
江谷水库鱼类网箱养殖富营养化及浮游植物功能群的指示作用
2015-10-22 02:01:04蓝于倩袁一文彭亮胡韧
生态环境学报 2015年6期
蓝于倩,袁一文,彭亮, ,胡韧, *
1.暨南大学生态学系,广东 广州 510632;2.广东省水库蓝藻水华防治中心,广东 广州 510632
网箱养殖是一种利用湖泊、水库等大中型水域进行局部高密度的养殖行为(蒋高中,2008),因人工投喂而直接或间接地影响水体营养状态和水生态系统的健康。很多研究认为,鱼类养殖往往改变了水生生态系统的结构与功能(Brander,2007)、导致水体富营养化(Guo等,2009)、水质污染恶化(Tovar,2000)、浮游生物群落结构转变和浮游植物小型化(Borges等,2010;Guo和 Li,2003)等问题的发生。有研究表明鱼类养殖过程中产生的排泄物和饵料废渣加速了水体的营养盐循环,它们很容易矿化为可溶性营养盐,被浮游植物吸收利用而大量生长(Guo和 Li,2003)。Degefu等(2011)发现网箱养殖的鱼类排泄物造成了 Allage水库发生了拟鱼腥藻的水华。此外,大规模的网箱养殖还可能影响水体浮游动物结构,带来水体浮游植物小型化等问题(Sereda等,2008)。江谷水库网箱养殖以肉食性鳜鱼为主,根据经典生物操纵理论,肉食性鱼类可以通过级联效应调节滤食性鱼类种群数量,从而达到调控生态系统的目的(Sereda等,2008;Gido,2002;此里能布等,2012),但实际中网箱养殖鳜鱼的调控作用是有限的,加上避免不了投饵、施肥等增加水体营养物质的过程,因此网箱养殖加速水体富营养化(张国华等,1997)。传统的水化指标可以较好地反映水体营养盐的变化,但浮游植物的群落组成更能准确地反映富营养对水质的影响。经典的浮游植物分类调查虽然能够在一定程度上反映浮游植物群落结构组成及变化,但不能更精确描述浮游植物的生态学功能,需要采用更有效的指标来衡量网箱养殖对水库生态环境的作用。
浮游植物功能群(Functional Groups)作为基于生态功能的分类方法,将反映某一特定的生态环境特征的种类集中到同一个类群(胡韧等,2015),从而能更好地反映浮游植物群落的变化情况(Reynolds等,2002;Padisák 等,2009;Hu 等,2013)。目前国内外已广泛运用浮游植物功能群的分析方法来研究湖泊、水库、河流等多种水体环境的状况(Reynolds,1999;董静等,2013),或者指示同一水体在不同因素作用下的时空差异(Becker等,2009a;Caputo 等,2008;Becker等,2009b),但应用于网箱养殖水体的报道不多。网箱养殖对水库的影响是多方面的,单独运用传统的水化指标进行影响分析较为单一,因而结合理化指标和生物指标进行综合分析尤为重要(Degefu等,2011)。而浮游植物功能群的分析不仅能够在一定程度上反映浮游植物群落结构组成及变化,而且还能更精确描述浮游植物的生态学功能,有效地衡量网箱养殖对水库生态环境的作用。
江谷水库是四会市重要的后备水源地,但在北库区存在网箱养殖,可能对水质及水生态系统造成一定影响。本研究在 2014年丰水期和枯水期,对江谷水库不同区域进行观测和对比,结合水质数据和浮游植物功能群分析方法,分析鱼类网箱养殖与富营养化和浮游植物功能群的关系,以期揭示网箱养殖对江谷水库水质的影响及其时空特征。
1 材料与方法
1.1 江谷水库及其采样位点
江谷水库地处于广东省肇庆市北部的四会市江谷镇,建于1960年,最大库容为6126万m3,是一座集防洪、灌溉、发电、供水、鱼类养殖等多种功能综合利用的中型水库。水库流域面积 137 km2,平均水深达11.69 m。上游主河流发源于广宁县十排山南麓,经由广宁县谭布镇、四会县江林镇注入水库,水库下游长30.8 km,向东南经过江谷、龙江等乡镇,最后注入绥江。江谷主要经济收入依靠发电和灌溉供水,还兼顾了防洪和渔业养殖。水库周边种植较多桉树和竹林,在个别库湾中围养鸭、鹅等禽类。由于流域存在污染源和库区的鱼类养殖,江谷水库富营养化较严重,近年来出现了水色发黑的现象,并在 2014年春节期间爆发了小规模蓝藻水华。
江谷水库属于亚热带季风气候温和多雨,具有明显的枯水期和丰水期。枯水期,水库网箱养殖区域分布于水库北部的河流-过渡区;而丰水期随着雨水增多、水位上升,网箱增多,养殖区域向上游河流区和下游大坝区扩展。网箱内主要养殖肉食性鳜鱼。
根据江谷水库的水文特征以及库区内鱼类网箱养殖情况,调查设置了丰水期和枯水期两次对比采样,并在水库内设置了7个采样点S1-S7(图1)。其中,S1-S3归为南库区,S4为大坝区,S5-S7为北库区(网箱区)。于 2014年3月26─27日和9月26日进行2次采样,样品进行理化指标的测定和浮游植物的定性定量分析。

1.2 水质指标的测定
用YSI-85型水质仪现场测定水库水体的温度(t)、pH值等指标,用萨氏盘测定透明度(SD),同时记录采样的位置、气温。采集水面以下0.5 m水体,采用国家水质标准方法进行总氮、硝氮、氨氮、总磷等水化指标的测定(国家环保总局,2003);叶绿素测定采用改进的反复冻融-浸提法(林少君等,2005)。浮游植物定性定量分析。浮游植物定性,用25号浮游生物网进行水平、垂直拖取;定量样品则采集0.5 m处水样,加入4%甲醛固定,沉淀浓缩后,在光学显微镜下进行藻类的鉴定和计数。
1.3 数据处理与分析
富营养化指数(TSI)根据广东省水库调查结果得到的运算公式进行计算(Carlson,1977;Lin,2001)。
根据Padisák等(2009)和Reynolds等(2002)确定的浮游植物功能类群分类方法对浮游植物进行功能类群划分。调查的数据使用SPSS进行方差检验,数据经log(x+1)转化后,采用Past 3.06进行NMDS(Nonmetric Multidimensional Scaling)排序分析,并用Origin 8.0进行绘图。
2 结果与分析
2.1 理化环境
2.1.1 理化因子
结果表明,从南库区到网箱区,氮磷营养盐、电导率和叶绿素的浓度依次增加,而水体的透明度则逐步降低(图2)。水体溶解氧在丰水期各点差异不大,枯水期网箱区的溶解氧明显低于其它两个区域。季节上看,枯水期到丰水期,虽然网箱区域有所扩大,受降雨影响,总体营养盐浓度仍然降低,仅氨氮出现上升现象。非网箱区的叶绿素浓度在水温较高的丰水期要高于枯水期,而网箱区则是枯水期高于丰水期,这可能与其在枯水期较高的营养盐支持有关。

2.1.2 营养状态
本文选取总磷、总氮、透明度以及叶绿素a浓度4个因素作为综合评价水体营养状态的参数。从图3可以看出,在枯水期南库区为中营养,大坝区和网箱区为富营养,丰水期则仅网箱区 TSI(∑)为富营养。从南库区到大坝区再到网箱区,TSI指数逐步增高。枯水期到丰水期,TSI(TN)、TSI(TP)和TSI(SD)有不同程度的降低,TSI(Chla)有所上升,水库总体富营养化指数降低。
2.2 浮游植物群落
2.2.1 浮游植物的组成和丰度
两次采样共发现6个门70多种浮游植物。浮游植物的丰度和生物量的最大值没有出现在营养盐水平和富营养化指数最高的网箱区,而是在其下游的大坝区,其生物量最大值在丰水期和枯水期分别为 16.837和 17.599 mg·mL-1(图 4)。从枯水期到丰水期,江谷水库浮游植物在数量和组成上发生了很大变化:枯水期,浮游植物的平均生物量为7.920 mg·mL-1,丰水期则上升为 13.326 mg·mL-1(图 4);群落组成由硅藻为主转化为绿藻占据绝对优势,且物种组成单一化,角星鼓藻(Staurastrumsp.)在生物量上占据绝对优势。枯水期大坝生物量(平均17.599 mg·mL-1)明显高于网箱区(平均 4.06 mg·mL-1)和南库区(平均 2.101 mg·mL-1);在丰水期这种差异相对减小,大坝区(平均 16.837 mg·mL-1)和网箱区(平均 13.814 mg·mL-1)的生物量比较接近,南库区(平均9.328 mg·mL-1)最低。


2.2.2 浮游植物功能群的组成
两次采集的浮游植物可以归入24个功能群(表1)。江谷水库浮游植物受季节气候影响大于空间影响,同一时期里北库区(网箱区)、南库区和大坝区的主优势功能群(相对比例>10%)组成相似。丰水期为功能群NA、J、K、F占据优势,枯水期则为功能群K、J、C、S1占据优势(图5),说明枯水期水体普遍高营养、停滞和浑浊,丰水期虽然营养水平降低、透明度有所提高但仍处于中-富营养水平。因此,仅从主优势功能群组成差异分析网箱养殖对于水体的影响并不明显,需要利用浮游植物次优势功能群(5%<相对比例<10%)的差异进行分析(图5)。

表1 浮游植物功能群的划分Table 1 Differentiation of phytoplankton functional groups

续表1Continued Table 1
表2列举了各区域的特有次优势功能群。枯水期,耐浑浊低光照的富营养功能群LO、D、P在网箱区占据优势,能迅速利用营养盐的X1和D在南库区占据优势,耐高光照的G和A在大坝区和南库区占据优势。以上功能群的组成,除了反映水库水体整体营养水平高的特征之外,还能较好地区分各个区域的特征:网箱区水体浑浊程度高、适合耐受低光照的浮游植物生长;大坝区水体浊度最低、可获得性光照强度大;南库区则处于二者之间。丰水期,网箱区与大坝区的浮游植物次优势功能群组成接近,均由适合高营养、耐受低光照的功能群S1、M、C组成,而南库区没有特有的次优势功能群,反映了网箱区影响扩大、营养水平和浑浊程度都较高。
各区域的浮游植物功能群组成的 NMDS分析表明,营养盐水平较低的南库区的样点分布在坐标系靠近X轴一侧,而浮游植物细胞丰度最高的两个样点——枯水期的大坝区和丰水期的网箱区,均分布在坐标系的上端(图6)。枯水期和丰水期的样点分别聚集在图的两侧,说明浮游植物受季节因素影响大,不同水期之间的差异大于空间差异。枯水期出现以K、J、G、C、S1、LO和A为优势(丰度达到5%以上)的21功能群,到丰水期时则转变为以NA、J、K、S1、F和M为优势(丰度达到5%以上)的 17个功能群。功能群 K(以色球藻Chroococcussp.、蓝纤维藻Dactylococcopsissp.为主)、J(以栅藻Scenedesmussp.、十字藻Crucigeniasp.为主)、S1(假鱼腥藻Pseudanabaenasp.、泽丝藻Limnothrixsp.为主)在枯、丰水期都占据较好的生长优势;枯水期的优势功能群G、C、LO、A在丰水期为功能群NA、F、M替代。此外,功能群A、TD、TB、MP、E、W1、WS、S2为枯水期特有,功能群SN、H1、M、LM和G2为丰水期特有。
NMDS排序图显示(图6)枯水期3个样点相隔较远,说明样点间浮游植物功能群存在较大差异;丰水期的网箱区和大坝区样点相互靠近,这与丰水期网箱养殖面积扩大,致使网箱区的影响扩至大坝区有关,两个样点的浮游植物功能群组成开始趋同。
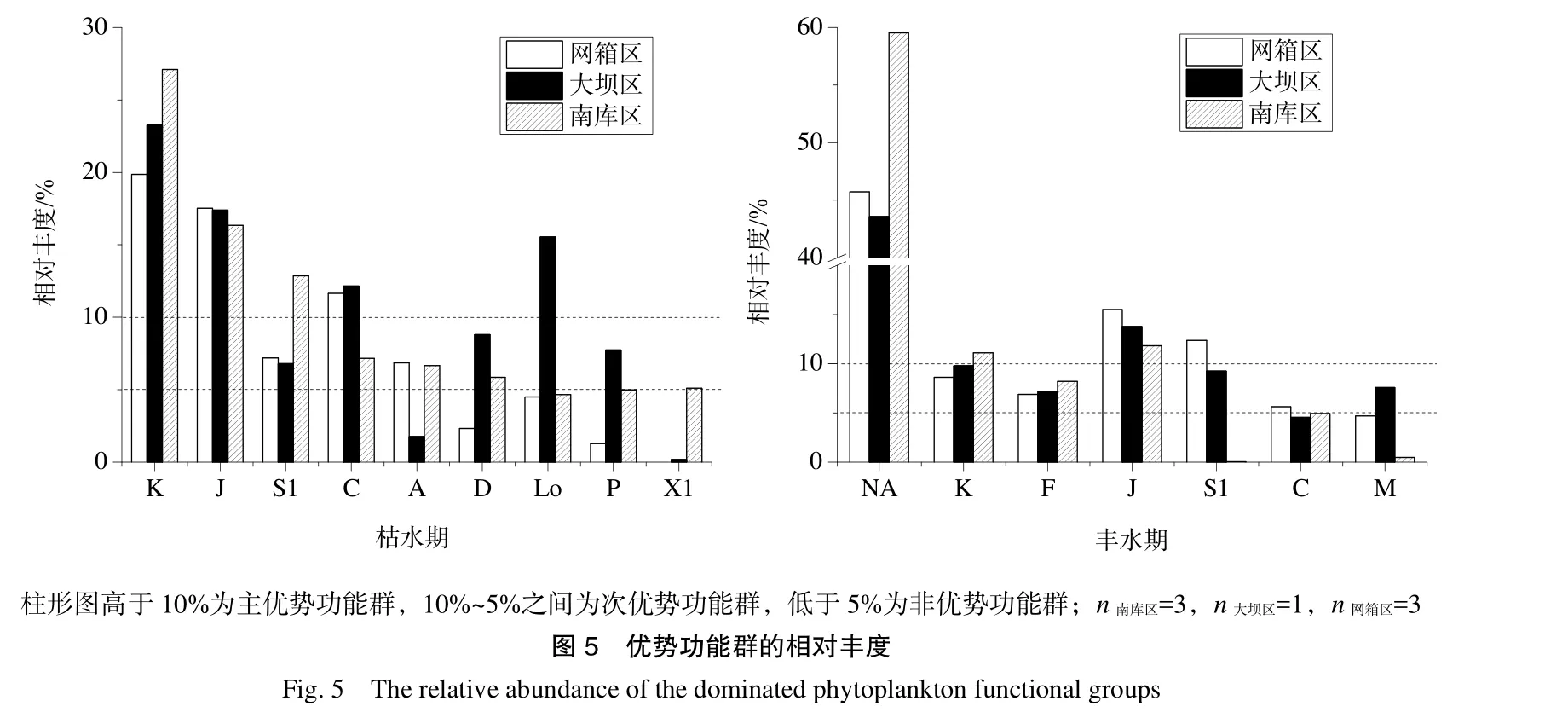

表2 丰/枯水期各区域次优势浮游植物功能群Table 2 Sub-dominant phytoplankton functional groups of different areas and times

3 讨论
3.1 网箱养殖对水库富营养化的影响
鱼类网箱养殖是很多水库的重要功能和经济来源之一。80年代以前,网箱养殖是人们动物蛋白的重要来源,如何通过鱼类养殖增产曾作为水库研究和管理的一个重要内容(Degefu等,2011)。然而,随着社会经济的发展,网箱养殖规模不断扩大、养殖密度和鱼类产量增加,作为对水生态系统强烈干预的鱼类养殖,在产生经济效益之余,也给水库带来了生态风险(Boyd,2003;Garcia等,2014;Li等,2014)。根据TSI富营养指数计算,江谷水库网箱区的营养水平明显高于水库其他区域,尤其丰水期养殖面积扩大后,氮磷营养盐的水平明显高于其他区域(P=0.001),说明鱼类养殖确实是水库富营养化加重的重要来源。Vanni(2002)和Schaus等(1997)研究也证实了鱼类养殖明显提高了水体的氮磷浓度。鱼类网箱养殖最直接结果就是其代谢废物加速了水体营养盐的循环过程,增加藻类的可获得性营养盐,从而导致水体富营养化(李琼丽,2014)。在鱼类养殖的新陈代谢过程中,水库水体成为了氮、磷的储存库(Garnier等,2000;Nowlin等,2005),其输入的速度远大于输出,致使营养水平的上升无法避免。我们的研究也表明,从枯水期到丰水期,随着网箱养殖的面积扩大,水体中溶解态的氮磷增加,尤其是氨氮浓度明显升高。氨氮是浮游植物最易吸收利用的氮源,其主要来源就是鱼类的有机代谢产物。
3.2 浮游植物功能群对网箱养殖的指示
从本质上来说,水库发生富营养化是营养盐水平较高,导致水体初级生产力过剩的表现。在大部分传统的研究中,并没有直接采用浮游植物的生物量,而是通过以营养盐为主导的TSI综合营养指数来衡量水体水环境的优劣(Cai等,2002;朱广伟,2008)。林秋奇等的研究也表明,在南亚热带水库中养殖富营养化与藻类水华的发生往往是相一致的(林秋奇等,2003a)。然而在本研究中,水体的富营养综合指数在丰水期较枯水期低,但浮游植物的生物量却更高,同时有害藻类的优势度也增加,出现了富营养化的理化指标和生物指标相悖的现象。这说明浮游植物的生长不仅仅依赖于营养盐,还依赖于水体的光照和水动力学条件等多个特征。林秋奇等在对流溪河水库的研究中也发现,受透明度和水流的影响,浮游植物丰度在营养盐较高的季节反而较低(林秋奇等,2003b)。因此,评价网箱养殖对环境造成的富营养化影响,不能简单地采用营养盐浓度或富营养指数评价,还需要考虑其与环境特征相适应的浮游植物功能群结构组成的变化。
江谷水库从枯水期到丰水期,适应高营养盐水平、对水体高度混合和高光照敏感的功能群K和J在水库大部分地区常年占据优势,反映出江谷水库水体常年处于较高营养水平、相对静止且透明度不高。从浮游植物功能群的季节性差异来看,枯水期耐受低光照的丝状体组成的功能群C和 S1在进入丰水期后优势度减少,并被适应洁净环境、可耐受低营养的功能群NA、F替代,这与说明丰水期雨水增多、水位抬升提高了水体的洁净程度,导致透明度增加、营养水平降低的环境特征是相适应的(Padisák 等,2009)。
网箱养殖带来的低光照、高营养盐的水环境特征导致其在浮游植物功能群的组成上和其它两个区域存在明显的空间差异。网箱区的特有优势功能群LO和P,比其他区域的优势功能群具有更强的耐受低光照的能力,且适应一定程度水体扰动混合作用。Bartozek等(2014)在巴西水库鱼类养殖影响研究的结果显示,网箱养殖区水体浑浊度稍高,但由于一般理化指标的差异并不显著,往往容易被忽略。网箱养殖在枯水期导致局部水体浑浊,此时浮游植物的生长主要受光限制,加上温度较低生物量处于较低水平。丰水期,随着网箱养殖面积的扩大,浮游植物功能群在网箱区和大坝之间的空间差异缩小。网箱区与大坝区都由功能群S1、C、M组成。其中功能群 S1由丝状蓝藻组成、耐浑浊低光照生境(Vieira等,2015),C为由浮游硅藻组成、耐受低光照和低含碳量的富营养生境,M 由具有伪空胞和胶被的微囊藻组成、适应超富营养生境(Wiśniewska 和 Paczuska,2015),都具有适应富营养水体生境的偏好和应对浑浊水体的机制(Padisák等,2009;Belkinova等,2014)。这些浮游植物功能群的出现,指示出网箱区和大坝区的环境接近,具有较高的营养水平和较低透明度。丰水期NA功能群(鼓藻)在全库区的优势则归因于其耐受较低营养盐的能力,和南亚热带水体夏季特有的日混合现象(Souza等,2008;冯喻等,2014)。
综上所述,与从富营养综合指数得出的枯水期高,丰水期低的结果相反,浮游植物功能群有效地反映了江谷水库在丰水期发生有害藻类(S1和M等蓝藻功能群)水华风险更高的实际情况。
4 结论
(1)从TSI指数来看,网箱区明显高于其他区域,说明网箱养殖加重了江谷水库的水体富营养化程度;
(2)浮游植物功能群的NMDS分析表明,网箱养殖对水体的影响范围在丰水期较大;
(3)虽然江谷水库枯水期富营养化TSI综合指数较高,但浮游植物功能群的研究表明江谷水库在丰水期发生富营养化和藻类水华的风险更高。
BARTOZEK E C R, BUENO N C, RODRIGUES L C.2014.Influence of fish farming in net cages on phytoplankton structure: a case study in a subtropical Brazilian reservoir [J].Brazilian Journal of Biology, 74(1):145-155.
BECKER V, DE SOUZA CARDOSO L, HUSZAR V L M.2009.Diel variation of phytoplankton functional groups in a subtropical reservoir in southern Brazil during an autumnal stratification period [J].Aquatic Ecology, 43(2): 285-293.
BECKER V, HUSZAR V L M, CROSSETTI L O.2009.Responses of phytoplankton functional groups to the mixing regime in a deep subtropical reservoir [J].Hydrobiologia, 628(1): 137-151.
BELKINOVA D, PADISAK J, GECHEVA G, et al.2014.Phytoplankton based assessment of ecological status of bulgarian lakes and comparison of metrics within the water framework directive [J].Applied ecology and environmental research, 12(1): 83-103.
BORGES P A F, TRAIN S, DIAS J D, et al.2010.Effects of fish farming on plankton structure in a Brazilian tropical reservoir [J].Hydrobiologia,649(1): 279-291.
BOYD C E.2003.Guidelines for aquaculture effluent management at the farm-level [J].Aquaculture, 226(1): 101-112.
BRANDER K M.2007.Global fish production and climate change [J].Proceedings of the National Academy of Sciences, 104(50):19709-19714.
CAI Q, LIU J, KING L.2002.A comprehensive model for assessing lake eutrophication [J].Ying yong sheng taixuebao= The journal of applied ecology/Zhongguo sheng tai xuexuehui, Zhongguokexueyuan Shenyang yingyong sheng tai yanjiusuozhu ban, 13(12): 1674-1678.
CAPUTO L, NASELLI-FLORES L, ORDONEZ J, et al.2008.Phytoplankton distribution along trophic gradients within and among reservoirs in Catalonia (Spain) [J].Freshwater Biology, 53(12):2543-2556.
CARLSON R E.1977.A trophic state index for lakes1 [J].Limnology and oceanography, 22(2): 361-369.
DEGEFU F, MENGISTU S, SCHAGERL M.2011.Influence of fish cage farming on water quality and plankton in fish ponds: A case study in the Rift Valley and North Shoa reservoirs, Ethiopia [J].Aquaculture,316(1): 129-135.
GARCIA F, KIMPARA J M, VALENTI W C, et al.2014.Emergy assessment of tilapia cage farming in a hydroelectric reservoir [J].Ecological Engineering, 68: 72-79.
GARNIER J, BILLEN G, SANCHEZ N, et al.2000.Ecological functioning of the Marne reservoir (upper Seine basin, France) [J].Regulated Rivers: research and management, 16(1): 51-71.
GUO L, LI Z, XIE P, et al.2009.Assessment effects of cage culture on nitrogen and phosphorus dynamics in relation to fallowing in a shallow lake in China [J].Aquaculture International, 17(3): 229-241.
GUO L, LI Z.2003.Effects of nitrogen and phosphorus from fish cage-culture on the communities of a shallow lake in middle Yangtze River basin of China [J].Aquaculture, 226(1): 201-212.
GIDO K B.2002.Interspecific comparisons and the potential importance of nutrient excretion by benthic fishes in a large reservoir [J].Transactions of the American Fisheries Society, 131(2): 260-270.
HU R, HAN B, NASELLI-FLORES L.2013.Comparing biological classifications of freshwater phytoplankton: a case study from South China [J].Hydrobiologia, 701(1): 219-233.
LIN Q Q, HAN B P, LIN J X.2001.Reservoir water supply and reservoir eutrophication in Guangdong Province (South China) [C]//International Lake Environment Committee.Proceedings of 9th International Conference on the Conservation and Management of Lakes.Shiga,Japan: Shiga Profectual Government, 3(1): 269-272.
LI N, ZHANG X, WU W, et al.2014.Occurrence, seasonal variation and risk assessment of antibiotics in the reservoirs in North China [J].Chemosphere, 111: 327-335.
NOWLIN W H, EVARTS J L, VANNI M J.2005.Release rates and potential fates of nitrogen and phosphorus from sediments in a eutrophic reservoir [J].Freshwater Biology, 50(2): 301-322.
PADISÁK J, CROSSETTI L, NASELLI-FLORES L.2009.Use and misuse in the application of the phytoplankton functional classification: a critical review with updates [J].Hydrobiologia, 621(1): 1-19.
REYNOLDS C S, HUSZAR V, KRUK C, et al.2002.Towards a functional classification of the freshwater phytoplankton [J].JOURNAL OF PLANKTON RESEARCH, 24(5): 417-428.
REYNOLDS C S.1999.Phytoplankton assemblages in reservoirs [J].Theoretical reservoir ecology and its applications.Backhuiss Pub/Brazilian Academy os Sciences, São Paulo, 439: 456.
SCHAUS M H, BREMIGAN M T, GARVEY J E, et al.1997.Nitrogen and phosphorus excretion by detritivorous gizzard shad in a reservoir ecosystem [J].Limnology and Oceanography, 42(6): 1386-1397
SEREDA J M, HUDSON J J, TAYLOR W D, et al.2008.Fish as sources and sinks of nutrients in lakes [J].Freshwater Biology, 53(2): 278-289.
SOUZA M B G, BARROS C F A, BARBOSA F, et al.2008.Role of atelomixis in replacement of phytoplankton assemblages in Dom Helvécio Lake, South-East Brazil [J].Hydrobiologia, 607(1): 211-224.
TOVAR A, MORENO C, MÁNUEL-VEZ M P, et al.2000.Environmental impacts of intensive aquaculture in marine waters [J].Water Research,34(1): 334-342.
VANNI M J.2002.Nutrient cycling by animals in freshwater ecosystems[J].Annual Review of Ecology and Systematics, 33: 341-370.
VIEIRA P C S, CARDOSO M M L, COSTA I A S.2015.Vertical and temporal dynamics of phytoplanktonic associations and the application of index assembly in tropical semi-arid eutrophic reservoir,northeastern Brazil [J].ActaLimnologicaBrasiliensia, 27(1): 130-144.
WIŚNIEWSKA M, PACZUSKA B.2015.Long-term changes in the dynamics and structure of cyanobacteria in KoronowoReservoir [J].Oceanological and Hydrobiological Studies, 44(1): 127-138.
此里能布, 毛建忠, 黄少峰.2012.经典与非经典生物操纵理论及其应用[J].生态科学, 31(1): 86-90.
董静, 李艳晖, 李根保, 等.2013.东江水系浮游植物功能群季节动态特征及影响因子[J].水生生物学报, 37(5): 836-843.
冯喻, 肖利娟, 韦桂峰, 等.2014.高州水库水位上升期坝前深水区浮游植物群落的响应特征[J].应用与环境生物学报, 20(3): 529-536.
国家环保总局.2003.地表水和污水检测规范(HJ/T91-2002)[M].北京:中国标准出版社.
胡韧, 蓝于倩, 肖利娟, 等.2015.淡水浮游植物功能群的概念、划分方法和应用, 27(1): 11-23.
蒋高中.2008.20世纪中国淡水养殖技术发展变迁研究[D].南京农业大学.
李琼丽.2014.探讨滤食性鱼类在淡水渔业中的碳汇作用[J].农业与技术, 2014(2): 142.
林秋奇, 胡韧, 段舜山, 等.2003.广东省大中型供水水库营养现状及浮游生物的响应[J].生态学报, 23(6): 1101-1108.
林秋奇, 胡韧, 韩博平.2003.流溪河水库水动力学对营养盐和浮游植物分布的影响[J].生态学报, 23(11): 2278-2284.
林少君, 贺立静, 黄沛生, 等.2005.浮游植物中叶绿素 a 提取方法的比较与改进[J].生态科学, 24(1): 9-11.
张国华, 曹文宣, 陈宜瑜.1997.湖泊放养渔业对我国湖泊生态系统的影响[J].水生生物学报, (3): 271-280.
朱广伟.2008.太湖富营养化现状及原因分析[J].湖泊科学, 20(1):21-26.
猜你喜欢
中国资源综合利用(2024年7期)2024-12-31 00:00:00
食品安全导刊(2024年5期)2024-06-26 00:00:00
食品安全导刊·中旬刊(2023年4期)2023-05-31 21:35:50
海洋通报(2022年4期)2022-10-10 07:41:48
皮革制作与环保科技(2020年14期)2020-03-17 07:16:04
水利技术监督(2017年6期)2017-12-19 13:28:17
环境保护与循环经济(2017年10期)2017-03-16 03:16:20
中国农业文摘-农业工程(2016年5期)2016-04-12 05:38:07
广东海洋大学学报(2015年3期)2015-12-22 10:05:28
应用海洋学学报(2015年3期)2015-11-22 07:39:10
