
水杨酸对菊花耐热性的影响
2015-10-20 00:21向地英等
江苏农业科学 2015年9期
向地英等

摘要:以切花菊神马为试验材料,分别喷施浓度为0、100、200、400、800 mg/L的水杨酸,研究水杨酸对神马菊耐热性的影响。结果表明,随着温度升高,神马菊的茎叶干物质含量、叶绿素含量大体上呈先升高后降低趋势;喷施100、200 mg/L水杨酸可以提高神马菊的耐热性,最高可耐45 ℃的高温;超过42 ℃,神马菊的叶绿素含量和干物质含量下降,但是喷施200 mg/L水杨酸处理的神马菊,其叶绿素含量、茎叶干物质含量仍维持相对较高的水平。水杨酸虽然不能根本阻止高温胁迫而避免菊花受伤害,但喷施适宜浓度(200 mg/L)水杨酸可使菊花对热胁迫的耐受时间延长。
关键词:水杨酸;菊花;耐热性;干物质;叶绿素;神马;胁迫
中图分类号:S682.1+10.1文献标志码: A文章编号:1002-1302(2015)09-0222-02
菊花(Chrysanthemum morifolium Tzvel)为菊科菊属宿根花卉,是世界著名四大切花之一,性喜冷凉,持续高温会造成新生叶无法展开,叶片出现黄斑、反卷下垂、干枯等症状,影响菊花的正常生长发育,降低了其观赏价值[1],进而影响菊花的周年生产。为满足菊花的周年稳定供应,克服夏季高温对菊花的影响,菊花耐热性研究成为人们关注的课题。
水杨酸(salicylic acid,SA)是一种酚类物质,广泛存在于植物体内,参与植物体内许多生理生化过程,是植物体内诱导逆境防御机制的重要信号分子[2],能够提高植物在非生物胁迫下的抗性[3-5]。研究表明,外源水杨酸处理可增强高羊茅、茉莉、鸡冠花、葡萄、百合等植物对高温的耐受性[6-9],但对菊花耐热性研究却鲜有报道。本试验通过研究不同浓度水杨酸对切花菊神马耐热性的影响,旨在探讨水杨酸对提高菊花耐热性的有效性,为生产应用提供技术支持。
1材料与方法
1.1试验材料和水杨酸处理
以切花菊神马为试验材料,待植株长至约45~50 cm,选取生长一致的植株15畦,每畦约300株,于下午16:00—18:00分别喷施浓度为100、200、400、800 mg/L的水杨酸,以清水为对照。每周喷2次,处理2周,每处理重复3次。
1.2高温处理
剪取菊花地上部分约25 cm,用去离子水清除样品表面的污物,插入盛水的容器;试验设20、30、34、38、42、46、50 ℃ 共7个温度梯度,每个温度处理6株,计210株,放入智能人工气候箱培养。升温速率为5 ℃/h,到达每个温度梯度时保持4 h,常温放置1 d后测定各项指标。
1.3电导率测定
参考张钢等的方法[10]测定电导率。每株取4张叶,用5 mm 打孔器在叶片中间部位打1个孔;将打孔叶片分别置于4个试管中,每个试管加去离子水15 mL,用Parafilm膜封口,放入摇床中培养24 h;用上海京科雷磁产DDSL-308型数字电导仪测定电导率(E1)和空白电导值(EB1);将试管于沸水中煮沸30 min,Parafilm膜封口,再放入摇床中培养24 h,测定终电导值(E2)和空白电导值(EB2),计算相对电导率(E):
E=(E1-EB1)/(E2-EB2)×100%。
参照Logistic方程估算耐热性,计算公式为:
y=A1+eB·(C-x)+D[11]。
式中:x为处理温度,℃;y为相对电导率,%;A为处理的相对电导率最高值,%;B为温度拐点处曲线的斜率;C为拐点温度,表示组织半致死温度,℃;D为处理相对电导率最低值,%。
1.4叶绿素含量测定
参考张宪政的丙酮乙醇混合液法[12]测定叶绿素含量。取相同节位叶片样品0.2 g,放入洁净的胶卷盒中,分别加入丙酮乙醇混合液20 mL,混匀,避光保存24 h,待组织颜色变白,用分光光度计分别测定提取液在645、663 nm处的吸光度,计算叶绿素含量(C),公式为:
C(mg/g)=(20.2D645 nm+802D663 nm)×20÷1 000÷0.2。
1.5干物质含量测定
分别取茎、叶,放入4个纸袋中;用电子天平分别称量茎、叶样本的鲜质量;把茎、叶样本放入烘箱中105 ℃杀青 30 min,于60 ℃烘48 h;取出样本,放入干燥皿中24 h,用电子天平分别称量茎、叶样本干质量,计算干物质含量,公式为:
干物质含量=干质量/鲜质量×100%。
1.6数据处理
数据用Excel统计绘图,用SPSS 17.0分析差异显著性。
2结果与分析
2.1水杨酸处理对菊花茎干物质含量的影响
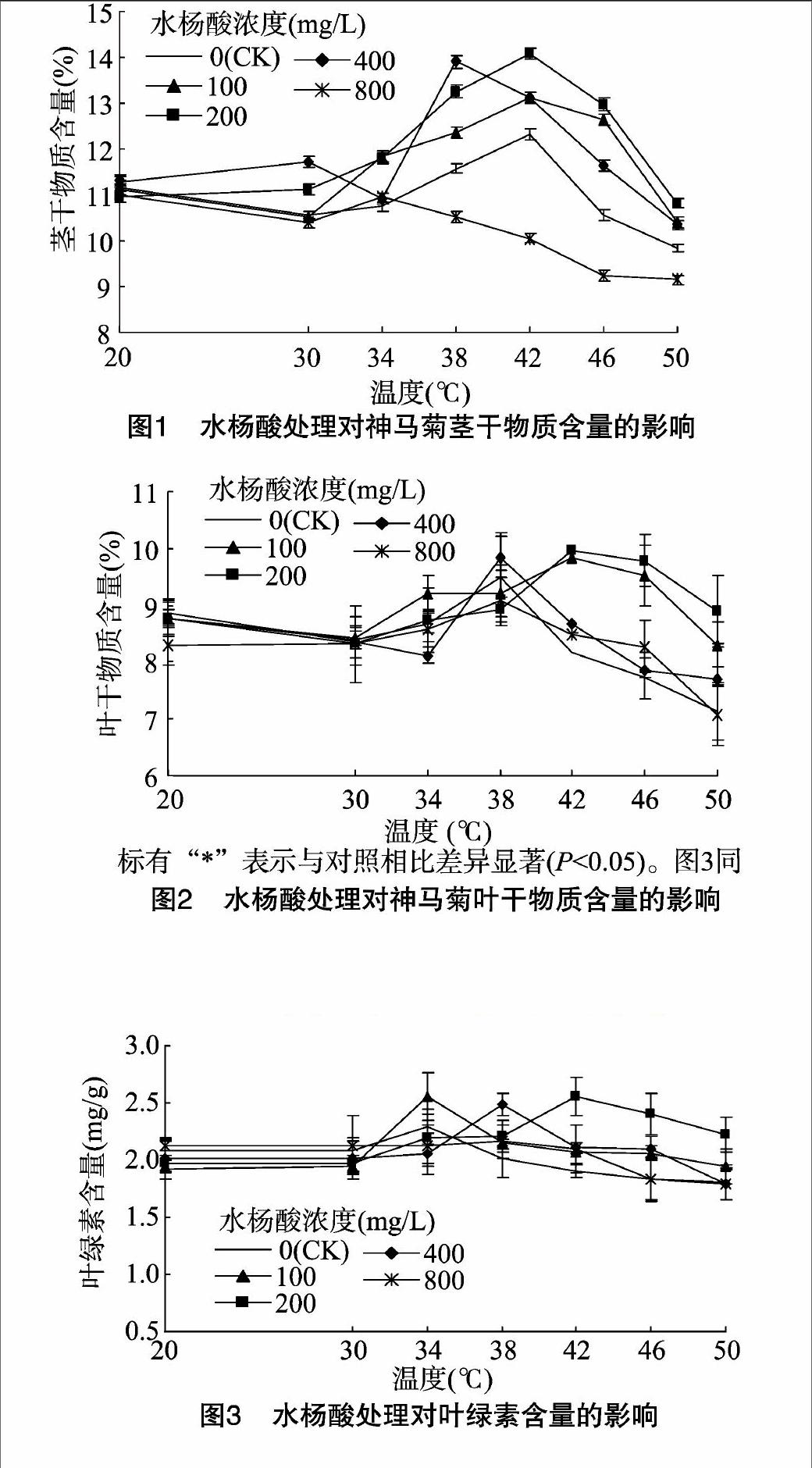
由图1可见,水杨酸处理的菊花,随处理温度的升高,茎干物质含量整体呈先升高后降低的趋势;38、42 ℃时,各处理茎干物质含量达到峰值,随后急剧下降,50 ℃时达最低值;各水杨酸处理的菊花在同一温度下,茎干物质含量差异不显著;200 mg/L 水杨酸处理的菊花,在超过34 ℃温度处理时,茎干物质含量均高于对照,而800 mg/L水杨酸处理的均低于对照。
2.2水杨酸处理对菊花叶干物质含量的影响
由图2可见,叶片的干物质含量变化走势与茎相似,大体上呈先升后降的趋势;20、30、34 ℃时,各水杨酸处理的叶干物质含量差异不明显;38 ℃时,400 mg/L水杨酸处理的叶片干物质含量最高;42、46、50 ℃温度下,200 mg/L水杨酸处理的叶干物质含量相对最高,均与对照有显著差异(P<0.05);100 mg/L水杨酸处理下,42 、46℃时叶干物质含量与对照有显著差异(P<0.05),更高温度处理的与对照无显著差异。
2.3不同浓度水杨酸对叶绿素含量的影响
由图3可见,水杨酸处理下,随处理温度的升高,神马菊叶绿素含量大致呈先上升后下降的趋势;未喷施水杨酸(对照)的,34 ℃时叶绿素含量开始下降;200 mg/L水杨酸处理,其叶绿素含量于42 ℃时达到峰值;20、30、34、38 ℃温度条件下,各处理与对照间均无显著性差异;在处理温度为42、46 ℃ 时,浓度为200 mg/L水杨酸处理的神马菊,其叶绿素含量与对照差异显著(P<0.05);400、800 mg/L水杨酸处理,在处理温度为46、50 ℃时,叶绿素含量与对照差异不显著。
2.4水杨酸对菊花耐热性的影响
由图4可见,经不同浓度水杨酸处理,菊花的耐热性发生明显变化,经100、200 mg/L水杨酸处理的菊花耐温度最强,分别为45.0 、45.5 ℃,耐热性分别比对照42.5 ℃高59%、7.1%;浓度为400、800 mg/L水杨酸处理的菊花,其耐热温度分别为35.0 、35.8 ℃,分别比对照低17.6%、15.8%。
3讨论
高温胁迫会导致植物细胞膜脂过氧化,叶绿体结构遭到破坏,导致叶绿素含量降低[13]。研究表明,叶面喷施脱落酸、水杨酸、Ca2+、多效唑(PP333)、6-苄基腺嘌呤(6-BA)可以提高植物的耐热性,而关于外源水杨酸提高作物的耐热性又是研究的热点[14]。本试验通过不同浓度的水杨酸对菊花进行处理,发现菊花的耐热性有差异,其中200 mg/L水杨酸处理的耐热性较强,表明外源水杨酸能诱导菊花产生耐热性。这可能是因为外源水杨酸降低了高温胁迫下菊花的质膜氧化程度,细胞膜透性减小,高温下菊花叶片细胞膜的稳定性增加。这与水杨酸诱导银杏、高羊茅、玉米等植物的结论[6,15-16]一致。
对于水杨酸的施用,浓度高低会导致菊花耐热性的差异。低浓度水杨酸处理的菊花耐热性强,而高浓度水杨酸处理的菊花耐热性降低,同时也表现在膜透性、茎叶干物质含量及叶绿素含量降低上,这与Fariduddin等研究结论[17]较为一致。随温度升高,水杨酸处理的神马菊茎叶干物质含量和叶绿素含量均呈先升高后降低的趋势,这与杨岚等研究结论[14,18]较为吻合。短期高温胁迫下,温度的升高使菊花的光合作用加强,叶绿素含量上升;但持续的高温胁迫造成叶绿体结构遭到破坏,位于类囊体膜中的光合系统受到损害,从而导致叶绿素含量下降[19]。当温度过高,超过42 ℃时,神马菊干物质含量、叶绿素含量都呈下降趋势,而200 mg/L水杨酸处理的神马菊叶绿素含量和茎叶干物质含量仍维持在较高水平,这表明水杨酸虽然不能根本阻止高温胁迫而避免菊花受到伤害,但喷施适宜浓度水杨酸可使菊花对热胁迫的耐受时间延长。
参考文献:
[1]孙宪芝,郭俊娥,郑成淑.菊花的高温伤害及生长恢复研究[J]. 山东农业大学学报:自然科学版,2013,44(1):6-11.
[2]Halim V A,Vess A,Scheel D,et al. The role of salicylic acid and jasmonic acid in pathogen defence[J]. Plant Biology,2006,8(3):307-313.
[3]郝敬虹,易旸,尚庆茂,等. 干旱胁迫下外源水杨酸对黄瓜幼苗膜脂过氧化和光合特性的影响[J]. 应用生态学报,2012,23(3):717-723.
[4]尚庆茂,宋士清,张志刚,等. 水杨酸增强黄瓜幼苗耐盐性的生理机制[J]. 中国农业科学,2007,40(1):147-152.
[5]孙艳,徐伟君,范爱丽.高温强光下水杨酸对黄瓜叶片叶绿素荧光和叶黄素循环的影响[J]. 应用生态学报,2006,17(3):399-402.
[6]何亚丽,刘友良,陈权,等. 水杨酸和热锻炼诱导的高羊茅幼苗的耐热性与抗氧化的关系[J]. 植物生理与分子生物学学报,2002,28(2):89-95.
[7]李永红,魏玉香,谷茂.水杨酸预处理对鸡冠花幼苗热胁迫的生理效应[J]. 西北植物学报,2008,28(11):2257-2262.
[8]王利军,黄卫东,战吉成.水杨酸和高温锻炼与葡萄抗热性及抗氧化的关系[J]. 园艺学报,2003,30(4):452-454.
[9]陈秋明,尹慧,李晓艳,等. 高温胁迫下外源水杨酸对百合抗氧化系统的影响[J]. 中国农业大学学报,2008,13(2):44-48.
[10]张钢,刘民,任元新.春季白皮松实生苗的脱锻炼与再锻炼[J]. 植物生理学通讯,2005,41(6):761-763.
[11]李亚青,张钢,卻书鹏,等. 白皮松茎和针叶的电阻抗参数与抗寒性的相关性[J]. 林业科学,2008,44(4):28-34.
[12]张宪政.作物生理研究法[M]. 北京:农业出版社,1992:148-149.
[13]刘大林,张华,曹喜春,等. 夏季高温胁迫对紫花苜蓿光合生理机制的影响研究[J]. 草地学报,2014,22(3):657-660.
[14]杨岚,师帅,王红娟,等. 水杨酸对高温胁迫下铁皮石斛幼苗耐热性的影响[J]. 西北植物学报,2013,33(3):534-540.
[15]曹福亮,欧祖兰.水杨酸对银杏幼苗抗高温胁迫能力的影响[J]. 浙江林学院学报,2008,25(6):756-759.
[16]杜朝昆,李忠光,龚明.水杨酸诱导的玉米幼苗适应高温和低温胁迫的能力与抗氧化酶系统的关系[J]. 植物生理学通讯,2005,41(1):19-22.
[17]Fariduddin Q,Hayat S,Ahmad A. Salicylic acid influences net photosynthetic rate,carboxylation efficiency,nitrate reductase activity,and seed yield in Brassica juncea[J]. Photosynthetica,2003,41(2):281-284.
[18]周中亮,包满珠,王文恩.高温胁迫对6个高羊茅株系生理指标的影响[J]. 草业科学,2011,28(7):1284-1290.
[19]Havaux M. Temperature sensitivity of the photochemical function of photosynthesis in potato (Solanum tuberosum) and a cultivated andean hybrid (Solanum×juzepczukii)[J]. Journal of Plant Physiology,1995,146(1/2):47-53.
猜你喜欢
动漫星空(兴趣百科)(2020年12期)2020-12-12
中州建设(2017年23期)2017-04-12
铝加工(2017年1期)2017-03-07
中国塑料(2015年6期)2015-11-13
绿色科技(2015年6期)2015-08-05
质量与标准化(2015年11期)2015-07-21
福建林业(2015年2期)2015-04-09
