
云杉落针病几个生理生化指标的变化
2015-07-15 03:28:34杨春琳许秀兰徐明庆刘应高
湖南农业大学学报(自然科学版) 2015年2期
杨春琳,许秀兰,徐明庆,刘应高*
(1.四川农业大学林学院,四川 雅安 625014;2.攀枝花市东区林业局,四川 攀枝花 617067)
云杉属(Picea Dietr.)植物为高大常绿乔木,分布于北半球温带及亚热带山地,全球现约有40 种,中国约有20 种。云杉是中国西部海拔1 000~4 500 m地区的主要造林树种。随着云杉广泛栽种,由散斑壳菌(Lophodermium sp. )引起的落针病日趋严重,对林木健康生长已造成影响。笔者以四川川西云杉(P. balfouriana)为研究对象,测定不同海拔云杉针叶枯死脱落前的生理生化指标,以期探明云杉落针前期针叶生理变化,为有效防治云杉落针病提供依据。
1 材料与方法
1.1 供试云杉
选择四川省甘孜州泸定县马厂梁林场(位于四川西部,青藏高原东部边缘,东经101°46′ ~ 102°25′,北纬29°54′ ~30°10′)海拔高度2 500 ~2 800 m 的川西云杉林为研究对象。
1.2 方法
1.2.1 样地环境和云杉落针病害调查
采用分级计数法,试验地选择海拔高度2 590、2 690 和2 790 m 的3个样地,每样地选取发病普遍的林分标准地10 块(10 m×10 m)。按照云杉落针程度分为4 级,即:0 级代表未发病植株;Ⅰ级代表植株针叶变黄,少量脱落,针叶发病百分率区间为(0,30];Ⅱ级代表植株针叶多数变黄、变褐,针叶发病百分率区间为(30,65];Ⅲ级代表植株针叶大量枯死,几乎完全脱落,针叶发病百分率区间为(65,100]。于2013年3、5、7、10月对各样地环境状况、云杉落针发病植株数等进行记录,计算病情指数。
1.2.2 云杉针叶生化指标测定
对每海拔各病级云杉,随机采样(考虑到数据的合理性和科学性,每次均定点采样,即每水平定点选取2 ~3 株植株进行随机采样,采样地均在向阳面),同一海拔不同病级云杉采集1年生针叶约20g,编号后存于冰桶中,用于生理生化指标测定。采样与病害调查同时进行。
过氧化物酶(POD)与过氧化氢酶(CAT)活性测定参照文献[2]的方法;多酚氧化酶(PPO)活性测定参照阎鉴等[3]的方法;苯丙氨酸解氨酶(PAL)活性测定参照谢世勇[4]的方法;针叶叶绿素含量测定参照朱莲等[5]的方法;丙二醛(MDA)含量测定采用硫代巴比妥酸显色法[6]。重复测定3次,酶活以相对酶活性表示。
1.3 数据处理
数据采用SPSS 20.0和Excel 2007软件进行统计和分析。采用Duncan 法进行多重比较分析,同时运用Origin 8.5 进行绘图。为便于数据分析,依据试验地实际情况,将病原菌的一个侵染循环分为侵染前期(3—5月)、侵染期(5—9月)和侵染后期(9—11月)。
2 结果与分析
2.1 海拔高度对云杉落针病害的影响
样地川西云杉均为17 ~20 龄幼林,密度6.5×103株/hm2左右。幼龄林发病在林场范围内相当普遍,呈版块状分布,已严重影响到林场云杉正常生长发育和蓄材。表1 表明,海拔越高,云杉落针病病情指数就越高。经调查统计,在海拔2 790 m 处,云杉平均发病植株率为72.13%;海拔2 690 m 处为70.57%;海拔2 590 m 处为52.1%。在发病最严重的10月,海拔2 790 m 处云杉发病株率达到80%;海拔2 690 m 处达76.08%;海拔2 590 m 处仅53.27%。
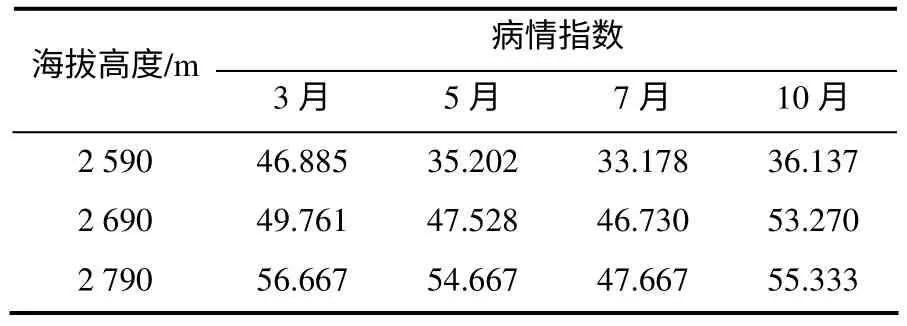
表1 不同海拔高度云杉的病情指数 Table 1 Disease indexes of P. balfouriana at different altitudes
2.2 川西云杉针叶生化指标变化
2.2.1 酶活性变化
由表2 可知,不同海拔,云杉落针病的病级越高,其针叶POD 活性也越高,且不同病级间差异显著;同一病级针叶中POD 活性,海拔2 650 m 处最高,海拔2 590 m 处最低。不同病级POD 活性变化呈先下降后上升或下降后上升再下降趋势。各月份间,针叶POD 活性差异不大。
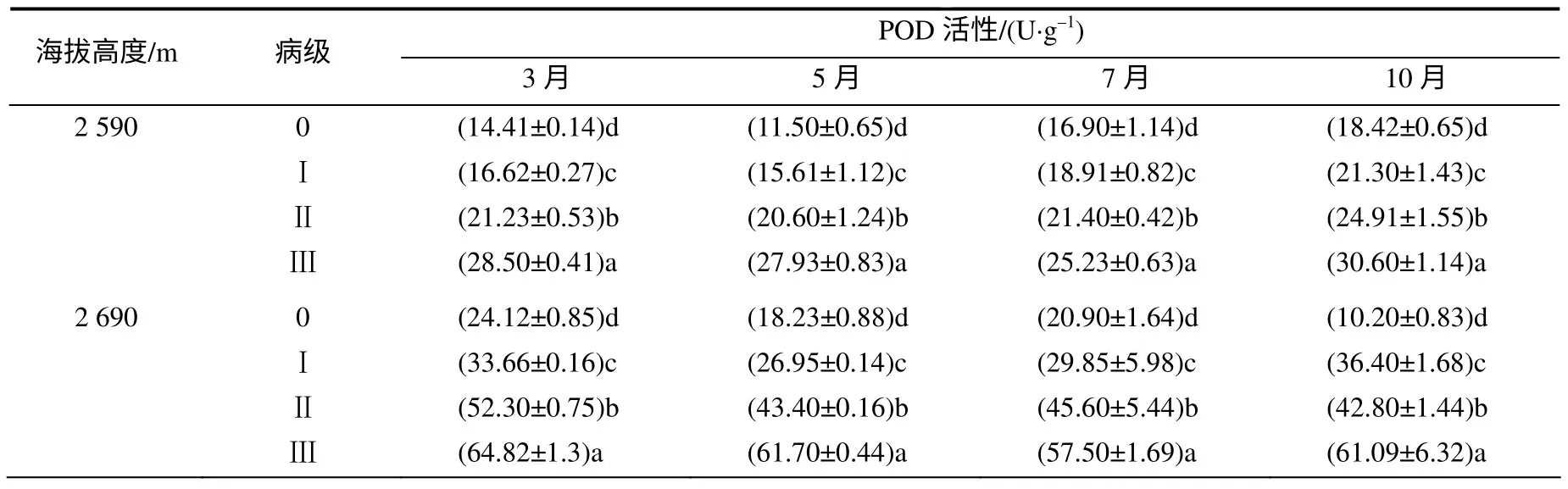
表2 各海拔高度不同病级云杉针叶的POD 活性 Table 2 POD activities of different disease level P. balfouriana at three altitudes

表2 (续)
表3 表明,病级为Ⅱ、Ⅲ级的云杉针叶的PPO活性高于病级为0、Ⅰ级的PPO 活性,两者间差异显著。同一病级,2 590 m 处PPO 活性低于其余2处。不同病级,PPO 活性变化大多数呈下降后再上升趋势。

表3 各海拔不同病级云杉针叶的PPO 活性 Table 3 PPO activities of different disease level P. balfouriana at three altitudes
由表4 知,随病级增加,各海拔高度云杉针叶的CAT 活性变化无明显规律。同一病级针叶的CAT活性,高海拔的略高于低海拔处。海拔2 590 m 处不同病级云杉CAT 活性变化呈下降再上升趋势;海拔2 690 和2 790 m 处病级为Ⅱ、Ⅲ级的云杉的CAT活性变化呈先上升后下降再上升趋势。
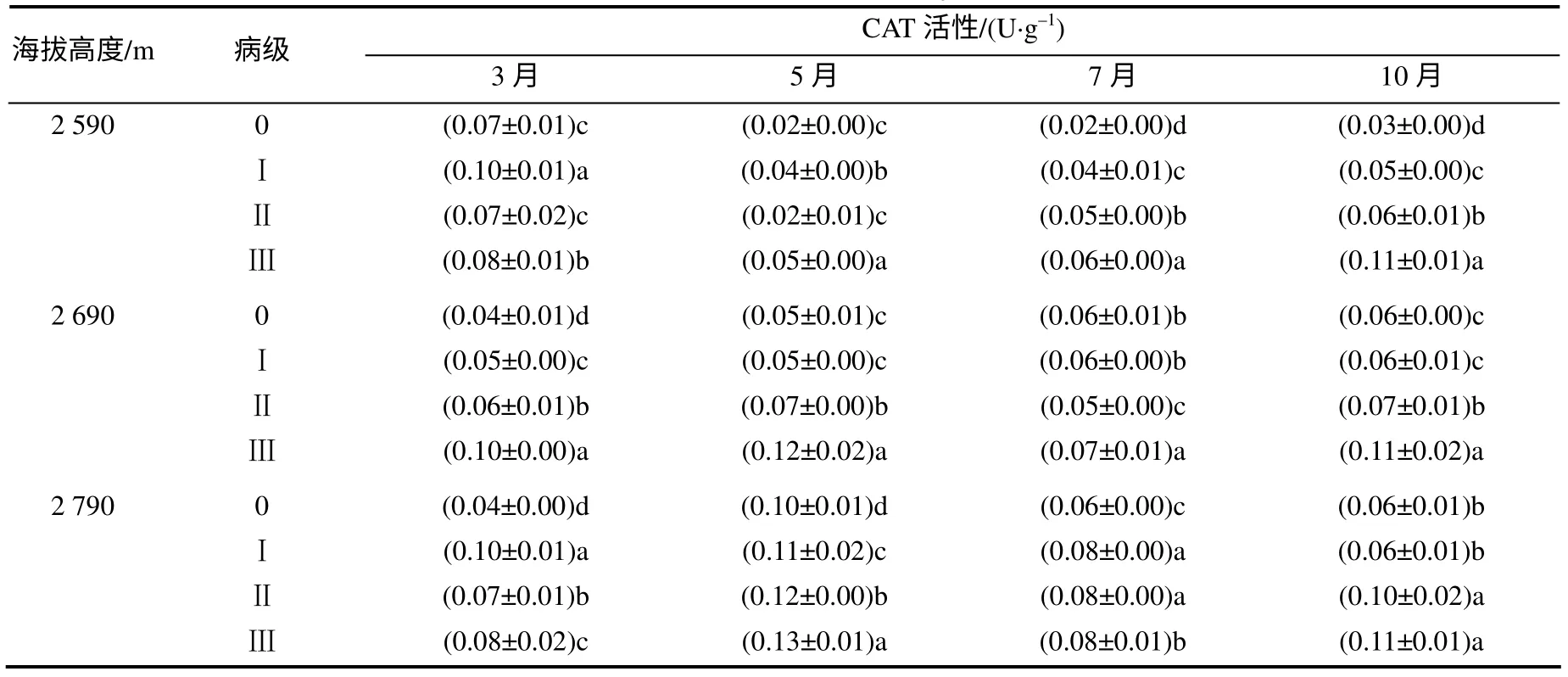
表4 各海拔不同病级云杉针叶的CAT 活性 Table 4 CAT activities of different disease level P. balfouriana at three altitudes
由表5 可知,病级越高,云杉针叶PAL 活性也越高(10月除外),且差异显著。同一病级云杉针叶,高海拔处PAL 活性略高于低海拔处。不同病级中,多数云杉针叶PAL 活性变化呈先下降再上升趋势,特别是海拔2 590 m 处,但后期PAL 活性较前期低。

表5 各海拔不同病级云杉针叶的PAL 活性 Table 5 PAL activities of different disease level P. balfouriana at three altitudes
2.2.2 叶绿素含量变化
表6 显示,叶绿素a(Chl a)较叶绿素b(Chl b)变化不规律,但病级为Ⅰ、Ⅱ、Ⅲ级的云杉针叶Chl a 含量最终均有不同程度增加,而健康云杉的Chl a含量变化较大,最终降低至较低水平,同时与其他病级间差异显著;Chl b 含量变化则与Chl a 相反,即病级越高,各海拔云杉针叶的Chl b 含量越低,但病级间差异不明显。另外,病级为Ⅰ、Ⅱ、Ⅲ级的云杉针叶的Chl b 含量最终有不同程度减少,而健康云杉的Chl a 含量最终有一定程度的增加。不同病级中,云杉针叶Chl b 含量变化多呈先上升后下降趋势。

表6 各海拔不同病级云杉针叶中的叶绿素a 和叶绿素b 含量 Table 6 Chl a (chl b) concentration of different disease level P. balfouriana at three altitudes
在各海拔处,病级越高,云杉针叶的总叶绿素含量在减少,总叶绿素含量呈先上升后下降趋势;在海拔2 590 m 处,病级为0、Ⅰ、Ⅱ的云杉,针叶总叶绿素含量在侵染初期还有下降过程,但最终总叶绿素含量均有不同程度的减少(表7)。

表7 各海拔不同病级云杉中总叶绿素含量 Table 7 Chl (a+b) concentration of different disease level P. balfouriana at three altitudes
2.2.3 MDA 含量变化
表8 中,健康云杉的MDA 含量基本都显著低于发病云杉。此外,MDA 含量在侵染后期多数比前期高。总体而言,同一病级云杉针叶MDA 含量,高海拔处的比低海拔处要高。
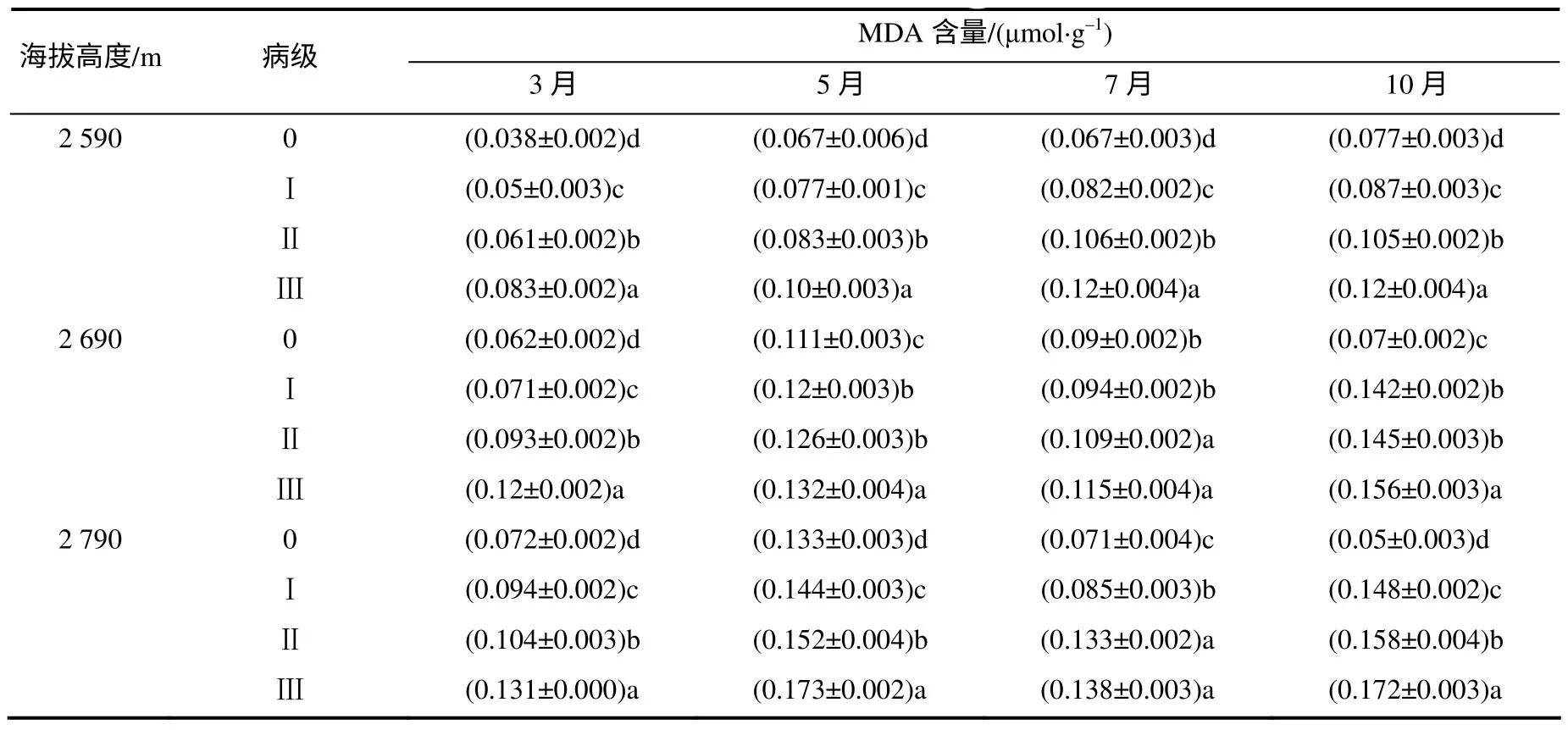
表8 各海拔不同病级云杉中的MDA 含量 Table 8 MDA concentration of different disease level P. balfouriana at three altitudes
3 结论与讨论
试验样地的云杉为幼龄林,散斑壳菌为其主要病原菌[7],该菌侵染周期为2年,共经历5个阶段[8]。1) 5—9月为子囊孢子释放期,子囊果成熟并破裂,同时释放子囊孢子经气孔或伤口等侵染针叶;2) 9—11月为症状发生期,针叶逐渐变为黄褐色或红褐色;3) 11 至次年3月为休眠期,病原菌以菌丝体等形式在针叶体内越冬,针叶症状无明显变化;4) 翌年3—5月为脱落期,前年被侵染针叶逐渐脱落,同时子囊果逐渐形成;5) 进入下一轮侵染循环,针叶中子囊孢子释放后,针叶逐渐被地表腐生物所分解。病原菌的持续侵染,可能导致寄主体内代谢紊乱,活性氧(ROS)含量增加[9]。本研究结果表明,随月份变化,云杉针叶POD 和CAT 酶活性均有所增加,两者在清除机体多余ROS 方面发挥了一定作用。有报道称,CAT 较POD 具有更高的H2O2催化效率[10],但病原菌引起机体过敏性反应(HR)时,CAT 活性将会受到抑制[11],这不利于ROS 的有效清除。另外,H2O2激增会反作用于病原菌[12],但过多H2O2会诱发植物细胞HR,最终导致细胞凋亡[13]。PPO 和PAL 活性前期均较高,对病原菌有一定抑制作用,但后期偏低,说明两者不能有效防御病原菌的侵染。随着海拔升高,云杉针叶的POD、PPO 和CAT活性以及MDA含量均有不同程度升高(PAL活性无显著差异),而且随病级增加有所增加,这说明海拔越高,云杉所承受的干扰力度越强(病级越高也是一样),体内会迅速产生并积累ROS,这种氧化猝发可能激活各防御酶基因,使其迅速转录并翻译酶蛋白,从而参与机体的抗病反应[14–15]。同时,这种氧化猝发也可能导致机体的蛋白质大量氧化,从而诱导机体自噬来缓解氧化胁迫[16]。虽然海拔越高,病级越高,各防御酶活性均有所升高,但MDA含量也在增加,这说明机体ROS 并没有因防御酶活性的增加而得到有效清除。可能是病原菌的持续侵染,使寄主细胞的DNA 遭受了损伤[17],最终使机体的ROS 含量增加,促使膜脂的进一步氧化。近来研究[18]发现,病菌持续侵染也会诱导寄主细胞自噬而发生过敏反应程序性细胞死亡(HR–PCD),这也能引起氧化猝发而使MDA 含量增加,而病原菌诱发HR–PCD 可能是导致云杉大量落针的主要原因。另外,总叶绿素含量随病级增加而减少,而且不同病级云杉的总叶绿素含量最终均有不同程度减少,也表明病原菌的持续侵染逐渐降低了针叶的光合作用。
[1] 杨春琳,许秀兰,黄晓丽,等.粗枝云杉幼林落针病的生理生化特性[J].东北林业大学学报,2014,42(9):148–152.
[2] 高俊凤.植物生理学试验指导[M].北京:高等教育出版社,2006:217–219.
[3] 阎鉴,麻彦明,张云龙,等.油松不同器官多酚氧化酶年变化规律的研究[J].山西农业大学学报,2001(2):135–137.
[4] 谢世勇,卢同,李本金,等.苯丙氨酸解氨酶、过氧化物酶与甘薯抗青枯病的关系[J].福建农业学报,2003,18(4):236–238.
[5] 朱莲,刘应高,李贤伟.老鹰茶白粉病罹病植株的生理生化研究[J].华北农学报,2007,22(增刊):266–270.
[6] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2003:165–260.
[7] 邱书志.云杉落针病发病规律的研究[J].西北林学院学报,1998,13(4):76–81.
[8] Jasinski J P P,Payette S.Holocene occurrence of Lophodermium piceae,a black spruce needle endophyte and possible paleoindicator of boreal forest health[J]. Quaternary Research,2007,67(1):50–56.
[9] 薛鑫,张芊,吴金霞.植物体内活性氧的研究及其在植物抗逆方面的应用[J].生物技术通报,2013(10):6–11.
[10] Kinnula V L,Lehtonen S,Sormunen R,et al. Overexpre- ssion of peroxiredoxins Ⅰ,Ⅱ,Ⅲ,Ⅴ,and in Ⅵmalignant mesothelioma[J].Journal of Pathology,2002,196(3):316–323.
[11] 刘延忠,王利民,李昶,等.植物细胞程序性死亡调控机制的研究进展[J].山东农业科学,2012,44(11):58–60.
[12] 郭泽建,李德葆.活性氧与植物抗病性[J].植物学报,2000,42(9):881–891.
[13] Levine A,Tenhaken R,Dixon R,et al.H2O2from the oxidative burst orchestrates the plant hypersensitive disease resistance response[J].Cell,1994,79(4):583.
[14] Choi D,Bostock R M,Avdiushkos,et al. Lipid-derived signals that discriminate wound and pathogen-responsive isoprenoid pathways in plants:Methyl jasmonate and the fungal elicitor arachiclonic acid induce different 3–hydroxy–3 methylglutary–l coenzyme A reductasegenes and antimicrobial isoprenoids in Solanum tuberosum L[J].Porc Natl Acad Sci USA,1994,91(6):23–29.
[15] 吴明江,于萍.植物过氧化物酶的生理作用[J].生物学杂志,1994(6):14–16.
[16] Xiong Y,Contento A L,Nguyen P Q,et al.Degradation of oxidized proteins by autophagy during oxidative stress in Arabidopsis[J].Plant Physiol,2007,143(1):291–299.
[17] 钟鸣,陈琢,刘宛,等.逆境胁迫下植物DNA 损伤和DNA 错配修复研究进展[J].生态学杂志,2012,31(9):2404–2411.
[18] Liu Y L,Schiff M,Czymmek K,et al.Autophagy regulates programmed cell death during the plant innate immune response[J].Cell,2005,121(4):567–577.
猜你喜欢
当代水产(2022年1期)2022-04-26 14:35:30
现代农村科技(2020年7期)2020-12-19 03:12:08
林业科学(2020年10期)2020-11-30 00:36:56
作文周刊·小学四年级版(2019年8期)2019-04-23 01:04:16
启迪与智慧·教育版(2018年12期)2018-02-26 12:24:06
山西林业科技(2017年4期)2018-01-23 05:45:05
甘肃林业(2016年2期)2016-11-07 08:56:12
西南农业学报(2016年5期)2016-05-17 05:42:33
西南农业学报(2016年6期)2016-04-16 05:12:51
国际木业(2016年6期)2016-02-28 10:27:03
