地处青藏高原的南缘, 基本属于寒温带气候,这里基本属于古北区系, 代表种有银带脉毛蚊Cu.niveitaeniata等。

图6 云南21个高原湖泊湿地蚊类沿纬度梯度分布的聚类分析Fig.6 Cluster analysis of mosquito species along latitudinal gradients in the 21 Lake Wetlands of YunnanⅠ:23°-; Ⅱ:24 °-; Ⅲ:25°-; Ⅳ:26°-;Ⅴ: 27°-.
根据不同海拔梯度带组成的聚类结果(图7)可以看出,云南高原湖泊湿地蚊类也可划分为3个主要区域或区系类型。 第1类仅含A带, 海拔1 000~1 400 m之间,该梯度带由于地处南亚热带气候,应属于东洋区系; 第2类包含B-D带, 海拔1 500~2 500 m之间,这里地处北亚热带至暖温带区间, 属于东洋区系与古北区系的过渡区, 特点是两区系物种分布混杂; 第3类为E带,海拔2 500~3 500 m之间,地处青藏高原的南缘,基本属寒温带气候, 基本属于古北区系,代表种可见有银带脉毛蚊和侵袭伊蚊Ae.intrudens等。
1:大屯海Datun Lake;2:长桥海Changqiao Lake;3:异龙湖Yilong Lake;4:普者黑Puzhehei Lake;5:杞麓湖Gilu Lake;6:星云湖Xingyun Lake;7:抚仙湖Fuxian Lake;8:滇池Dianchi Lake;9:阳宗海Yangzong Lake;10:青海湖Qinghai Lake;11:洱海Erhai Lake;12西湖Xihu Lake;13:茈碧湖Cibi Lake;14: 剑湖Jianhu Lake;15: 草海Caohai Lake;16:程海Chenghai Lake;17:文笔海Wenbi Lake;18:拉市海Lashi Lake;19: 泸沽湖Lugu Lake;20: 纳帕海Napa Lake;21: 大山包Dashanbao Lake.

图7 云南21个高原湖泊湿地蚊类沿海拔梯度分布的聚类分析Fig.7 Cluster analysis of mosquito species along altitudinal gradients in the 21 Lake Wetlands of YunnanA:1 000 m-; B:1 500 m-; C:2 000 m-;D:2 500 m-;E:3 000m-.
上述不同垂直梯度带蚊类组成相似性的聚类结果呈现了与纬度梯度近似的特征。因此认为:在云南北纬26°N ~27°N 带和海拔2 000~2 500 m之间的区间,应该是云南蚊类区系沿纬度或海拔梯度发生明显改变和区系分异的区域。
2.7 云南高原湖泊湿地蚊类物种多样性与地理和气候环境因素的关系
2.7.1蚊类物种多样性与地理位置关系 从表5中可以看出,不同地理位置的21个高原湖泊湿地蚊类物种多样性显然各不相同,物种多样性和均匀度最高者是地处滇西北海拔和纬度较高、气候温凉、人口稀少、森林植被和环境保护较好,属于自然保护区泸沽湖(1.8281和0.7127)、纳帕海(1.4427和0.6566)和大山包(1.4210和0.6834),而物种多样性和均匀度最低的是位于南部海拔和纬度较低,人口密集,近邻城市、开发和污染较大的长桥海(0.1832和0.0941)、大屯海(0.3842和0.1818)、异龙湖(0.1923和0.0770)等湖泊。
2.7.2蚊类物种多样性与重要环境因素的关系

表6 云南21个高原湖泊湿地蚊类物种多样性与重要环境因素和主要蚊种多度的关系分析Tab.6 Mosquito abundance and species diversity related to the environmental factors in the 21 Lakes Wetlands of Yunnan
*P< 0.05;**P< 0.01.
分析: 通过21个高原湖泊湿地蚊类物种多样性(物种丰富度、多样性指数、均匀度指数、优势度指数、优势种多度)与重要地理环境因素之间关系的分析,结果发现:(1)物种丰富度与海拔(r=0.124)、降水量(r=-0.034)和气温(r=0.122)之间的关系均不显著;(2)物种多样性指数和均匀度指数与海拔高度(r=0.760)和r=0.790)呈显著正相关,与气温(r=-0.710 和r=-0.794)呈显著负相关,与降水量(r=-0.059 和r=-0.015)无相关,但与三带喙库蚊种多度(r=-0.832和r=-0.851)和物种优势度(r=-0.552)均呈显著负相关。(3)物种优势度指数与海拔呈负相关(r=-0.446),与气温(r=0.226)和降水量(r=0.366 )关系不显著,但与三带喙库蚊种多度(r=0.712)呈显著正相关。
3 讨论
3.1 湖泊湿地蚊类物种多样性的区域特征
湖泊湿地是我国的重要湿地生态系统之一,具有许多经济价值和社会服务功能。既往,全国所记录的湖泊湿地蚊类仅有2亚科4属14种,其中云南湖泊湿地分布2属5种(臧颖慧等, 2009b)。本研究采用通宵灯诱捕法对云南21个天然高原湖泊湿地进行调查后,共发现蚊类2亚科6属34种,丰富和填补了我国西南高原湖泊湿地蚊类物种多样性空间分布特征等研究的空白,同时也反映了当地高原湖泊湿地蚊类的区系组成较为复杂和多样性较高的区域特征。
在所获34种蚊类中,以库蚊属种类最多(18种),其次是按蚊属(7种)和伊蚊属(6种),其他种类较少。比较而言,所获蚊种主要以夜间吸血活动习性为主的库蚊和按蚊属的偏多,而其他在白昼吸血活动为主的伊蚊、阿蚊属等的种类或数量均相对较少。说明应用灯诱法进行通宵捕蚊的调查结果可能存在一定的局限性。如果在通宵灯诱的基础上,并针对以白天吸血活动为主的成蚊和稀有蚊类的幼虫标本进行辅助性采集,将会对湖泊湿地蚊类物种多样性的了解更加全面。
3.2 蚊类空间分布的聚类分析与区系特征
蚊类区系的划界是我国蚊虫自然地理学的重要内容, 中国蚊类区系已往的划界主要是依据某些蚊属、种团和代表种, 如按蚊属和尖音库蚊复合组Culexpipienscomplex的界间替代分布来确定的, 至今其结论和意见仍不统一(陆宝麟,1997)。由于我国西南横断山区特殊的地域特征, 形成当地生物两大区系及其分布形成交叉重叠的特有现象, 致使该地域的区系划界问题存在众多的争议。本研究将云南省北纬21°08′~ 29°01′;东经97°31′~ 106°11′和海拔500~3 500 m之间的区域作为研究的空间范围,以各纬度带或海拔带蚊类组成的异同, 应用系统聚类分析的方法将其划分为东洋区系、两区系交汇或过渡区和古北区系3个主要地域区系类型。可以认为:在云南北纬26°N ~27°N 带和海拔2 000~2 500 m之间的区间,应该是云南蚊类区系沿纬度或海拔梯度发生明显改变和区系分异的区域。 此结果与 “三江并流”的澜沧江、怒江流域的蚊类(葛军旗等,2008;张菊仙等,2008)以及横断山区小型兽类(权寿瑛等,2009)及其寄生蚤类区系水平和垂直分布格局与特征(龚正达等,2005;2007)的研究结果基本一致, 它们共同反映了我国西南部横断山区动物、昆虫区系空间分布格局的一般规律和区域特征。
3.3 蚊类多样性空间分布格局与重要环境因素的关系
云南地势南低北高, 气温和雨量由南到北或由低到高按一定的规律变化, 形成完整的气候和植被带谱(热带、亚热带、暖温带和寒温带等)。因其所处的地貌和海拔高度不同,不同地理位置、气候带湖泊的蚊类组成和分布存在明显的变化, 反映出蚊类多样性空间分布格局与地理环境和气候条件之间的变化与联系。总体而言,在纬度和海拔较低、气候较热、人口密集、湖边大量种植水稻或莲藕、污染较重和人为干扰较大的区域蚊类密度较高,而物种多样性和均匀度指数都相对较底,反之则高。说明人为干扰因素对云南高原湖泊湿地蚊类多样性的地理分布变化趋势具有重要影响。
优势种三带喙库蚊多度与物种多样性指数、均匀度指数和海拔高度呈显著负相关;而与密度和物种优势度指数、气温显著正相关。三带喙库蚊主要分布于海拔较低和气候偏热的区域,它不仅分布广泛且是数量上优势种,随着当地地理、气候环境和条件的变化与差异,其物种多度的空间分布特征主导了云南高原湖泊湿地蚊类密度、优势度和物种多样性的地理变化趋势。
在自然环境中的不同条件下,蚊类的孳生都必须依赖各种类型天然或人为形成的水体。蚊类物种多样性指数与均匀度、生态优势度、海拔和年平均气温呈相关关系,而与降水量关系较小,说明蚊类物种多样性与湖泊湿地特有的水湿条件等重要环境因素关系密切。 湖泊湿地是介于水域生态系统和陆地生态系统之间的一种特殊而稳定的生态系统,尤其是非季节性(永久性)的天然湖泊湿地湖畔周边的常年积水和过湿土地可为蚊类幼虫常年孳生提供稳定的水湿环境和生长条件。因此,湿地蚊类的孳生和生长完全可以不依赖当地雨量多少而存在。湖泊湿地蚊类物种多样性与年降水量无关,说明永久性的天然湖泊湿地这类非常稳定和特殊的生态系统对当地蚊类物种的保存、滋生、繁衍和物种扩散都可能起到了非常重要的作用。
此外,湖泊湿地蚊类物种多样性与当地降雨量无关,也形成了它与云南西部横断山区蚤类(龚正达等,2001)和 “三江并流”区域的澜沧江、怒江和金沙江流域的蚊类(葛军旗等,2008;张菊仙等,2008;李栋等,2011)不同地理环境、区域类型和生态系统中,物种多样性空间分布格局与雨量的多少、分配特征密切相关而显然不同的生态学特征。
3.4 蚊类与虫媒疾病的关系
云南由于其特殊的地理位置和复杂的自然气候,研究证实存在着乙型脑炎、登革热、基孔肯雅、辛得比斯、巴泰、可提、致病性环状病毒等多种蚊媒病毒(边长玲等,2009),新发现浓核病毒和kadipiro病毒等虫媒病毒(Sunetal.,2009)。云南不仅为我国蚊类区系和物种分布和分化的中心(张菊仙等,2008),也是我国目前蚊传虫媒病毒检出和发现种类最多的地区(边长玲等,2009)。
近年来, 媒介生物群落多样性在疾病传播中所起的作用正在被人们所认识, 并受到越来越多的关注(Ostfeldetal, 2000; Ezenwaetal.,2006;Mills, 2006)。媒介生物群落的高多样性能降低某些人畜共患传染病(zoonoses)的流行风险。本次调查发现, 云南湿地蚊类在北纬25°以南和海拔2 000 m以下区域的均匀度和多样性指数都较低,而蚊类的密度和优势种三带喙库蚊多度均较高。而这些区域正好多是当地乙型脑炎等虫媒疾病流行和频发的区域(边长玲等,2009)。
云南高原湖泊湿地蚊类的优势种三带喙库蚊(86.26%)、常见种中华按蚊(9.36%)和致倦库蚊(2.81%) 它们不仅数量多,且分布较为广泛。目前,据国内研究结果统计:三带喙库蚊和中华按蚊分别能自然感染8种虫媒病病原,而致倦库蚊能自然感染6种虫媒疾病病原(边长玲等,2009),三者均为国内自然感染率高且传播病原种类最多的蚊种。此外,有学者认为随着候鸟迁飞过境和在湖泊停栖, 输入西尼罗病毒而引发西尼罗病毒病可能性很大(瞿逢伊,2006)。因此,天然湖泊湿地这类稳定和特殊的生态系统对上述媒介蚊种常年的孳生、繁衍以及各类病原的长期保存、扩散和疾病流行都会起到非常重要的作用或存在一定的关系。
云南地形和气候环境复杂多样,在我国5大类湿地中,除缺乏滨海湿地之外,沼泽湿地、湖泊湿地、河流湿地和人工湿地4大类湿地在不同地理和气候环境内均广泛分布。除本研究涉及的高原湖泊湿地外,还有高原河流湿地、高原溪沟湿地、高原沼泽湿地、高原水库湿地、高原塘池湿地和高原水田湿地等(金振洲,2009)。鉴于此, 云南不同类型湿地蚊类群落和物种多样性空间分布特征、重要媒介物种多度分布与气候环境、干扰因素和疾病之间的关系,以及蚊媒疾病的防控等内容都是值得进一步研究和探讨的问题。
致谢现场调查工作中得到迪庆、丽江、玉龙、宁蒗、永胜、剑川、鹤庆、祥云、玉溪、通海、澄江、昭通、红河、蒙自、石屏、昆明、宜良、晋宁、文山、丘北等地州市县疾控中心和乡镇卫生院的支持和帮助,谨此致谢!
猜你喜欢
热带作物学报(2022年9期)2022-10-17
大自然探索(2021年9期)2021-11-07
中央民族大学学报(自然科学版)(2018年4期)2018-03-19
齐鲁周刊(2017年29期)2017-08-08
环境昆虫学报(2016年4期)2016-08-23
动物医学进展(2015年11期)2015-06-11
植物营养与肥料学报(2014年1期)2014-03-11
测绘学报(2014年2期)2014-01-11
寄生虫与医学昆虫学报(2013年4期)2013-11-19
中国烟草学报(2012年1期)2012-04-09
