
豇豆产量性状与SSR分子标记的关联分析
2015-10-28 22:14潘磊等
湖北农业科学 2015年16期
潘磊等

摘要:利用156对多态性豇豆[Vigna unguiculata(L.)Walp.]简单重复序列(Simple sequence repeat,SSR)引物检测83份豇豆种质材料基因组,分析群体遗传结构,并对14个豇豆产量性状与SSR标记进行全基因组关联分析。结果表明,群体遗传结构分析将83份豇豆样品划分为2个亚群。关联分析则有10个SSR标记位点与8个性状关联,主要分布在LG2、LG3、LG4、LG7、LG11连锁群上。这些关联位点在豇豆基因组上分布不均,对关联性状的表型变异解释率为9% (CLM0022)~33%(CLM0347);有1个标记(CLM0251)与多个性状关联;另外也有同一个性状与多个SSR标记关联,包括叶宽(CLM0251、CLM0850),单荚重(CLM0347、CLM0251、CLM0614)。这些与豇豆性状关联的SSR标记将为豇豆分子育种和遗传改良提供依据。
关键词:豇豆[Vigna unguiculata(L.)Walp.];产量性状;简单重复序列;关联分析
中图分类号:S643.4;Q343.1+5;Q348 文献标识码:A 文章编号:0439-8114(2015)16-3952-07
DOI:10.14088/j.cnki.issn0439-8114.2015.16.031
Association Analysis between Yield Trait in Cowpea and SSR Molecular Markers
PAN Lei,LI Yi,YU Xiao-lu,GUO Rui,CHEN Chan-you
(Hubei Province Engineering Research Center of Legume Plants/School of Life Sciences,Jianghan University,Wuhan 430056,China)
Abstract: Correlation of yield traits and molecular markers in cowpea [Vigna unguiculata(L.)Walp.] were investigated by using mixed line model in genome-wide association study. A set of 156 polymorphic Simple sequence repeat (SSR) markers were applied to genotyped 83 typical cowpea accessions. Based on the polymorphic data, population genetic structure analysis revealed two sub-populations revealed in the experimental 83 samples. After association analysis was performed between 14 yield traits in cowpea and the 156 polymorphic SSR molecular markers,ten SSR were found to be significantly associated with eight yield traits. These markers distributed in different linkage groups,including LG2,LG3,LG4,LG7,LG11, which explained 9%(CLM0022)~33% (CLM0347) of the observed phenotypic variance.There were one SSR markers (CLM0251) associated with two or more traits. On the other side, there two yield traits correlated with two or more SSR markers,including leaf width (CLM0251 and CLM0850) and single pod weight (CLM0347,CLM0251 and CLM0614).These SSR marker related with cowpea traits will be useful for molecular breeding and genetic improvement in cowpea in the future.
Key words: cowpea[Vigna unguiculata(L.)Walp.]; yield trait; Simple sequence repeat; association analysis
豇豆[Vigna unguiculata(L.) Walp.]是一种重要的豆科(Leguminosae)蔬菜作物之一,其嫩荚和种子可供食用,是日常饮食中植物蛋白质和膳食纤维的重要来源之一。中国的豇豆栽培历史悠久,资源类型多样,遗传变异丰富。目前,中国农业科学院国家农作物种质资源保存中心(http://icgr.caas.net.cn/)已搜集保存了1 000份以上的豇豆种质资源;而这些豇豆资源中,生长习性以蔓生、半蔓生和矮生类型为主,花色多为紫色或白色,鲜荚颜色有深绿、浅绿、白色、红色、紫红色、龙纹等,诸多优异农艺性状可用作遗传改良资源。探究遗传变异的基础,挖掘优异等位基因或性状连锁的遗传位点,将是豇豆种质资源与分子育种中的重要研究方向之一,关联分析(Association analysis)为此提供了一个十分有效的研究方法,关联分析是近年发展起来的一种高通量分析表型与遗传多态性之间关联性的遗传分析方法,能在群体水平上大量发掘与目标性状相关的基因及其等位基因。2001年关联分析首次用于植物研究[1],之后逐渐成为植物数量性状遗传研究的热点之一。关联分析又称为关联作图(Association mapping),或连锁不平衡作图(Linkage disequilibrium mapping,LD mapping),是以基因或者位点之间的连锁不平衡为基础,从而筛选鉴定出某一群体内的目标性状与遗传标记或候选基因,主要有基于候选基因的关联分析和全基因组关联分析两种策略[2]。关联分析中多采用混合线性模型(Mixed liner model,MLM),因为此模型兼顾群体结构和系谱关系,能显著降低误差[3]。目前尚未见到豇豆研究应用关联分析的报道,但是豇豆资源的亚群中存在一定的遗传结构和较高的连锁不平衡(LD)水平[4]。理论上位点之间连锁越紧密,LD水平则越高,而LD较高的物种或群体应用较少的标记即可实现全基因组扫描。此外,自花授粉的物种,在经历了瓶颈效应和强烈的人工选择后,群体中仅包含所有群体中少部分的等位基因,这更适宜于全基因组关联分析的应用[5]。豇豆是典型的自花授粉植物,基因组较小(约587 Mb)[6],群体内的LD水平较高。因此,全基因组扫描的关联分析可以应用在豇豆研究中。
多态性DNA分子标记是开展全基因组关联分析的前提。与RADP、AFLP、ISSR等分子标记相比较,简单重复序列(Simple sequence repeat,SSR)分子标记由于在基因组上分布广、经济方便、多态性高、重复性好、易于基因分型等优点,在植物中的应用备受青睐。在豇豆的DNA分子标记研究中,国内外的研究者陆续报道了运用多种DNA分子标记(RAPD、RFLP、AFLP和SSR等)技术构建了豇豆的连锁遗传图[7-12],这些遗传图谱的标记虽然数量少,但为豇豆基因组的进一步研究奠定了基础。
试验选取了全国范围内的83份具代表性的豇豆种质材料,对豇豆产量的相关性状,包括商品成熟荚相关的性状(鲜荚荚长、鲜荚荚厚、鲜荚单荚重、首荚节位和单株结荚数等)以及储藏期的干种子相关性状(千粒重、种子长和种子宽等)进行了有重复的表型鉴定,利用豇豆全基因组上均匀分布的175对SSR分子标记,进行产量性状与SSR标记的亚群聚类和关联分析。基于供试材料来源不同和SSR分子标记的多态性、一个关联位点的多个等位变异(多态性条带)能对应多少个表型性状等的特异性,来筛选出优异的等位变异,现将结果报告如下。
1 材料与方法
1.1 群体的选择
试验所用的83份豇豆种质材料均来自江汉大学湖北省豆类蔬菜植物工程技术研究中心,其性状表型稳定,基本情况见表1。分别在武汉市东西湖区柏泉农场和武汉市蔡甸区永安街柏木村,连续进行两年三季(2012年秋季、2013年春季和秋季)田间试验,观测记录豇豆产量的相关性状。所有样品的春播为4月,秋播为7月,田间管理为常规方法。
1.2 表型性状的测定与分析
试验测定了14个与豇豆产量相关的性状,包括商品成熟荚性状6个(鲜荚单荚长、叶宽、鲜荚荚厚、鲜荚单荚重、首荚节位、单株结荚数)和干种子形态性状8个(千粒重、种子面积、种子周长、种子长/宽、种子长、种子宽、种子直径、种子圆度),每个性状的测定有3次生物学重复。商品成熟荚性状的测定参照王佩芝等[13]的方法;干种子采用万深SC-E型种子外观品质检测分析仪系统(杭州万深检测科技有限公司)测量种子的形态性状。此外,采用电子天平称量干种子百粒重,然后换算为千粒重。将测定的性状汇总后,录入到Microsft Office Excel 2010软件里,计算各性状在品种内和品种间的变异系数。
1.3 SSR分析
1.3.1 基因组总DNA提取 选取豇豆嫩叶,用改进的CTAB法提取基因组总DNA[14],于-20 ℃环境下保存备用。采用0.8%琼脂糖凝胶电泳和紫外分光光度计测定DNA的质量和浓度。
1.3.2 引物筛选 挑选豇豆全基因组分布的175 对SSR引物[11],并委托上海生工生物工程技术服务有限公司合成引物,筛选具备多态性SSR引物,用于供试样品的扩增分析。
1.3.3 PCR扩增反应 PCR总体系为20 μL,其中包含20 ng DNA模板,引物10 pmol,4 nmol dNTPs,30 nmol MgCl2,0.5 U Taq DNA聚合酶(上海博彩生物科技有限公司),10×PCR Buffer,其余体积用ddH2O补齐。在Eppendorf Mastercycler gradient PCR扩增仪(德国艾本德中国有限公司)上进行PCR扩增反应,扩增条件:94 ℃预变性5 min; 94 ℃变性30 s,50~60 ℃退火30 s,72 ℃延伸 40 s,35个循环;最后72 ℃延伸5 min。以100 bp DNA Ladder作为参照,记录扩增片段的大小;此外,按照0/1二进制的方法,将有条带的记作1,无条带的记作0,录入到Microsft Office Excel 2010中形成数据矩阵。
1.4 数据分析
采用STRUCTURE 2.3软件(http://pritchardlab.stanford.edu/structure.html)分析供试豇豆群体的遗传结构,基于数学模型划分亚群,计算相应的Q值。主要分析步骤为:采用Admixed model模式,K值(亚群数目)设为1~12,参数Iterations设为10 000,Burn in-period设为100 000。每个K值重复运行5次,然后计算出每个K值对应的后验概率lnP(D)值。通过分析ΔK值的变化规律,来判别合适的K值作为亚群数目[15]。运用SPA Gedi1.2软件计算83份豇豆种质材料两两之间的亲缘系数,进行Kinship分析[16]。
SSR位点与豇豆产量性状的关联分析中,采用TASSEL 2.2软件的MLM模型分别进行多态性SSR位点与豇豆产量性状的关联分析[17]。主要步骤为:以Q值和Kinship系数矩阵作为协变量,综合156对多态性豇豆SSR引物获得的等位基因位点、亚群分类结果(依据K值)和表型数据,计算获得每个位点的显著性水平及其对表型变异的解释率。以P-Maker值来衡量关联标记的显著性,试验选择的显著水平为P<0.01。
2 结果与分析
2.1 表型性状的遗传变异分析
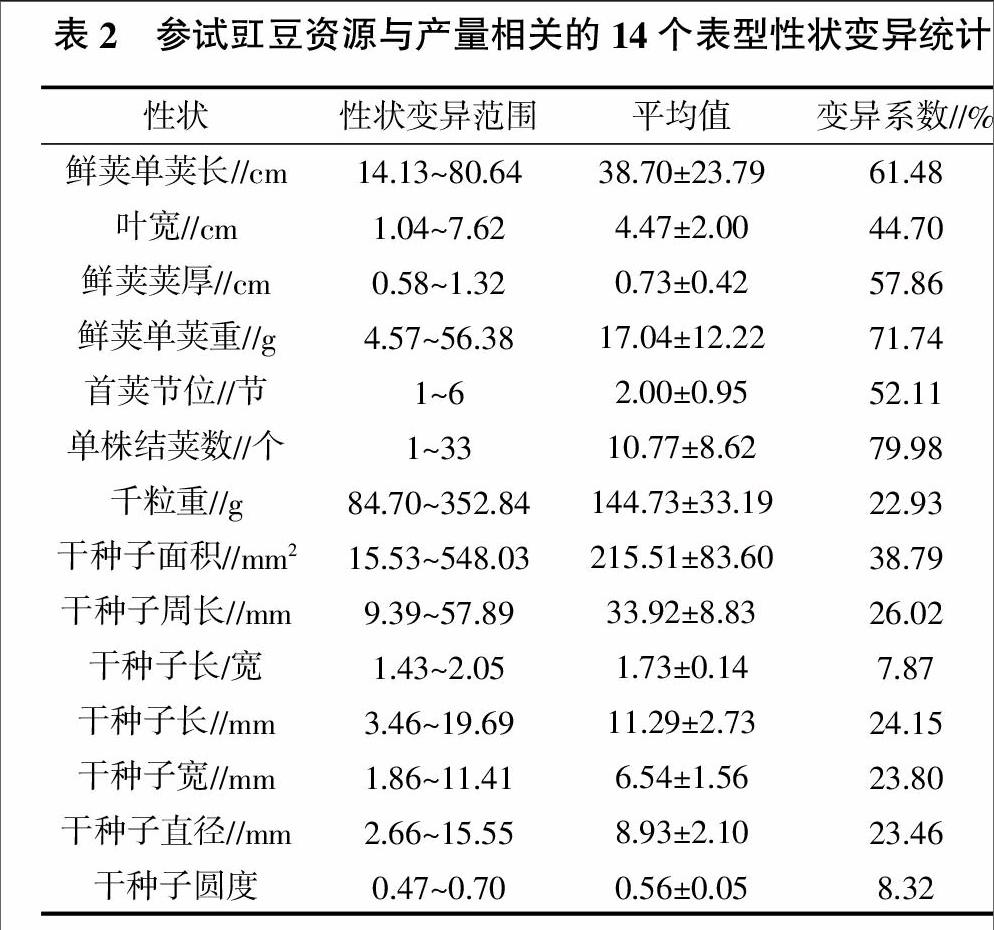
试验测定的豇豆鲜荚单荚长、叶宽、鲜荚荚厚、鲜荚单荚重、首荚节位、单株结荚数以及千粒重、种子面积、种子周长、种子长/宽、种子长、种子宽、种子直径、种子圆度14个与豇豆产量相关的表型性状变异统计结果见表2。从表2可见,供试的83份豇豆种质材料中14个产量相关性状存在明显的遗传变异;其中变异程度最大的性状为单株结荚数,其变异系数为79.98%,其次是鲜荚单荚重,其变异系数为71.74%,而干种子长/宽的变异程度最小,变异系数仅为7.87%,就各个性状而言,遗传变异的差别十分明显。在与鲜荚产量相关的6个性状中,鲜荚单荚长度的变异范围为14.13~80.64 cm,平均为38.70 cm;叶宽的变异范围为1.04~7.62 cm,平均在4.47 cm;鲜荚荚厚的变异范围在0.58~1.32 cm,平均在0.73 cm;鲜荚单荚重的变异范围在4.57~56.38 g,平均为17.04 g;首荚节位变异范围在1~6节,平均首荚节位为2节;单株结荚数变异范围在1~33个,平均结荚数约为11个。
在与干种子产量相关的8个性状中,千粒重变异范围在84.70~352.84 g,平均为144.73 g;干种子面积变异范围在15.53~548.03 mm2,平均在215.51 mm2;干种子周长变异范围在9.39~57.89 mm,平均为33.92 mm;干种子长/宽变异范围为1.43 ~2.05,平均为1.73;干种子长度变异范围在3.46~19.69 mm,平均为11.29 mm;干种子宽度变异范围在1.86~11.41 mm,平均为6.54 mm;干种子直径变异范围为2.66~15.55 mm,平均为8.93 mm;干种子圆度变异范围在0.47~0.70,平均为0.56。
2.2 遗传多样性分析
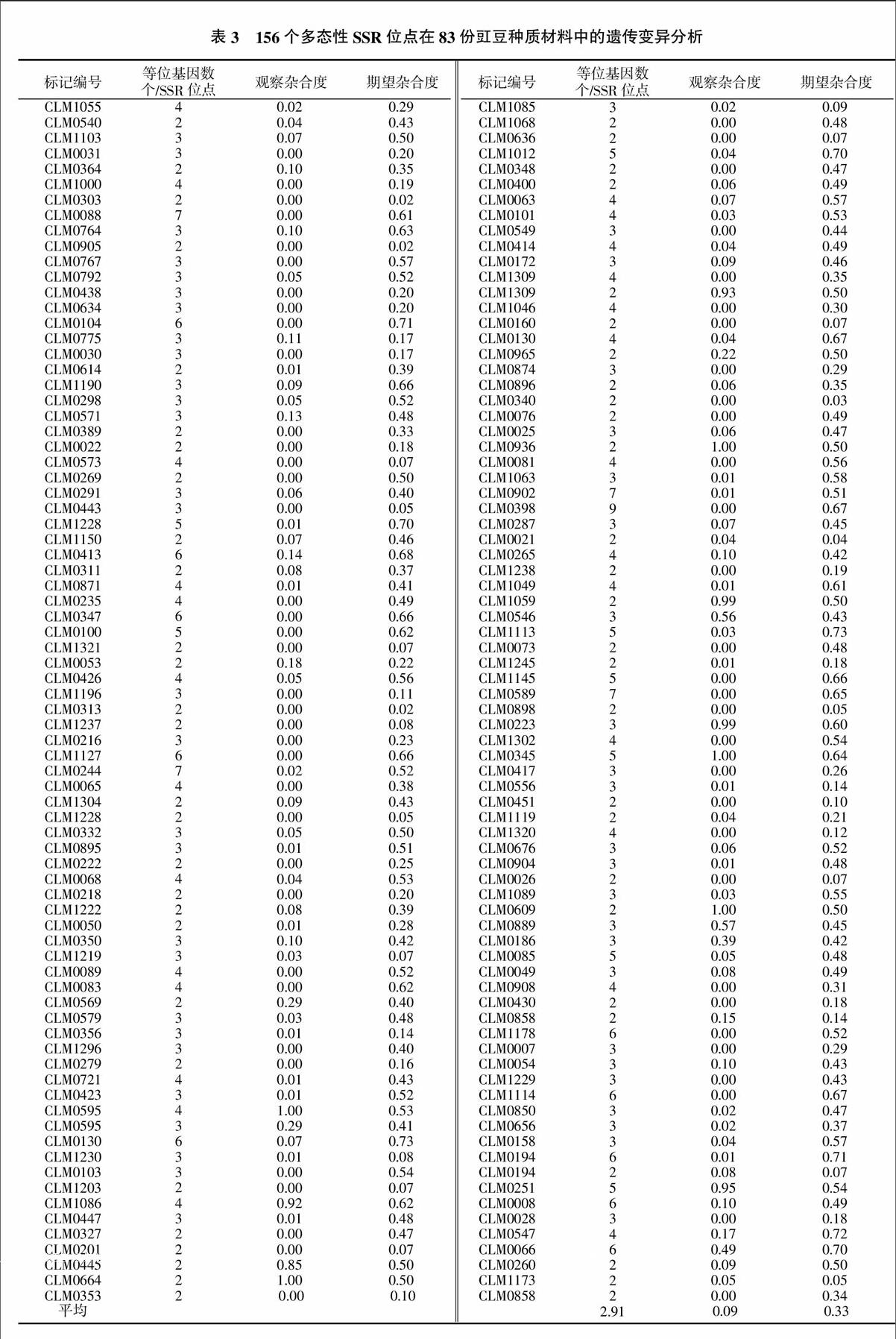
试验从175 对豇豆基因组SSR引物中筛选出了156 对多态性引物,并用于83份豇豆种质材料的扩增分析,结果见表3。检测结果表明,每对多态性引物中等位基因数为2~9个/SSR位点,平均为2.91个/SSR位点;观察杂合度的变异范围在0.00~1.00,平均观察杂合度为0.09;期望杂合度的变异范围在0.03~0.73,平均期望杂合度为0.33。据此可见,供试豇豆种质材料的遗传多样性水平为中等偏低。
2.3 群体遗传结构和亲缘关系聚类分析
83个豇豆种质材料基于多态性SSR位点和数学模型的群体遗传结构K值分析结果见图1。从图1可见,采用STRUCTURE 2.3软件建立的数学模型中,当K=2时,其模型的后验概率lnP(D)最大,即符合Hardy-Weinberg平衡的亚群数为2。据此,可将83个豇豆种质材料群体分为2个亚群,详情见图2。群体结构的存在会影响LD,从而造成关联分析结果的真实性和准确性,因此,将各个品种的Q值作为协变量,加入SSR分子标记与表型性状的关联分析中。
2.4 标记与性状的关联分析
豇豆产量性状与SSR分子标记位点的关联分析结果见表4,与产量性状显著关联的10个豇豆基因组SSR标记信息见表5,表4、表5中的连锁图数据引自文献[11]。从表4可见,10个SSR标记位点与8个豇豆产量性状相关联,与商品成熟荚产量性状(鲜荚单荚长、叶宽、鲜荚单荚重、首荚节位、单株结荚数)相关的位点有8个,分布于连锁群LG2(鲜荚单荚长,标记位点CLM0068;鲜荚单荚重,标记位点CLM0347)、LG3(鲜荚单荚重,标记位点CLM0614;单株结荚数,标记位点CLM1182)、LG4(叶宽,标记位点CLM0251;鲜荚单荚重,标记位点CLM0251)、LG7(首荚节位,标记位点CLM0445)、LG11(叶宽,标记位点CLM0850);与干种子形态(千粒重、种子宽、种子圆度)相关的位点3个,分布于LG3(千粒重,标记位点CLM0022)、LG10(种子宽,标记位点CLM0546;种子圆度,标记位点CLM0589)。
这些与表型性状相关联的SSR位点在豇豆基因组上的分布不均衡。与豇豆鲜荚单荚长度相关的位点1个,位于连锁群LG2;与叶宽相关联的位点有2个,分别位于连锁群LG4和LG11;与鲜荚单荚重相关联的位点有3个,分别位于连锁群LG2、LG3和LG4;与首荚节位相关联的位点1个,位于连锁群LG7;与单株结荚数相关联的位点1个,位于连锁群LG3;与千粒重相关联的位点1个,位于连锁群LG3;与种子宽相关联的位点1个,位于连锁群LG10;与种子圆度相关联的位点1个,位于连锁群LG10。这些标记对关联性状的表型变异解释率范围为9%(CLM0022)~33%(CLM0347),解释率越高说明关联程度越大;其中,以千粒重(9%)最低,鲜荚单荚重最高(33%)。
3 小结与讨论
近年来,在DNA分子水平上对豇豆的研究越来越受到关注[18]。尽管如此,与其他豆科农作物相比较,豇豆的分子水平研究基础与应用研究还十分滞后[19]。比如,豆科中的百脉根(Lotus corniculatus L. var. japonicus Regel)、大豆[Glycine max(L.)Merr.]、蒺藜苜蓿(Medicago truncatula Gaertn.)、木豆[Cajanus cajan(L.)Millsp.]、鹰嘴豆(Cicer arietinum L.)、菜豆(Phaseolus vulgaris L.)、绿豆[Vigna radiata(L.)Wilczek]和红小豆[V. angularis(Willd.)Ohwi et Ohashi]等均已完成全基因组测序,为豆科植物深入研究基因组结构、发掘功能基因和分子辅助育种等研究奠定了坚实的基础。豇豆中尚未开展基因组测序,因此,筛选豇豆基因组上多态性DNA分子标记技术、探究表型性状与分子标记的关联性、发掘与性状显著关联的分子标记、揭示关联位点的等位变异等,将为豇豆的分子育种和遗传改良提供分子水平依据。
试验从83个豇豆种质材料中共筛选出豇豆全基因组分布的156个多态性SSR标记,揭示了供试豇豆群体具有中等偏下的遗传多样性水平。而且对豇豆种质材料进行群体遗传结构分析后,发现其中存在一定的遗传分化,可将83个豇豆种质材料划分为2个亚群结构,推测可能是由于所选样品多为人工选育的后果,在长期的人工选择作用下,豇豆出现了遗传分化、各种质材料的遗传背景趋于狭窄、遗传多样性偏低造成的。因此,加大力度开展豇豆种质资源及其野生资源的收集,充实和丰富豇豆基因库资源,是今后豇豆种质资源与遗传育种研究所需要努力的重点方向。
豇豆的产量性状是一类复杂的数量性状,容易受到环境影响。因此试验选取不同地点进行田间种植,进行了连续两年三季的观测。分析发现,单株结荚数(变异系数79.98%)和鲜荚单荚重(变异系数71.74%)受环境影响相对比较大,而干种子长/宽(变异系数7.69%)则受环境影响相对较小。由此可见,在开展豇豆产量性状的分子辅助育种中,要考虑地区生态环境的差异对单株结荚数和鲜荚单荚重等产量性状的影响,借鉴当地环境下的豇豆产量性状相关基因或者QTL位点,对于育种而言是十分重要的。
目前,科技工作者已经发现了一批豇豆产量相关性状的QTL位点,包括豇豆子粒大小和鲜豆荚大小[20]、荚长[12]、始花期、始花节位、单株结荚数[21]、盛花时间[22]等相关QTL位点。但由于遗传背景、构图群体和生长环境的差异影响,只能反映出少量杂交组合中产量相关性状的遗传信息,局限性还比较明显。
与上述研究结果不同,本试验在豇豆群体水平上,通过SSR分子标记与豇豆产量相关性状的关联分析,发掘出一批与产量性状显著关联的SSR标记位点,揭示了豇豆基因组中产量性状的遗传位点分布特点。这些与表型性状关联的SSR标记在豇豆基因组上的分布不均,主要分布在连锁群LG2、LG3、LG4、LG7、LG11上,其SSR分子标记对关联性状的表型变异解释率范围在9%~33%,解释率越高说明关联程度越大;其中,千粒重的解释率最低(9%),鲜荚单荚重的解释率最高(33%)。进一步分析发现,同一个性状有多个SSR标记与之相关联,包括叶宽有2个SSR标记与之关联(标记位点CLM0251和CLM0850),鲜荚单荚重有3个SSR标记与之关联(标记位点CLM0347、CLM0251、CLM0614);同一个标记也可有多个性状相关联,例如,标记位点CLM0251与叶宽、鲜荚单荚重均有关联。这可能是性状相关基因多效应的遗传基础。
总之,试验采用SSR分子标记在豇豆中开展了全基因组关联分析研究,发掘出一批与豇豆产量性状关联的SSR分子标记,这将为在豇豆育种中开展关联分析与分子标记辅助育种等研究提供借鉴。
参考文献:
[1] THORNSBERRY J M, GOODMAN M M, DOEBLEY J, et al. Dwarf 8 polymorphisms associate with variation in flowering time[J]. Nature Genetics, 2001, 28: 286-289.
[2] FLINT-GARCIA S A, THORNSBERRY J M, BUCKLER E S. Structure of linkage disequilibrium in plants [J]. Annual Review of Plant Biology, 2003, 54: 357-374.
[3] YU J, PRESSOIR G, BRIGGS W H, et al. A unified mixed-model method for association mapping that accounts for multiple levels of relatedness [J]. Nature Genetics, 2006, 38: 203-208.
[4] XU P, WU X H, WANG B G, et al. Genome wide linkage disequilibrium in Chinese asparagus bean (Vigna unguiculata ssp. sesquipedialis) germplasm implications for domestication history and genome wide association studies [J]. Heredity, 2012, 109: 34-40.
[5] ROSTOKS N, RAMSAY L, MACKENZIE K,et al. Recent history of artificial outcrossing facilitates whole-genome association mapping in elite inbred crop varieties [J]. Proceedings of the National Academy of Sciences USA,2006,103:18656-18661.
[6] IWATA A, GREENLAND C M, JACKSON S A. Cytogenetics of legumes in the phaseoloid clade [J].The Plant Genome,2013,6(3):1-8.
[7] MEN?魪NDEZ C M, HALL A E, GEPTS P. A genetic linkage map of cowpea (Vigna unguiculata) developed from a cross between two inbred, domesticated lines [J]. Theoretical and Applied Genetics, 1997, 95(8): 1210-1217.
[8] OU?魪DRAOGO J T, GOWDA B S, JEAN M, et al. An improved genetic linkage map for cowpea (Vigna unguiculata L.) combining AFLP, RFLP, RAPD, biochemical markers, and biological resistance traits [J].Genome,2002,45:175-188.
[9] MUCHERO W,DIOP N N, BHAT P R,et al. A consensus genetic map of cowpea [Vigna unguiculata (L) Walp.] and synteny based on EST-derived SNPs [J].P Natl Acad Sci USA,2009, 106: 18159-18164.
[10] LUCAS M R, DIOP N N,WANAMAKER S,et al.Cowpea-soybean synteny clarified through an improved genetic map[J].Plant Genome,2011,4:218-225.
[11] XU P, WU X H,WANG B G,et al. A SNP and SSR based genetic map of asparagus bean (Vigna unguiculata ssp. sesquipedialis) and comparison with the broader species [J]. Plos One,2011-01-06, DOI:10.1371/journal.pone.0015952.
[12] KONGJAIMUN A, KAGA A, TOMOOKA N, et al. An SSR-based linkage map of yardlong bean [Vigna unguiculata (L.) Walp. subsp. unguiculata cv. Sesquipedalis group] and QTL analysis of pod length[J].Genome,2012b,55: 81-92.
[13] 王佩芝,李锡香.豇豆种质资源描述规范和数据标准[M].北京:中国农业出版社,2005.27-33.
[14] 潘 磊,郑 鹏,徐 杰,等.磁珠富集制备莲藕基因组的微卫星分子标记[J].中国蔬菜,2007(S):7-13.
[15] EVANNO G, REGNAUT S, GOUDET J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study [J]. Molecular Ecology, 2005, 14(8): 2611-2620.
[16] HARDY O J,VEKEMANS X. SPAGeDi:A versatile computer program to analyse spatial genetic structure at the individual or population levels[J].Molecular Ecology Notes,2002,2: 618-620.
[17] BRADBURY P J, ZHANG Z, KROON D E, et al. TASSEL: Software for association mapping of complex traits in diverse samples[J].Bioinformatics,2007,23: 2633-2635.
[18] TOMOOKA N,NAITO K, KAGA A,et al.Evolution,domestication and neo-domestication of the genus Vigna[J].Plant Genetic Resources,2014, 12(S1): 168-171.
[19] BOHRA A,PANDEY M K, JHA U C,et al. Genomics-assisted breeding in four major pulse crops of developing countries: Present status and prospects[J].Theoretical and Applied Genetics,2014,127(6):1263-1291.
[20] KONGJAIMUN A,KAGA A,TOMOOKA N,et al. The genetics of domestication of yardlong bean,Vigna unguiculata (L.) Walp. ssp. unguiculata cv. Sesquipedalis [J].Annals of Botany,2012a,109:1185-1200.
[21] XU P,WU X H,WANG B G,et al. QTL mapping and epistatic interaction analysis in asparagus bean for several characterized and novel horticulturally important traits [J]. BMC Genetics,2013,14(4):1-10.
[22] ANDARGIE M,PASQUET R S,MULUVI G M,et al. Quantitative trait loci analysis of flowering time related traits identified in recombinant inbred lines of cowpea (Vigna unguiculata) [J].Genome,2013,56:289-294.
猜你喜欢
中老年保健(2022年7期)2022-09-20
保鲜与加工(2022年8期)2022-09-01
电脑迷(2020年8期)2020-12-16
电脑迷(2020年7期)2020-12-16
趣味(数学)(2020年4期)2020-07-27
支部建设(2020年15期)2020-07-08
中国与非洲(法文版)(2019年7期)2019-07-31
种子(2018年6期)2018-07-10
上海蔬菜(2016年5期)2016-02-28
新疆农垦科技(2014年10期)2014-02-28
