
青藏高原高寒草甸钝裂银莲花种子特征研究
2015-10-26 09:11刘左军薛德艳蓝雯琳
中国食品工业 2015年12期
高 望,刘左军,薛德艳,王 强,蓝雯琳
(兰州理工大学 生命科学与工程学院,兰州730050)
青藏高原高寒草甸钝裂银莲花种子特征研究
高 望1,刘左军*,薛德艳2,王 强2,蓝雯琳2
(兰州理工大学 生命科学与工程学院,兰州730050)
以青藏高原高寒草甸具有花色多态性的钝裂银莲花为材料,研究其种子形态、种子大小、种子萌发率、发芽势和贮藏时间对种子生活力的影响。结果表明:三种花色钝裂银莲花种子在种子形态、种子大小以及萌发率方面无显著差异;在种子发芽势上白色居群种子与其他两色种子之间有显著差异,可能与白色居群物候期较早有关;种子贮藏时间长度与种子生活力负相关,但是其种子经过长期保藏依旧具有一定种子活力。种子以稳定的形态和大小遗传、较高的萌发率和种子持久的生活力是其对恶劣环境的一种适应。
钝裂银莲花;花色;种子特征
植物种子在植物的种族延续上,既是植物遗传信息的保存者与传递者,也是植物在环境胁迫中保证物种繁衍和繁殖的一种适应性策略[1,2]。在植物生活史过程中,通过最佳的资源分配格局,来提高植物适合度[3]。植物对种子的资源投入作为植物总的资源分配的一部分,主要体现在种子的数量、重量、结构及着生位置上,而不同的资源分配策略也是对所处环境的一种适应[4-6]。钝裂银莲花(Anemone obtusiloba)是毛茛科(Ranunculaceae)银莲花属(Anemone L.)多年生草本植物,其花分为黄色(yellow)、浅黄色(pale-yellow)和白色(white),具有花色多态性,是高寒草甸草地退化的一种指示性毒杂草。本文通过研究比较三种花色钝裂银莲花种子形态及大小、种子萌发率和不同贮藏时间对种子生活力的影响,旨在为进一步揭示其繁衍机制、预测其潜在分布范围以及综合治理提供依据,这对于高寒草甸的植被恢复有着极其重要的意义。

1、材料与方法
试验材料采集地点在甘肃省甘南藏族自治州合作市高寒草地与湿地系统定位研究站(102°53′E,34°57′N),海拔2973m。年平均气温2.0℃,最冷的1月份平均气温为-9.0℃,6~8月最热,其平均气温为11.6℃。年均降雨量400~800mm,年蒸发量1200mm,降水主要分布在6~9月。年日照时数2560h。年均无霜期≤90天。植被类型以高寒草甸为主,土壤为亚高山草甸土。
1.1种子形态特征及千粒重
样地内分别选取三种花色钝裂银莲花各50株并挂牌标记,进入果期时使用无纺布袋进行套袋处理,防止种子脱落不整齐性造成的种子遗失,并在9月下旬种子成熟后按照花色分类回收种子并带回实验室,用游标卡尺测量种子形态,即种子的长、宽、厚。每种花色种子测量8组,每组100粒种子。对不同花色植株种子,按照单株种子进行称重,计算每种花色种子的千粒重,精确到万分之一克。
1.2种子萌发率和发芽势
选取大小、重量相对一致的三种花色的钝裂银莲花种子。分别将种子用40目尼龙袋装好,封紧袋口,于2013年10月埋回合作站点内,埋藏深度为1cm,进行春化处理。在2014年4月,回收埋藏种子,带回实验室进行萌发实验。采用纸上萌发法,将钝裂银莲花种子消毒并冲洗后,在超净台内接种至已经灭菌、铺有两层用蒸馏水湿润的滤纸的培养皿中进行萌发,并记录称重。每个培养皿接入50粒钝裂银莲花种子,实验设置三个平行。萌发条件为恒温20℃,8h黑暗,16h光照。每天统计其萌发率,称重法补水,并将已萌发(胚根突破种皮2mm,则视为已萌发)的种子移出,计算发芽势。连续7d不再有种子发芽结束发芽试验。萌发实验结束后,采用TTC染色法鉴定种子活力。
萌发率=发芽结束时种子发芽数/供试种子数×100%
发芽势=3d内种子的发芽数/供试种子数×100%
1.3贮藏时间对种子生活力的影响
采用当年种子收获后去除杂质,封装于纸袋内,室内常温贮藏1-4年的不同花色的钝裂银莲花种子。种子生活力检测采用TTC染色法。将钝裂银莲花按照年份和花色分类后,放入用蒸馏水湿润的滤纸上,进行缓慢纸床预湿2h。预湿结束后,将种子剥去种皮,将种子放入有1%TTC染液的培养皿中,使染液刚好浸没种子。将培养皿移至恒温箱内,20℃黑暗染色4h。染色结束后,将种子纵切,观察染色情况。
有生活力的种子数=该组被染色的种子数/该组供试种子数
以上数据用Microsoft Excel 2010及SPSS 20.0软件处理。
2、结果与分析
2.1种子形态特征及千粒重比较
如表1所示,钝裂银莲花种子在种子形态特征和千粒重方面均呈现黄色>浅黄色>白色的关系,但是三种花色之间无显著差异。

表1 三种花色的钝裂银莲花种子长、宽、厚和千粒重Table 1 Length, width, thickness and 1000-seed weight of seed of Anemone obtusiloba
2.2种子萌发率和发芽势比较
如图1所示,三种花色钝裂银莲花种子在萌发率都较高,其中黄色96.00%>白色95.33%>浅黄色94%,但是三种花色种子萌发率之间无显著差异。三种花色钝裂银莲花种子发芽势方面,白色84%显著大于黄色72%和浅黄色70%种子的发芽势,说明白色居群种子发芽整齐度高。
2.3贮藏时间对种子生活力的比较
如图2所示,随着贮藏时间的增加,钝裂银莲花种子的生活力在逐渐减弱,种子活力与贮藏时间负相关。如表2所示,三种花色钝裂银莲花花间与贮藏时间相互比较。

表2 钝裂银莲花种贮藏时间与生活力比较Table 2 Storage time and life force of Anemone obtusiloba
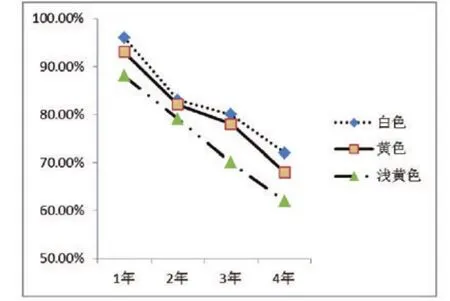
图1 钝裂银莲花种子发芽率和发芽势Fig 1 Germination rate and germination potential of Anomone obtusiloba

图2 贮藏时间对钝裂银莲花种子生活力的比较Fig 2 Storage time on seed-viability of Anomone obtusiloba
3、讨论
在自然条件下,种子特征是相对稳定的,由于可利用资源的变化而引起的种子生产受到影响时,一般首先是种子数量的影响,其次才是种子大小[7]。植物个体间以及个体内种子大小变异是一普遍现象,且种群内种子大小变异主要来源于各个个体间[8-10]。因此,在钝裂银莲花种子形态及千粒重方面,未检测到花色之间的显著差异。说明钝裂银莲花遗传上的进化具有相对稳定性。在高寒草甸这种比较特殊的生境条件下,具有花色多态性的钝裂银莲花可能就是凭着稳定的遗传特性在高寒草甸占有自己的独特的种群地位,成为一种优势的毒杂草。
钝裂银莲花种子具有较高的萌发率,只要满足了基本的萌发条件便可发芽并形成幼苗。但是其具有生理性休眠,不经过打破休眠的处理(如低温层积处理,自然状态下的春化处理及冷湿处理等),则其萌发率较低[11,12]。通过花色间种子萌发率的比较,发现花色之间种子萌发率不存在显著差异。但是在其发芽势上,说明的发芽整齐度较高,在适合的条件下能够尽快萌发。通过几年的观察,白色居群较之其他颜色居群有更早的物候期,出苗时间和开花时间都较早,也说明了白色花种子较高发芽势,能够较快萌发。
钝裂银莲花作为高寒草甸上生存的一种常见毒杂草,具有持久的生命力。原因可能是其种子在不适宜种子萌发的条件下具有休眠的特性,从而能够长期保持生命力而不丧失发芽能力。据报道[13,14],如车前草(Plantago asiatica) 的种子发芽力能保持10年之久;马齿苋(Portulaca oleracea)能在保藏20-40年之内还具有生活力。
钝裂银莲花作为高寒草甸上常见的一种毒杂草,在环境选择的压力下演化成为一种高度进化的植物种群,其繁殖过程中表现出来的花色多态性、种子特征的稳定遗传、种子萌发率较高、具有休眠过程保持种子生命力持久等,都是长期以来适应高寒草甸这严酷环境所形成的生殖对策。”CFI
[1] 祖元刚,赵则海,杨逢建,等.植物生活史型的划分及其相互转化[J].生态学报.2005,(09):2347-2353.
[2] 宋松泉,程红众,姜孝成等.种子生物学[M.]科学出版社,2009
[3] Fenner.,M.,Thompson. K. The Ecology of Seeds[M]. New York: CABI Publishing, 2005
[4] 李金花,潘浩文,王刚.草地植物种群繁殖对策研究[J].西北植物学报.2004,(02):352-355.
[5] 佘诚棋,杨万霞,方升佐,等.青钱柳天然群体种子性状表型多样性[J].应用生态学报.2009,(10):2351-2356.
[6] Maranz S,Wiesman Z.Evidence for indigenous selectionand distribution of the shea tree,Vitellaria paradoxa,and its potential significance to prevailing parkland savanna treepatterns in sub-Saharan Africa north of the equator[J]. Journalof Biogeography,2003,30:1505 -1516.
[7] 刘左军,杜国祯,陈家宽,等.影响黄帚橐吾种子生产的因素Ⅰ.生境和花序结构[J].植物生态学报.2003,(05):677-683.
[8] Pitelka L F, ThayerM E, Hansen S B. Variation in acheneweight in Aster acum inatus[J]. Can.J.Bot., 1983,61(5): 1415-1420.
[9] Thompson JN. Variation among individual seed masses in Lomatium grayi(Umbelliferae) under controlled conditions[J]. magnitude and partitioning of the variance.Ecol, 1984,65(2): 626-631.
[10]A anzen D H. Variation in seed size within a crop of a Costa Rican Mucuna and reana(Legum inosae)[J]. Am.J.Bot.,1977,64(3): 347-349.
[11] 路宁娜,崔现亮,王桔红,等.不同贮藏条件和光照对5种蒺藜科植物种子萌发的影响[J].中国沙漠.2008,(06):1130-1135.
[12]李富香,刘左军,高望等.不同贮藏环境和光照对钝裂银莲花种子萌发的影响[J].中国食品工业,2014,6:50-53
[13]李博.植物竞争-作物与杂草相互作用的实验研究[M].北京:高等教育出版社,2001.
[14]郭水良,王勇,曹同.杂草繁殖方式的多样性及其环境的适应[J].上海师范大学学报(自然科学版).2006,35(3):103-110
Study on seed characteristics of Anemone obtusiloba in Qinghai-Tibet plateau alpine meadow
GAO Wang1, LIU Zuo Jun*, XUE De Yan2, WANG Qiang2, LAN Wen Lin2
(School of Life Science and Engineering of Lanzhou University of Technology,Lanzhou,Gansu Province,730050,China)
∶ In Qinghai-Tibet plateau alpine meadow Anemone obtusiloba with color polymorphism as materials,seed shape,seed size, seed germination rate, germination energy and storage time on seed viability. The results showed that∶ three colors Anemone obtusiloba seed was no significant difference in seed shape, seed size and germination rate; there are significant differences between the white population and the other two color seed germination potential on seeds and white populations may phenophase earlier related; the length of seed storage time and negatively correlated with seed viability, but its longterm preservation seed after seed still has some vigor. Seeds in a stable form and size genetic, higher germination rate and seed lasting viability which is one of its adaptation to harsh environments.
∶ Anemone obtusiloba; color; seed characteristics

O633.13
A
0438-1157(2015)00-0000-00
国家自然科学基金资助项目(30960066)资助
高望(1988-),男,陕西西安人,在读硕士研究生。研究方向为生态学。
刘左军,教授,硕士生导师,主要从事生态学方向研究。
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
中国农业科学(2022年13期)2022-07-26
青海草业(2022年2期)2022-07-23
海外文摘·文学版(2021年3期)2021-05-19
现代园艺(2020年19期)2020-10-02
江苏农业科学(2020年21期)2020-03-08
种子(2019年6期)2019-07-13
种子(2019年4期)2019-05-28
活力(2019年21期)2019-04-01
现代农村科技(2019年4期)2019-01-06
