
长白山野生笃斯越橘根系内生菌资源调查
2015-07-18 18:40刘凤红程显好顾亮等
山东农业科学 2015年1期
刘凤红 程显好 顾亮等


摘要:为探讨长白山野生笃斯越橘菌根的形态结构和内生真菌的分布特征及种类,通过压片法对野生笃斯越橘菌根形态和内生真菌的分布进行了研究,并采用根段组织直接培养分离法和根段组织研磨培养分离法对根系内生真菌进行了分离培养,结合真菌形态和分子生物学方法对分离菌株进行了鉴定和多样性分析。结果表明,野生笃斯越橘菌根侵染率高达75%;对菌丝的形态观察得出,至少3类不同形态的内生真菌参与菌根的形成,粗菌丝形成致密的菌丝团,细菌丝形成疏松的菌丝团,疏松的粗菌丝贯穿细胞。两种分离方法共分离得到175株内生真菌,其中150株经形态学和分子综合鉴定为5属8种,其中Phialocephala spp.是优势属,占菌株总数的68.57%。根段组织研磨培养分离法较根段组织直接培养分离法获得的内生真菌多,细菌根较粗菌根多。研究获得的大量内生真菌资源为进一步生物防治菌株的筛选、开发与利用奠定了基础。
关键词:野生笃斯越橘; 菌根; 内生真菌; 形态特征; 分子鉴定
中图分类号:S476.1 文献标识号:A 文章编号:1001-4942(2015)01-0041-06
Abstract The mycorrhizal morphology and distribution of endophytic fungi in wild Vaccinium uliginosum Linn. in Changbai Mountain were studied through tabletting method. The endophytic fungi were isolated through root segment culturing method and grinding method, and were identified and analyzed based on fungal morphology and molecular biology technique. The results showed that the mycorrhizal infection rate was as high as 75%. According to the morphology of hyphae, the wild Vaccinium uliginosum Linn. contained at least three kinds of endophytic fungi, among which, thick hyphae formed dense hypha body, loose thick hyphae ran through the cell and thin hyphae formed loose hypha body. A total of 175 strains of endophytic fungi were isolated. Through morphology and molecular identification, 150 strains were divided into 8 species in 5 genera, among which, Phialocephala spp. was the dominant genera with the frequency of 68.57%. The amount of endophytic fungi isolated by root segment grinding method was larger than that of root segment culturing method, and that isolated from thin mycorrhiza was larger than that from thick mycorrhiza. The isolation of plentiful endophytic fungi laid foundations for further screening, development and utilization of bio-control strains.
Key words Vaccinium uliginosum Linn.; Mycorrhiza; Endophytic fungi; Morphological characteristics; Molecular identification
笃斯越橘(Vaccinium uliginosum Linn.)属杜鹃花科(Ericaceae)越橘属(Vaccinium)多年生落叶灌木,没有根毛,为浅根系植物,果实为蓝紫色小浆果[1,2]。研究表明,越橘果实中花青素含量是所有水果与蔬菜之最[3],花青素具有抗氧化、抗癌和改善视力等巨大的保健价值[4],此外,越橘还含有尼克酸、黄酮等特殊成分,因此常被誉为“浆果之王”[3]。由于具有丰富的营养保健价值,越橘在我国乃至世界的栽培规模不断扩大,已成为最具发展潜力的果树树种之一[5]。目前,我国栽培的越橘品种均引自国外,国内丰富的野生越橘资源未能得到有效的利用。东北的笃斯越橘作为我国重要且极具经济价值的野生越橘资源之一[6],其改良和驯化均未取得理想的成果,越橘菌根共生学方面的基础研究将为野生越橘资源改良和驯化提供必要的理论和实践支撑。
在自然条件下,越橘根系与菌根真菌共生形成杜鹃类菌根(Ericoid Mycorrhizal Fungi,EMF),又称欧石楠类菌根[1]。杜鹃类菌根真菌在缓解根系对水分及营养吸收能力低、植物对逆境因子抗性能力偏弱等问题上具有极其重要的作用[7]。在人工栽培条件下越橘根系和菌根真菌的共生需要较长的时间,且侵染率极低,有研究表明人工接种有效的菌根真菌将会显著促进越橘的生长发育[8]。因此根系真菌的分离纯化这一工作将有望解决野生笃斯越橘的移栽、驯化与改良等问题。endprint
本研究对长白山地区野生笃斯越橘菌根的形态结构和分布特征进行初步观察,并对其根系内生真菌进行了分离与鉴定,为笃斯越橘菌根生态学研究及内生真菌资源合理开发和利用提供理论依据。
1 材料与方法
1.1 材料与试剂
1.1.1 试验材料 试验材料为长白山野生笃斯越橘菌根。长白山自然保护区海拔1 300 m以下的笃斯越橘部分生长在泥炭藓沼泽草甸子里,部分生长在苔藓层较厚的落叶松林下。调查样地中心点位于N 43°08′27″、E 127°02′16″,距圆池入口西侧约9.5 km、边防公路北约1.8 km处。取样时轻轻去掉根系表面附着的苔藓枯死残留物,将健康的根连同泥土一并装入无菌采样袋,带回实验室立即进行处理。
1.1.2 培养基 内生真菌的分离与纯化使用MEA培养基,保藏使用PDA培养基。
MEA培养基:麦芽浸膏3%、大豆蛋白胨0.3%、葡萄糖2.0%、琼脂1.5%、蒸馏水定容至所配体积。
PDA培养基:马铃薯20.0%(煮汁过滤)、葡萄糖2.0%、琼脂1.5%、蒸馏水定容至所配体积。
MEA培养基和PDA培养基在灭菌之前均将pH调至5.7,121℃高压蒸汽灭菌30 min,灭菌后待培养基温度降至60℃左右,向培养基中加入青霉素(50 μg/mL)和链霉素(100 μg/mL)后使用。
1.2 试验方法
1.2.1 样品预处理 挑取新鲜、健康的根,将根上的苔藓、土壤清洗干净,根据不同的试验选择不同粗细的根。
1.2.2 菌根染色、形态观察 将选取的根切成1 cm长小段,用软化酸处理后于FAA固定液中固定。采用以下方法染色观察[9]:(1)组织透明:将根放入装有10% KOH溶液的试管中,并将试管置于80℃恒温水浴锅中水浴30 min,对根样进行透明处理;(2)漂洗:倒掉KOH溶液,用蒸馏水冲洗3次;(3)酸化:在室温条件下用5%双氧水浸泡15 min,倒出双氧水溶液,再用5%盐酸浸泡10 min, 倒掉盐酸溶液;(4)染色:向试管中加入适量台盼蓝溶液,同时将试管置于80℃恒温水浴锅中染色2 h;(5)脱色:染色完成后,取出根样在乳酸中脱色10 min;(6)镜检:将脱色后的根放到滴有乳酸的载玻片上,盖上盖玻片进行光学显微观察。
用镊子挑取粗细均匀的根段,剪成约0.5 cm长小段,共100个根段,每5个根段整齐排列于滴有乳酸的干净载玻片上,盖上干净盖玻片,用光学显微镜观察,记录菌根的菌丝形态与分布及每个根段的侵染情况。
1.2.3 侵染率计算方法 按照根段频率标准法测定菌根侵染率[10],即根据每段根系存在菌根结构的长度占根段总长度的百分比(没有菌根结构的根段侵染率为0,整条根段都被侵染的为100%,只有一半长度被侵染的为50%),按0、10%、20%、……、90%、100%的侵染程度计算根段侵染率,并记录各种侵染率下的根段条数,然后按照下列公式计算该样品的侵染率:
侵染率(%)=[∑(0×根段数+10%×根段数+20%×根段数+……+90%×根段数+100%×根段数)]/总根段数
1.2.4 内生真菌的分离 将根样分成粗和细两种类型,并分别转移至两个培养皿中。
根段表面消毒:(1)细根:向装有细根的培养皿中倒入10%双氧水浸泡8 min后,用无菌镊子将根转移至无菌培养皿(每次加入溶液或洗根之前都重复此操作)中,无菌水洗3遍,再倒入6%的次氯酸钠消毒1.5 min,再用无菌水洗5遍,最后用无菌滤纸吸干,待用;(2)粗根:步骤同细根的消毒方法,仅改变消毒的时间,10%双氧水消毒10 min, 6%次氯酸钠消毒2 min。分别将消毒后的粗根和细根剪成0.3~0.5 cm长的根段。
内生真菌的培养:(1)根段直接分离培养:每个培养皿中放置4个根段,将根段用消毒过的镊子轻压进培养基,模拟内生真菌在土壤中的环境,有利于营养的吸收。将培养皿置于20℃培养箱中黑暗培养2~3周。粗根和细根分别重复25次。(2)组织研磨分离培养:向研钵中加入100个根段和少量无菌水,充分研磨后,用移液枪取适量研磨溶液至培养皿中,再向培养皿中倒入降温到60℃左右的培养基,轻微平摇培养皿使研磨溶液均匀分布于培养基中。粗根和细根分别重复25次。
内生真菌的纯化:在无菌条件下用接种针挑取菌丝至MEA培养基中,每个菌落在同一平板内点3个菌斑,20℃黑暗培养2周后看菌落形态、生长速度等是否一致,若不一致,继续进行二次纯化。
内生真菌的筛选:纯化完成后,将所有菌落都接种到PDA培养基上,一个菌落点1个菌斑,做3个重复,于20℃黑暗培养2周,若菌株都能生长,则进行形态鉴定;若出现部分不能生长的菌株,只有一个菌株时,该菌株就作为最终分离筛选的结果,而数量较多时,就只在MEA培养基上进行形态鉴定与筛选。对筛选出来的菌落特征进行描述与记录,主要包括菌落直径、颜色、质地等形态特征。记录完成后挑取菌落边缘菌丝,经处理后在光学显微镜下观察菌丝体、孢子、孢子梗等,进行形态鉴定。不能明确鉴定的菌株,进一步进行分子鉴定。
1.2.5 DNA的提取与序列测定 根系内生真菌培养及DNA提取参考庄彩云等[11]的方法;根系内生真菌菌株rDNA ITS区段的PCR扩增、产物纯化参考李潞滨等[12]的方法;PCR扩增产物直接送北京华大基因发展有限公司切胶纯化测序。
1.2.6 内生真菌的鉴定 真菌鉴定依据形态特征和ITS序列分析相结合的方法。菌种经纯化鉴定后,接入保藏培养基PDA斜面,于鲁东大学农学院微生物菌种保藏室4℃冰箱中保藏。
2 结果与分析
2.1 长白山野生笃斯越橘菌根着生状态
2.1.1 菌根侵染形态 根外菌丝:在所观察的部分野生笃斯越橘根系表面,根外的菌丝较为常见,比较常见的有两种类型,一种是细的菌丝,通常能密集包裹、缠绕在整个根系表面(图1A);另一种是较粗的有隔菌丝,比较稀疏的分布在根系表面(图1B、C)。endprint
内生真菌菌丝:在野生笃斯越橘根系内观察到了三类不同形态的内生真菌菌丝,在细胞间游走的菌丝(图1F、G)、细胞内菌丝团(图1D、E)和贯穿细胞的菌丝(图1H)。在细胞间隙游走的菌丝以粗菌丝为主,菌丝都有隔,但是形态有所不同,有的较为规则地沿着细胞间隙纵向生长(图1G),有的则不规则、弯曲地贴着细胞生长(图1F);细胞内菌丝团同根外菌丝一样,也有粗和细两种菌丝体,粗菌丝形成致密菌丝团(图1D),会明显撑大所侵染的细胞,在显微镜下菌丝膨大成泡囊状,细菌丝则形成疏松的菌丝团(图1E),不会明显影响细胞的形状;贯穿细胞壁的菌丝较少见。除了以上三类内生真菌菌丝外还观察到了根细胞内正在降解的菌丝(图1I)。
2.1.2 菌根侵染率 经观察与统计,长白山野生笃斯越橘菌根侵染率达到75%,且带有根尖的根段侵染率普遍较低。
2.2 内生真菌的分离
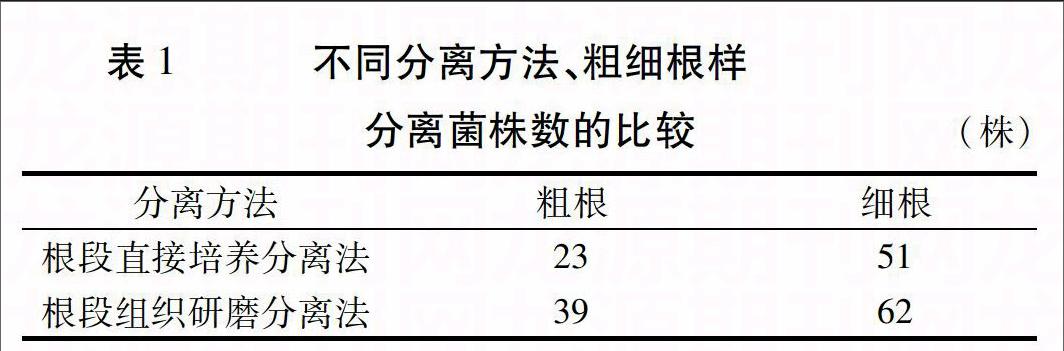
2.2.1 不同分离方法、粗细根样对内生真菌分离效果的影响 内生真菌分离试验结果(表1)显示,从粗根和细根分离的结果来看,粗根中分离得到62个菌株,细根分离得到113个菌株,细根分离到的菌株数明显高于粗根的菌株数。从分离方法上看,根段组织直接培养分离法得到74株菌株,根段组织研磨培养分离法获得101株菌株,根段组织研磨培养分离法优于根段组织直接培养分离法。
2.2.2 菌株形态特征 将分离到的175株内生真菌根据其形态特征差异分为12类,各类菌株形态描述如表2。
2.3 内生真菌分类
分离得到的175株内生真菌,其中150株经形态学和分子综合鉴定为5属8种(表3);另外25株无法命名到属。Phialocephala spp.是优势属,占菌株总数的68.57%。
3 结论与讨论
3.1 长白山野生笃斯越橘菌根着生状态
从显微观察结果来看,长白山野生笃斯越橘菌根的菌丝形态多样,根外菌丝与根内菌丝的形态具有一致性,同时根外菌丝从稠密程度上明显高于根内菌丝,这为人工接种内生真菌提高侵染率提供了理论依据[11]。在表皮细胞内观察到的粗菌丝形成致密的菌丝团,在一定程度上改变了细胞的形状,这种变化可以增加营养吸收的面积还是会对表皮细胞造成一定的危害,尚需要进一步试验求证。长白山野生笃斯越橘内生真菌的侵染率达到了75%,这对于分离多种多样的内生真菌提供了丰富的资源。有试验表明菌根真菌能侵染中柱[2],由于本试验未做越橘菌根的横切片,只对毛根处理后直接压片,未能见到侵染中柱的真菌,该方面还需要进一步研究。
3.2 内生真菌的分离
从本次试验结果来看,细根不论是从分离菌株数量上,还是种类上明显多于粗根,其中粗根分离的菌株菌丝的颜色主要以灰黑色和白色为主,而细根则以灰黑色为主。细根与粗根本质上是根龄不同,说明分离出的白色菌株侵染性比较持久或者是白色菌株专性侵染根龄较长的根系。从细根的显微镜观察中看到正在降解的菌丝可以推测,造成细根分离出的菌株多于粗根的原因可能是某些种类的内生真菌到了一定的时间就会降解,造成根龄较长的菌根分离出来菌株的数量和种类较少。
无论从分离出菌株的数量还是种类上,采用根段组织研磨分离法分离出来的菌株均比根段直接培养法分离出来的菌株多。造成数量的差异主要是因为,研磨相当于把细菌根分成很小的根段,或者说通过研磨能把在菌根内缠在一起的菌丝最大程度地分离开,使菌丝尽可能形成独立的菌株,避免覆盖,而剪成一定长度的根段,由于不同种类的内生菌之间存在竞争关系,相互之间容易覆盖,一般一个小根段只能分离出一株内生真菌,数量是有限的,所以研磨处理分离的菌株数量多;同样的原因,造成种类的差异,研磨能把各个种类的真菌分离开,减少不同真菌之间的竞争,使得长势慢的真菌与长势快的真菌各自分离,所以研磨得到的种类多。根段组织研磨分离法为分离更多的菌种资源提供了切实可行的方法。
3.3 菌株的鉴定
分离到的175株内生真菌,根据其形态差异共分为12类,其中150株经形态学和分子综合鉴定为5属8种,其中有3个种都为Phialocephala属,有文献指出,该属是菌根中常见的内生真菌[13],形态多样。另外,还有25株真菌无法鉴定到属,有可能代表新真菌。
参 考 文 献:
[1] 伏洪峰, 杨秀丽, 闫伟. 大兴安岭野生越橘菌根形态学研究[J].内蒙古农业大学学报, 2013, 34(2): 165-169.
[2]苗迎秋,王贺新,李根柱, 等. 长白山笃斯越橘菌根形态结构及内生菌的分布特征[J]. 东北农业大学学报,2013,44(1): 81-85.
[3]胡雅馨, 李京, 惠伯棣. 蓝莓果实中主要营养及花青素成分的研究[J]. 食品科学, 2006,27(10): 600-603.
[4]方仲相, 胡君艳, 江波, 等. 蓝莓研究进展[J]. 浙江农林大学学报, 2013,30(4): 599-606.
[5]肖军, 杨涛, 杨镇, 等. 蓝莓菌根菌的分离与回接试验[J]. 辽宁农业科学, 2012(5): 13-16.
[6]宗长玲,邓萌,宗成文,等. 笃斯越桔研究进展[J]. 北方园艺, 2011(12): 173-176.
[7]张春英,戴思兰. 杜鹃花类菌根研究进展[J].北京林业大学学报,2008,30(3):113-119.
[8]高丽霞, 李森, 莫爱琼, 等. 丛枝菌根真菌接种对兔眼蓝莓在华南地区生长的影响[J].生态环境学报,2012,21(8): 1413-1417.
[9]Koske R E, Gemma J N. A modified procedure for staining roots to detect VA mycorrhizas[J]. Mycological Research, 1989,92(4):486-505.
[10]朱毓霞, 宋杰, 肖文娟. 中江石泉单参丛枝菌根真菌鉴定[J]. 中药与临床, 2011(3): 17-20.
[11]庄彩云, 李潞滨, 胡陶, 等. 适用于rDNA ITS分析的兰属植物菌根真菌培养及DNA 提取方法[J]. 北京农学院学报, 2007,22(3): 4-6.
[12]李潞滨, 胡陶, 唐征, 等. 我国部分兰属植物菌根真菌rDNA ITS序列分析[J]. 林业科学, 2008(2): 160-164.
[13]Heather D, Sarah H, Randolph S. Distribution and molecular characterization of the root endophyte Phialocephala fortinii along an environmental gradient in the boreal forest of Alberta[J]. Mycological Research, 2000,104(10): 1213-1221.endprint
猜你喜欢
中国医药导报(2016年32期)2017-02-28
江苏农业科学(2016年8期)2017-02-15
东方教育(2016年9期)2017-01-17
中国医药导报(2016年30期)2016-12-28
法制与社会(2016年33期)2016-12-15
新闻世界(2016年11期)2016-12-10
戏剧之家(2016年4期)2016-03-25
