
力竭运动对不同状态白鼠生理生化指标研究
2015-08-02 00:57徐宝丰
湖北体育科技 2015年7期
徐宝丰,薛 亮
疲劳分为肌肉疲劳(或称体力疲劳)和精神疲劳(或称脑力疲劳)两种。肌肉疲劳是指过度紧张的肌肉局部出现酸痛现象,一般只涉及大脑皮层的局部区域;精神疲劳则与中枢神经活动有关,它是一种弥散的、不愿意再作任何活动和懒惰的感觉,意味着肌体迫切需要休息。病理状态包括身体的一些慢性疾病或亚健康状态如心肌炎,某些器质性心脏病,重感冒期间或受到短暂、急剧的异常刺激造成病理状态。本文用白鼠做实验,主要从两方面着手,1)对疲劳白鼠突然增加运动强度;2)对有心脏病变的白鼠突然增加运动强度。观察这些白鼠损伤的程度,机体内血红蛋白(HB),血液中肌酸激酶(ck)、反应ATP-CP磷酸原系统供能情况,在各个时间段的生理生化指标,并对这些指标进行统计分析,得出运动猝死是否和这些因素有显著的相关性,观察这些指标和心脏损伤方面的关联,推断这些白鼠心源性损伤的程度。
1 研究对象与方法
1.1 实验对象与分组
饲养雄性白鼠120只,将白鼠平均分成4组,A组为正常对照组30只,B组为长期缺乏运动组30只,C组为运动量加大至疲劳组30只,D组为精神疲劳组30只。先将B组30只白鼠进行小笼饲养,笼子很小,几乎没有转身的环境,尽量让白鼠保持安静和不运动约60天后进行试验。对A、C、D进行正常饲养1周后,对C组进行干扰试验,在临近实验前两周采取日夜干扰的方法使得白鼠不能正常休息,特别是接近实验的前3天加大干扰的力度造成白鼠精神疲劳模型。
1.2 实验方式
动物模型准备完成后进行递增负荷跑台训练实验,运用辛东[6]参照 Bedford制定的递增运动负荷 5级标准[8]。 第 1天采用第 1级负荷:坡度 50°,速度 6m/min,时间 10min,进行适应性训练2次;第2天直接进行第5级负荷:坡度50°,速度28m/min,运动至力竭。运动时使用声、光刺激或用毛刷刺激动物尾部,以保证运动强度。力竭标准为:在5级运动负荷中,动物未能坚持本级负荷,经常在刺激情况下滞留跑道上。行为特征:呼吸急深、幅度大、俯卧位、刺激无反应。运用5级负荷模拟大运动量造成猝死模型,对运动中死亡白鼠立即取心尖部以及室间隔适量组织,10%甲醛固定。并抽血实验完成生理生化实验数据的收集整理。
1.3 样品采集
递增负荷跑台训练结束立即取 A、B、C、D组白鼠各 10只,取一侧颈动静脉混合血 5ml,离心(3 000rpm,10min)分离血清,20ul全血进行HB的测定;35ul血清用于CK的测定。3h后取 A、B、C、D组白鼠另外10只取血,24h后对每组最后 10只白鼠取血,取一侧颈动静脉混合血5ml,离心(3000rpm,10min)分离血清,20ul全血进行HB的测定。
HB采用高铁氰化钾法测定,试剂盒由上海欣科生物制剂公司提供;CK、BU的测定采用酶学方法,试剂盒由上海复星长征医学科学有限公司提供。
1.4 数据处理
测试数据采用SPSS PASW statistics18.0软件进行统计学分析,各组比较采用方差分析,方差不齐时采用校正t检验。相关系数采用显著性检验,分别采用P<0.05和P<0.01区别显著性和非常显著性差别。
2 结果和分析
2.1 不同组别白鼠在力竭运动后血红蛋白 (HB)各指标(+S)测试结果
正常对照组的白鼠在一次性力竭运动后血红蛋白快速下降,与安静状态下的样本均数T检验(P=0.46<0.05)有显著性降低,运动3h候继续下降,直到24h后仍然不能恢复至安静状态水平。长期缺乏运动组的血红蛋白水平与正常对照组的白鼠HB的水平没有差别,无论运动疲劳组还是精神疲劳组运动前机体的HB水平比正常状态下的白鼠水平高,血红蛋白量呈代偿性增加。力竭运动后即刻缺乏运动组和疲劳组血红蛋白下降很快,与安静状态下的HB相比有非常显著性降低(P<0.01)。运动3h后,B、C、D三组白鼠的HB含量与运动即刻相比都有所上升。运动24h后持续上升,B、C组都没有超过运动前的水平,D组超过运动前水平,说明HB的完全恢复需要一天以上的时间。另外,B、C两组白鼠在运动后24h其HB含量与精神疲劳组相比有非常显著的差别(P<0.01)。
2.2 不同组别白鼠在力竭运动后血红蛋白 (CK)各指标(+S)测试结果
表1 不同组别白鼠在力竭运动后血红蛋白(HB)各指标(+S)测试结果

表1 不同组别白鼠在力竭运动后血红蛋白(HB)各指标(+S)测试结果
注:和安静对照组Rest相比★代表P<0.05有显著性差异,★★代表P<0.01非常显著性差别;和正常状态下白鼠相比▼代表P<0.05有显著性差异,▼▼代表P<0.01非常显著性差别;和运动疲劳组相比#代表P<0.05有显著性差异,##代表P<0.01非常显著性差别。
安静对照组R e s t(g.l-1) 运动后即刻(g.l-1)A正常状态对照组B长期缺乏运动组C运动疲劳组D精神疲劳组1 4 6.1 2±7.4 1 1 4 5.1 3±7.2 3 1 6 2.3±8.6 7 1 5 3.1±4.8 1运动后3 h(g.l-1) 运动后2 4 h(g.l-1)1 2 1.2 2±6.8 5★9 8.3 0±5.6★★▼1 1 7.1 7±8.5 5★★1 2 2.3±5.6 7★★1 1 9.3 3±7.2 5★1 1 9.5±4.8 5★★1 3 4.6 3±6.2 3 1 3 1.8 3±2.3 5★1 2 9.1 5±8.7 6 1 2 1.8 4±6.5 1★##1 3 6.5 7±5.7 7##1 5 9.3±4.7 7
表2 不同组别白鼠在力竭运动后肌酸激酶CK(u.l-1)各指标(+S)测试结果
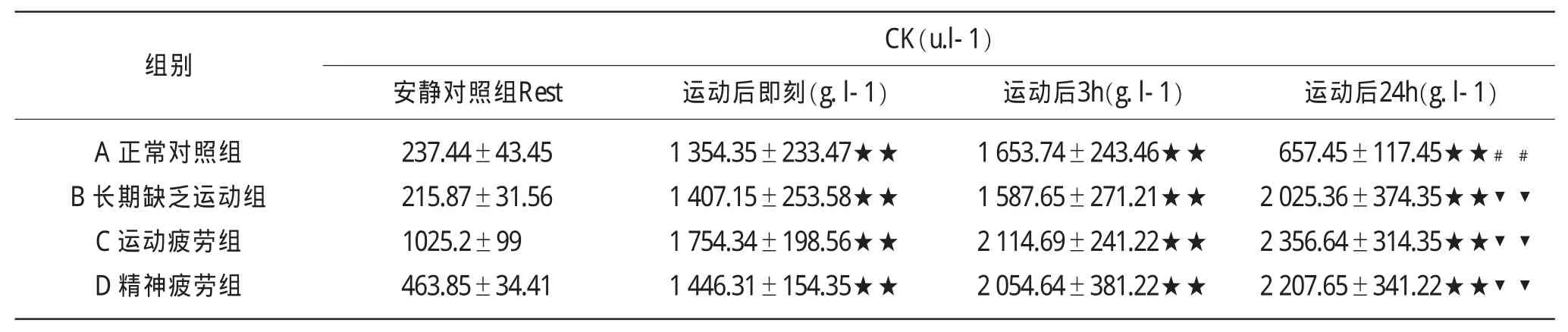
表2 不同组别白鼠在力竭运动后肌酸激酶CK(u.l-1)各指标(+S)测试结果
注:和安静对照组Rest相比★代表P<0.05有显著性差异,★★代表P<0.01非常显著性差别;和正常状态下白鼠相比▼代表P<0.05有显著性差异,▼▼代表P<0.01非常显著性差别;运动后24h与运动即刻组相比﹟代表P<0.05有显著性差异,﹟﹟代表P<0.01非常显著性差别。
组别C K(u.l-1)安静对照组R e s t 运动后即刻(g.l-1) 运动后3 h(g.l-1) 运动后2 4 h(g.l-1)A正常对照组B长期缺乏运动组C运动疲劳组D精神疲劳组2 3 7.4 4±4 3.4 5 2 1 5.8 7±3 1.5 6 1 0 2 5.2±9 9 4 6 3.8 5±3 4.4 1 1 3 5 4.3 5±2 3 3.4 7★★1 4 0 7.1 5±2 5 3.5 8★★1 7 5 4.3 4±1 9 8.5 6★★1 4 4 6.3 1±1 5 4.3 5★★1 6 5 3.7 4±2 4 3.4 6★★1 5 8 7.6 5±2 7 1.2 1★★2 1 1 4.6 9±2 4 1.2 2★★2 0 5 4.6 4±3 8 1.2 2★★6 5 7.4 5±1 1 7.4 5★★﹟﹟2 0 2 5.3 6±3 7 4.3 5★★▼▼2 3 5 6.6 4±3 1 4.3 5★★▼▼2 2 0 7.6 5±3 4 1.2 2★★▼▼
正常状态下的白鼠在运动后即刻,CK水平立即升高,和基础值相比有非常显著性升高(P<0.01)。运动后3h,其CK的水平继续升高。运动后24h后和基础值相比仍然有非常显著性升高(P<0.01),和运动后3h相比有非常显著性下降,但仍高于安静状态下的水平。长期不运动白鼠在安静状态下CK的水平比正常对照组白鼠略有降低,经过力竭运动后,其身体CK的水平非常显著性增高(P<0.01),超过正常组力竭运动后水平,运动后3h机体CK水平继续上升,运动24h后身体肌酸激酶的水平继续上升,和正常对照组相比有非常显著的差别(P<0.01)。运动疲劳组,由于前期大运动量的积累,在进行力竭运动前,身体处于疲劳状态,经过力竭运动后,白鼠身体CK水平进一步增加,此后的12h,CK水平继续呈增高趋势,白鼠机体呈病理性损伤,精神疲劳组CK发展趋势和运动疲劳组上升曲线相似。
3 结论和分析
3.1 力竭性运动下不同状态白鼠HB水平分析
血红蛋白可以反映人体的携氧能力,血红蛋白低会造成运动能力受到限制,运动后恢复减慢。及时检测血红蛋白的含量,不仅能够掌握自身机体状况,还能及时调整运动形式和强度,加强营养的补充,从而有效改善机能状况和提高运动能力。本文用白鼠做实验,1)对长期不运动白鼠增加运动强度至力竭;2)对运动疲劳和精神疲劳白鼠增加运动强度至力竭,白鼠机体内血红蛋白(HB),在各个时间段的生理生化指标(见图1、2)。

图1 不同时间段各种状态白鼠HB量的变化
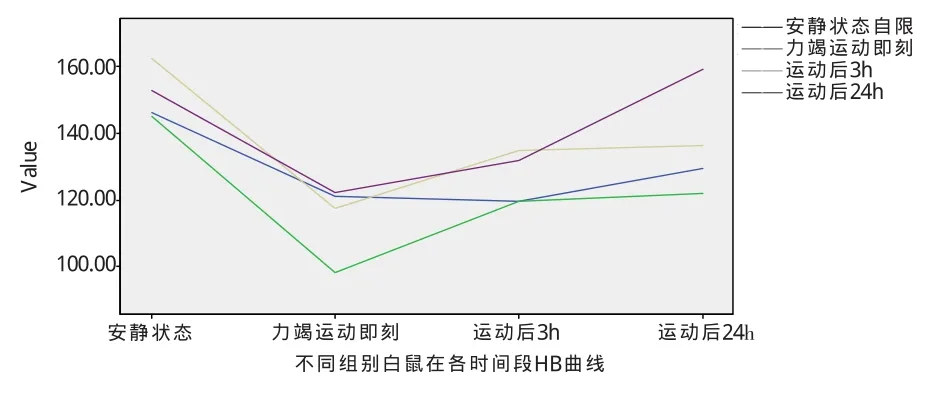
图2 不同组别白鼠在各时间段HB水平曲线
4种状态下,白鼠HB在运动后即刻全部有所下降,在运动后 3h,安静组没有回升,B、C、D组 HB都开始回升,运动1天后,精神疲劳组HB回升并超过机体原有水平,提示运动由于精神疲劳组HB蛋白的恢复。说明力竭性运动对不同组别白鼠的HB影响不大,在运动后即刻有所下降,然后随着时间的延长HB水平就逐渐恢复。
图2看出,A-D组的白鼠在一次性力竭运动后血红蛋白都快速下降,研究认为这是红细胞自由基的生成增加及红细胞氧化能力相对降低引起的红细胞破坏增加[6],Duca等认为马拉松运动员红细胞生成的改变是包括造血系统、溶血和氧化应激等多种因素相互作用所致[7]。Hb浓度降低是大运动量的早期反应。正常对照组的白鼠在一次性力竭运动后血红蛋白快速下降经过3h的休息恢复后,HB的水平又会回升,24h后继续恢复,这是机能改善和运动能力提高的表现。而B组白鼠HB的恢复水平最慢,1天后仍然低于其他任何组的恢复水平,提示长期不运动会降低机体对血红蛋白的恢复能力。而D组HB在运动1天后超过了安静状态下的水平,存在超量恢复现象,提示运动对精神疲劳有积极的作用,具体原因有待进一步研究。另外,B、C两组白鼠在运动后24h其HB含量与精神疲劳组相比有非常显著的差别,推断运动有助于精神疲劳组的血红蛋白的恢复。
长期缺乏运动组的血红蛋白水平与正常对照组的白鼠HB的水平在安静状态下没有差别,无论运动疲劳组还是精神疲劳组运动前机体的HB水平比正常状态下的白鼠水平高,血红蛋白量呈代偿性增加[8]。力竭运动后即刻缺乏运动组和疲劳组血红蛋白下降很快,与安静状态下的HB相比有非常显著性降低。肖建原、万发达等对大鼠进行不同负荷的训练后发现,大负荷训练后红细胞计数显著减少,红细胞破坏增多[9-10]。运动3h后,B、C、D三组白鼠的HB含量与运动即刻相比都有所上升。运动24h后持续上升,B、C都没有超过运动前的水平,说明HB的完全恢复需要一天以上的时间。
3.2 力竭性运动后不同状态的白鼠CK水平分析
肌酸激酶(Creatine Kinase,CK)通常存在于动物的心肌、骨骼肌、平滑肌、以及脑等组织的细胞浆和线粒体中,是一个与细胞内能量运转、肌肉收缩、ATP再生有直接关系的重要激酶,它可逆地催化肌酸与ATP之间的转磷酰基反应。在各种病变包括肌肉萎缩和心肌梗塞发生时,人的血清中肌酸激酶水平迅速提高,急性心梗时,酶活力上升程度与疾病严重程度相一致。因此,在心肌梗塞的诊断中测定肌酸激酶的活性比做心电图更为可靠。另外,多发性肌炎、皮肌炎、心肌炎等也可引起肌酸激酶CK升高,肌酸激酶因其具有重要的生理功能和临床应用价值已引起人们广泛的重视和深入的研究。所以,对机体运动后肌酸激酶水平的研究,对预防猝死的研究有很大的指导意义。白鼠机体内血液中肌酸激酶(ck)在各个时间段的生理生化指标(见图 3、4)。

图3 不同组别白鼠在不同时间段CK量的变化

图4 不同组别白鼠CK随时间变化曲线图
正常白鼠无论在那种状态下身体CK含量都很低,与B、C、D组白鼠相比其柱状面积明显矮短。C运动疲劳组一开始身体CK量就超过其他3组,在运动后身体CK水平继续增高,到运动后24h达到最大量,提示存在着运动疲劳现象。无论是缺乏运动组还是运动疲劳和精神疲劳组,其CK上升很快,在运动24h后,血清中肌酸激酶水平迅速提高,酶活力上升程度与疾病严重程度相一致。CK为细胞内酶,正常时在血清中的活性很低,当各种原因引起细胞膜通透性增加或细胞坏死时才会大量释入血液,使血清酶活性升高。由于运动引起氧自由基增加,造成细胞膜通透性增加或破损,细胞内酶向血液“漏出”增多,可能是导致血清酶活性升高的原因之一[11]。运动性疲劳是中枢神经系统出现的保护性抑制反应,受诸多因素的影响。健康人力竭运动后引起的血清酶活性改变应属正常的生理性变化,运动疲劳造成的细胞膜通透性增高或损伤是可逆的,在短期内即可恢复[12]。
除了A正常对照组,B、C、D组白鼠CK水平随着时间的延长,在1天内持续升高,在运动后24h升至最高。A组在最初也是继续升高,到3h后逐渐有所下降,到24h后已经非常显著性降低,但是仍然高于安静状态下水平的近3倍。说明疲劳和缺乏运动都会导致机体CK水平上升,且运动3h后持续上升,这种状态会持续到一天以后。Tiidus等人发现在大强度运动后,血清CK的活性一般在运动后5-6h开始上升,8-24h达到最大值,48h后开始下降。这与本人的研究结果相似,但后期的恢复趋势有待进一步研究[13]。由于方法技术等原因,更长时间的恢复没有进一步检测,可以推断如果长时间不能恢复[14],身体会出现病理性变化。
实验结果显示,长期缺乏运动组白鼠相对正常对照组的白鼠而言,其CK水平略有降低,而运动疲劳组由于其疲劳积累,身体CK水平明显升高,由精神原因引起疲劳组白鼠的CK也同样升高,提示不管什么性质的疲劳都容易引起身体肌酸激酶水平的升高。
医学上证明心肌梗死时肌酸激酶在起病6h内升高,24h达高峰,3-4日内恢复正常。证明健康状况下运动的白鼠在1天后机体的CK水平应有大幅度下降,而病理性的损伤导致CK的增高应在1天后继续升高,这应该是对运动机体是否是病理性进行判断的一个标志。同时也说明机体在病理状态或者疲劳状态下CK的恢复会比较慢。王福文等在研究中指出,过度运动力竭性运动可以诱发心肌损伤,且在运动后恢复期会出现延迟性心肌损伤。陈扬也在研究中发现,训练程度高的运动员比训练程度低的运动员血清CK活性恢复快[15],CK恢复慢的选手,专项素质和技术难以巩固和提高[16]。说明CK也与个体身体素质和个人能力相关。
长期不运动白鼠在安静状态下CK的水平比正常对照组白鼠略有降低,经过力竭运动后,其身体CK的水平非常显著性增高,(P<0.01)超过正常组力竭运动后水平,运动后3h机体CK水平继续上升,运动24h后身体肌酸激酶的水平继续上升,和正常对照组相比有非常显著的差别(P<0.01),说明长期不运动的人其机体对CK的反应非常敏感,且运动后CK升高很快,至少一天以后机体才完全恢复需要一天以上的时间,而正常对照组在运动24h后已能够恢复一半以上。
运动疲劳组,由于前期大运动量的积累,在进行力竭运动前,身体处于疲劳状态,经过力竭运动后,白鼠身体CK水平进一步增加,此后的12h,CK水平继续呈增高趋势,白鼠机体呈病理性损伤,精神疲劳组CK发展趋势和运动疲劳组上升曲线相似。
[1]马继政.运动诱发心肌病猝死机制的初步研究[D].南京:南京医科大学,2007.
[2]马继政,孙 飙,吕远远.运动性猝死预防及筛查策略[J].中国体育科技,2008(5).
[3]许思毛,刘涛波,苏全生.大负荷运动对大鼠心肌细胞膜纳钾泵、钙泵与肌浆网钙泵活性的影响[J].体育科学,2010(12).
[4]Laugens RP.Effect of acute and exhaustive exercise upon the fine structure of heart mitochondria[J].Experientia,1966(22).
[5]周 强.长袍运动至疲劳后血清酶活性变化[J].成都体育学院学报,2001(27).
[6]林 文,徐国琴,翁锡全.递增负荷运动引起运动性血红蛋白底下的特点及机制[J].北京体育大学学报,2011(5).
[7]Duca L,Da Ponte A,Cozzi M,etal.Changes in erythropoiesis,ironmet abolism and oxidative stress after half-marathon[J].Inter Emerg Med,2006(1).
[8]孟宪法.促红细胞生成素生成与调节机制的研究进展[J].高原医学志,1992(4).
[9]肖建原,赵 歌,郭建荣,等.不同负荷运动训练对大鼠红细胞膜的影响-氧化、抗氧化及膜流动性的变化[J].北京体育大学学报,2003(4).
[10]万发达,彭峰林,邓树勋.递增负荷运动训练对大鼠红细胞及血红蛋白的影响[J].军事体育进修学院学报,2008(4).
[11]张宏磊,王凤阳,张立立.递增负荷力竭运动对血清CKCK-MB活性的影响[J].沈阳体育学院学报,2004(3).
[12]周君来,王 建.FDP对运动性疲劳大鼠心肌与血清中酶活性及自由基代谢的影响[J].北京体育大学学报,2005(8).
[13]Tiidus P.M,Lanuzzo C.D.Effects of intensity and duration of muscular exercise on delayed soreness and serum enzyme activities [J].Med Sci Sports Exerc,1983(6).
[14]王福文,赵敬国,王 燕,等.力竭性运动后大鼠血清CK、CK-MB活性和心肌组织形态学的动力性改变 [J].Chin J Appl Physiol,2011(1).
[15]陈 扬.运动对血液中肌酸激酶乳酸脱氢酶琥珀脱氢酶的影响[J].解放军体育学院学报,2000(1).
[16]归予恒,陈文斌,曹文元.运用血清肌酸激酶值监控举重运动员的训练负荷承受力[J].中国运动医学杂志,2004(2).
猜你喜欢
冰雪运动(2021年1期)2021-07-28
安徽医专学报(2020年3期)2020-12-25
数学大王·低年级(2020年3期)2020-03-12
中成药(2018年3期)2018-05-07
岁月(2017年7期)2017-07-18
中国老年学杂志(2017年10期)2017-06-15
莫愁(2016年30期)2016-11-09
中国运动医学杂志(2016年3期)2016-07-10
中国当代医药(2015年21期)2015-03-01
当代体育科技(2015年8期)2015-02-27
