
青羊湖林场不同林分物种多样性与水源涵养能力的研究
2015-07-05 07:03:46梁文斌刘文胜张合平周晓光
西北植物学报 2015年9期
梁文斌,刘文胜,张合平,周晓光,潘 登
(中南林业科技大学 生命科学与技术学院,长沙410004)
国有林场是以植树造林和培育森林资源为主要任务的林业事业单位,在维持陆地生态系统平衡、保障人类基本生存条件和实现社会经济持续发展上起着重要的作用,建设好国有林场是充分发挥林场的经济、生态、社会效益的主要环节。提高森林的生态功能是国有林场的主要任务之一,它主要体现在维持生物多样性、涵养水源等方面。生物多样性保护是森林的重要功能之一,林分不同,其维持生物多样性的能力也不一样[1];同时,森林生物多样性又受经营模式的影响[2]。另一方面,森林地上部分水源涵养能力通常较低(小于15%),其林下土壤则是水源涵养的主体[3],而土壤的水源涵养能力受森林类型、经营模式等方面的影响[4]。因此,不同林分的生物多样性高低及林下土壤、枯落物的水源涵养能力是影响森林生态系统功能的关键因素。
水源地具有水源充足、土壤条件较好等特点,为植物的生长提供了良好的条件。但由于水源地植被承担着净化水质、保持水土等重要生态功能,植被状况对保障人们的正常生产、生活具有非常重要的作用。尽管有关不同林分物种多样性保护及水源涵养能力研究较多[4-6],但水源地附近的林分类型对土壤保水能力影响的研究仍较少[7,8],这制约了人们对水源地森林经营模式的认识。湖南省青羊湖国有林场水源充足,植被生长良好,为造林适宜地。南酸枣(Choerospondias axillaris)、栎(Quercus spp.)类、枫香(Liquidambar formosana)等阔叶树种及马尾松(Pinus massoniana)等针叶树种是当地的常见树种,它们常形成阔叶林、针叶林及针阔混交林等林型,在该区森林组成中具有重要作用。尤其是马尾松是当地的分布面积较广的树种,该树种具有耐干旱、耐瘠薄、适应能力强等特点,也是国有林场造林的主要树种之一[9],在造林中占重要地位[10]。
本研究选择位于水源地的湖南省青羊湖国有林场马尾松林、南酸枣林、马尾松-南酸枣混交林为研究对象,通过群落学调查、林下土壤及枯落物特性的分析,研究其林下物种多样性、水源涵养能力,探讨在水源地附近不同林分的水源涵养能力差异,其目的是探讨针叶林的物种多样性及水源涵养能力是否较阔叶林及混交林低,为水源地森林经营及水土保持提供科学依据。
1 研究区概况
研究区位于湖南省宁乡县西部的青羊湖国有林场,为雪峰山余脉自中低山向丘陵过渡地带,地理位置为E111°58′~112°05′,N28°08′~28°10′。气候类型属中亚热带向北亚热带过渡的大陆性季风湿润气候,气候温和,雨量充沛,年平均气温为16.2 ℃,最高气温高达40 ℃,最低气温低至-10.9 ℃;≥10℃的活动积温为4 900~5 000 ℃;年平均无霜期为273d,年平均日照为1 737.6h;年均降水量达1 438.8mm,相对湿度85%。成土母岩为板页岩。
由于林场所在区域的植被受人为活动影响大,多年的砍伐导致该区原生植被大部分被破坏,现有的植被类型主要为次生林和人工林,包括马尾松林、南酸枣林、马尾松-南酸枣混交林等。
马尾松林由人工飞播所形成,其林龄达20年,为该林场主要森林类型之一。为促进马尾松生长,该林场在2010年7月采用综合疏伐法+生长伐对部分林分进行间伐。未抚育的马尾松群落高达12 m,总盖度达90%以上,由乔木层、灌木层、草本层构成。乔木层郁闭度达0.8,主要树种有马尾松、杉木(Cunninghamia lanceolata)等;灌木层主要物种有檵木(Loropetalum chinensis)、山茶(Camellia japonica)、山莓(Rosa corchoriferus)等;草本层主要植 物 包 括 狗 脊 (Woodwardia japonica)、碗 蕨(Dennstaedtia scabra)、乌 蕨 (Stenoloma chusanum)等。抚育过的马尾松群落高度约12m,总盖度达90%,郁闭度达0.6,乔木层主要树种有马尾松、杉木等;灌木层盖度达70%,主要植物有楤木(Aralia chinensis)、大 青 (Clerodendrum cyrtophyllum)、海桐(Pittosporum glabratum)等;草本层主要植物有淡竹叶(Lophatherum gracile)、鳞毛蕨(Dyropteris championi)、狗脊等。南酸枣群落高度达15m,总盖度达90%以上。乔木层郁闭度达60%,主要树种有南酸枣、青冈、枫香、山茶、槲栎等;灌木层主要物种有山茶、栲、槲栎等;草本层主要植物包括狗脊蕨、碗蕨、乌蕨等。马尾松-南酸枣混交林(以下简称混交林)是飞播马尾松林的同时,人工植被与自然植被同时恢复而成,群落高度约14 m,总盖度达90%。乔木层主要树种有马尾松、南酸枣、枫香、青冈(Cyclobalanopsis glauca)等;灌木层盖度达70%,主要植物有楤木、大青、海桐等;草本层主要植物有淡竹叶、鳞毛蕨、狗脊等。
2 研究方法
2.1 样地设置与调查
采用样地调查法,在青羊湖林场选择生境因子(包括坡位、坡向、土壤特性及光、热、水分等)基本一致的南酸枣林(阔叶林)、马尾松-南酸枣混交林(针阔混交林)、未抚育马尾松林及抚育马尾松林(针叶林)进行群落调查。在乔木样地布设5 个20 m ×20m 的样方调查树种多样性,同时在每个样方中央及四角各设置1个5m ×5m 的灌木样方调查灌木植物多样性,在每个灌木样方内布设1个1m×1 m 的草本样方调查草本植物多样性。乔木样方主要调查和记录树高、胸径、冠幅等指标;灌木(包括高度≤2.0m 的乔木幼树)样方主要调查和记录各物种的名称、个体数、树高、胸径(地径)和物候等指标;草本样方主要调查并记录草本植物种名、个体数、高度、盖度等。
2.2 土壤持水量的测定
在4块样地的4个角和中心位置挖5个土壤剖面。土壤容重、最大持水量测定均采用环刀浸泡法分3层(0~10cm,10~20cm,20~30cm)进行[3]。
2.3 枯落物持水量测定
在每个样地的四个角和中心部位分别设置20 cm×20cm 的枯落物取样面积,按枯落物未分解和已分解分别取样,采用浸泡法计算枯落物的最大持水率及持水量。将采集的枯落物在自然状态下风干并称重,然后将风干样品装入沙布袋浸入水中,48h后取出样品,于100目筛中静置约5min,当无水滴滴下时称重,于75 ℃下烘干48h后称干重,根据公式(1)计算枯落物最大持水率和公式(2)计算枯落物持水量[3]。
(1)枯落物最大持水率(%)=(浸泡后的枯落物质量-烘干质量)/烘干质量×100%
(2)枯落物持水量(V)=L×C
式(2)中,V 代表枯落物持水量(t/hm2);L 代表枯落物蓄积量(t/hm2);C 代表枯落物最大持水率(%)。
2.4 数据处理
2.4.1 物种多样性的测度方法[11]
(3)Simpson多数性指数:D=1-∑Pi2
(4)Shannon-Wiener多数性指数:H′=-∑PilnPi
(5)Pielou均匀度指数:E=(-∑PilnPi)/lnS式(3)、式(4)和式(5)中Pi为某一层次第i个物种的个体数占所有物种的个体数的比值;式(5)中S为样地内某一层次所有物种的数量。
2.4.2 重要值(IV) 地上植被的物种重要值:IV=(相对密度+相对盖度+相对频度)/3。
2.4.3 DCA 分析 将物种重要值组成二维数据表作为DCA 分析的原始矩阵[12],采用软件Canonco 4.5 进 行DCA 分 析,运 用Sigmaplot 10.0 软 件作图。
2.4.4 统计分析 数据处理采用Microsoft Excel 2010。运用SPSS 12.0进行One-Way ANOVA 方差分析和LSD 差异性检验(α=0.05)。
3 结果与分析
3.1 物种组成及多样性
根据调查和统计,青羊湖林场各样地的维管束植物共有135种,隶属58科97属,其中南酸枣林物种相对较丰富(88种),混交林次之(76种),马尾松林较少(65种)。从表1可知,乔木层物种组成中,南酸枣林以南酸枣为优势种,其重要值达46.32,伴生种包括青冈、槲栎等,其重要值分别为17.21、10.78;混交林中马尾松和南酸枣均为优势树种,重要值均较高,分别为30.31 和23.58,杉木、青冈和枫香为伴生树种,重要值分别为13.59、12.07 和8.01;抚育马尾松林和未抚育马尾松林均以马尾松为绝对优势树种,其重要值均在98以上,杉木为伴生树种,重要值则很低,分别为1.39和1.11。
灌木层物种组成中,南酸枣林灌木层以山茶、栲(Castanopsis fargesii)、崖 豆 藤(Millettia dielsiana)为优势种,其重要值分别为13.32、8.07 和7.60;马尾松-南酸枣混交林灌木层山茶和大青的重要值较大,分别为12.17和11.29;未抚育马尾松林的灌木层优势植物为檵木和山莓,其重要值均超过10;抚育马尾松林灌木层的山莓和大青为优势植物,其重要值均在10以上。
草本层物种组成中,4种林分林下草本层优势种组成的差别不明显,均以芒萁(Dicranopteris linearis)、鳞毛蕨、淡竹叶和狗脊蕨为优势植物。
物种多样性研究表明(表2),不同林分的乔木层、灌木层和草本层的物种多样性均表现出明显差异。乔木层中马尾松林的物种多样性指数均低于0.20,而南酸枣林与混交林的多样性指数均较高,Shannon-Wiener多数性指数最高分别达1.390 与1.358;灌木层中,抚育马尾松林物种多样性较低,未抚育马尾松林具有最高的灌木物种多样性,南酸枣林与混交林居中,说明林分类型对物种多样性具有重要影响。

表1 不同林分类型主要物种组成及其重要值Table 1 Plant composition and important value of different forest types

表2 不同林分物种多样性Table 2 Species biodiversity of different forest types
3.2 群落结构相似性分析
DCA 二维排序图较好地反映了南酸枣林、马尾松-南酸枣混交林、未抚育马尾松林、抚育马尾松林4个群落结构间的差异(图1)。图1中第一轴解释植物群落结构32.6%差异,较好地区分混交林和南酸枣林,其得分明显高于其它两种方式(P<0.05);第二轴解释26.1%变异,显著区分未抚育马尾松林,而且在第二轴上得分明显低于其它3 种群落类型(P<0.05)。总体而言,南酸枣林和马尾松-南酸枣混交林相似性程度较高,说明群落类型对群落物种多样性有较大的影响。与第一轴显著正相关的物种有山茶、槲栎;与第一轴显著负相关的物种有狗尾草、酢浆草;与第二轴显著负相关的物种为南酸枣、山莓;与第二轴显著正相关的物种为马尾松、芒萁等。
3.3 水源涵养能力
枯落物的特性是反映林地涵养水源能力的重要因素之一,它体现在枯落物厚度、蓄积量、最大持水量等方面。从表3可知,不同林分的枯落物特性表现出明显的差异。从枯落物厚度来看,未抚育马尾松林未分解和已分解的枯落物厚度均最高,分别为15.5及8.2mm,南酸枣林未分解和已分解的枯落物厚度则最低,分别为6.0及6.7 mm,同时也发现马尾松林抚育后未分解枯落物厚度显著低于未抚育(P<0.05)。从枯落物蓄积量来看,未抚育马尾松林的林下枯落物蓄积量为9.9t·hm-2,南酸枣林和混交林的林下枯落物蓄积量分别为8.9和8.4t·hm-2,较未抚育马尾松林分别低10.1%和15.2%(P<0.05)。同样,马尾松林抚育后枯落物蓄积量也显著低于未抚育(P<0.05)。从最大持水率来看,马尾松林最大持水率高,高达293%,远高于南酸枣林(148%)和混交林(147%)。从最大持水量来看,未抚育马尾松林最大持水量达24.6t·hm-2,南酸枣林和混交林的林下枯落物最大持水量分别为16.2和13.6t·hm-2,比马尾松林低34.1%和44.7% (P<0.05)。从表3仍可以看出,马尾松林抚育后枯落物的最大持水量显著低于未抚育(P<0.05)。因此,南酸枣林和混交林的林下枯落物水源涵养能力低于两种针叶林。
林地土壤是水分贮蓄的主要场所,林地土壤水分蓄持能力与土壤厚度密切相关。土壤容重反映了土壤的松紧度和对地表水的蓄积能力,是影响森林土壤水源涵养功能的主要因素。容重小,土壤疏松,有利于涵养水源。从表4可知,不同林分类型土壤容重呈现出随着土壤加深,土壤容重增加的趋势,说明随深度加深,土壤储水量降低。不同林分类型土壤容重大小的顺序为:未抚育马尾松林>抚育马尾松林>混交林>南酸枣林。与未抚育马尾松林相比,南酸枣林和混交林0~10cm 土壤的容重分别降低14.7%、7.4%,10~20cm 土壤的容重分别降低14.1%、4.0%,20~30cm 土壤容重分别降低8.7%、4.9%。这说明南酸枣林树种在植被组成所占比例的提高,增加了土壤的疏松度和土壤的保水能力。
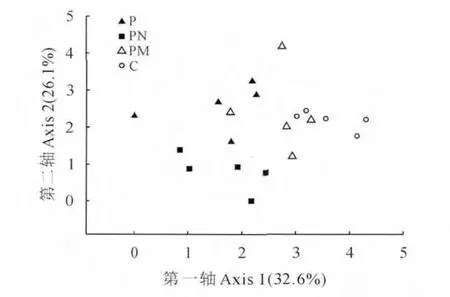
图1 不同群落的DCA 二维排序图PN.未抚育马尾松林;P.抚育马尾松林;PM.马尾松-南酸枣混交林;C.南酸枣林Fig.1 DCA biplot of plant community of 4forest types PN.P.massonianauntended forest;P.P.massonianatended forest;PM.P.massoniana-C.axillaris mixed forest;C.C.axillaris forest

表3 不同林分林下枯落物的厚度和储水特性Table 3 Water conservation capability of litter from different types of forests

表4 不同林分类型土壤的容重和储水量Table 4 Water conservation capability of soil from different forest types
由表4可知,不同林分类型土壤总储水量随着土壤深度的加深而减少。不同林分类型土壤总储水量大小顺序为:南酸枣林>混交林>抚育马尾松林>未抚育马尾松林。方差分析显示,南酸枣林土壤储水量为1 790.80t·hm-2,高于其他各林分(P<0.05),这与土壤容重的结果基本一致。混交林3层土壤总的储水量达1 614.78t·hm-2,抚育马尾松林 为1 580.06t·hm-2,未 抚 育 马 尾 松 林 为1 545.25t·hm-2,说明混交林及生态抚育提高了土壤总储水量。南酸枣林和混交林总储水量则较未抚育马尾松林高15.8%和4.5%。
4 讨 论
4.1 森林类型对物种多样性的影响
提高植物多样性是增强森林生态功能的重要环节,而林分类型对林下植物多样性具有显著影响,因而选择合适的林分类型在森林经营中具有重要意义。本研究结果表明,南酸枣林、混交林较未抚育马尾松林的物种多样性高,这与沈琪等[1]的研究结果一致。张克荣等[13]的研究结果也表明针阔混交林较针叶林物种多样性高,其原因是由于阔叶林下枯落物分解速度较针叶林快,对土壤具有改良作用[14],改善了林下植物的生境,利于许多植物定居,从而提高了植物多样性。这说明与阔叶林混交是提高森林物种多样性的重要途径。
另外,生态抚育对马尾松林林下物种多样性也有重要影响。本研究结果生态抚育降低了林下灌木的物种多样性,增加了草本植物的多样性。这与平亮等[15]及柏广新等[16]的研究结果一致;而与于立忠等[17]、段劼 等[18]、鲍 斌 等[19]的 研 究 结 果 不 完 全 一致,他们的研究发现抚育增加了林下灌木层与草本层物种多样性。这可能是因为抚育措施不同所导致,本研究主要是采用综合疏伐法+生长伐的方法进行抚育,以促进马尾松的生长,因而在短期内增加了林下层的光照,下层的草本植物获得光照而能够进行生长与发育,而灌木层经砍伐后在短期内尚未生长起来,因而提高了草本植物多样性、降低了灌木层的多样性。于立忠等[17]研究中的抚育措施主要是对乔木进行疏伐,未对灌木层及草本层进行干扰,其结果是林地下层获得光照,利用灌木及草本植物的生长,从而提高了灌木及草本植物的多样性。这说明抚育措施是影响植物多样性的重要方法。
4.2 森林类型对水源涵养能力的影响
提高森林水源涵养能力是森林经营的关键,而枯落物蓄积量和土壤蓄水量是衡量水源涵养能力的重要指标。本研究结果表明,南酸枣林、混交林枯落物蓄积量、最大持水量较两种马尾松林低,而总储水量则较两种马尾松林高。吴庆贵等[4]、康冰等[20]研究也显示阔叶林及针阔混交林的水源涵养能力较针叶林高。其原因可能是阔叶树种枯落物的木质素、纤维和次生代谢产物含量较低,分解速度较针叶植物快,对土壤具有改良作用[14],因此,降低了土壤的容重,能较好地改善土壤的通气性能,从而提高了土壤的持水量。这说明林分类型是影响水源涵养能力的重要因素。
生态抚育对马尾松林水源涵养能力也有重要影响。本研究结果表明抚育降低了马尾松林枯落物蓄积量及最大持水量,但提高了总储水量,增强了水源涵养能力。这与杨新兵等[8]对华北落叶松(Larix principis-rupprechtii)的研究结果一致,但与贾芳等[3]和贾忠奎等[21]的研究结果存在差异,即抚育不仅提高了林地枯落物蓄积量及最大持水量而且还提高了林地土壤总储水量。造成这种差异的原因可能与森林所处生境的水热及土壤条件有关。本研究与杨新兵等[8]研究中的样地所处位置均位于水源地附近,空气湿度较大,土壤肥沃、土壤水分含量较高,生态抚育使林内光照增强、气温增高,加速森林枯落物分解,枯落物蓄积量也将降低,从而利于改善土壤结构。而贾芳等 和贾忠奎等 研究中的样地则土壤瘠薄、保水能力差、土壤含水率较低,因而枯落物分解速度较慢。这说明抚育对枯落物及总储水量的影响与当地的生境条件密切相关。
[1] SHEN Q(沈 琪),ZHANG J(张 骏),ZHU J R(朱锦茹),et al.Changes of species composition and diversity in the restoration processes of ecological public-welfare forests in Zhejiang,East China[J].Acat Ecologica Sinica(生态学报)2005,25(9):2 131-2 138(in Chinese).
[2] LUO Y H(罗应华),SUN D J(孙冬婧),LIN J Y(林建勇),et al.Effect of close-to-nature management on the natural regeneration and species diversity in a masson pine plantation[J].Acta Ecologica Sinica(生态学报),2013,33(19):6 154-6 162(in Chinese).
[3] JIA F(贾 芳),JIA ZH K(贾忠奎),MA L Y(马履一),et al.Effects of Thinning on water conservation of young Pinus tabulaeformis plantation in Beijing mountain area[J].Journal of Soil and Water Conservation(水土保持学报),2009,23(6):235-239(in Chinese).
[4] WU Q G(吴庆贵),ZOU L J(邹利娟),WU F ZH(吴福忠),et al.Water Conservation capacity of five main forest in the hilly area of Fujiang river basin[J].Journal of Soil and Water Conservation(水土保持学报).2012,6(6):254-258(in Chinese).
[5] WANG Y(王 芸),OUYANG ZH Y(欧阳志云),ZHENG H(郑 华),et al.Effects of three forest restoration approaches on plant diversity in red soil region,southern China[J].Acta Ecologica Sinica(生态学报).2013,33(4):1 204-1 211(in Chinese).
[6] HAN L(韩 路),WANG H ZH(王海珍),LÜR H(吕瑞恒),et al.Water-holding characteristics of litter under different forest types in the upper reaches of Tarim river[J].Journal of Soil and Water Conservation(水土保持学报),2014,28(1):96-101(in Chinese).
[7] DUAN W B(段文标),LIU SH CH(刘少冲),CHEN L X(陈立新).Study on hydrological effect of water conservation forest in Lianhua Lake reservoir area[J].Journal of Soil and Water Conservation(水土保持学报),2005,19(5):26-30(in Chinese).
[8] YANG X B(杨新兵),ZHANG W(张 伟),ZHANG J H(张建华),et al.Effects of ecological tending on the litter and soil hydrological in young stand of north-China larch[J].Journal of Soil and Water Conservation(水土保持学报).2010,24(1):119-122(in Chinese).
[9] XIANG W H(项文化),TIAN D L(田大伦).Nutrient cycling in Pinus massonianastands of different age classes[J].Acta Phytoecologica Sinica(植物生态学报).2002,26(1):89-95(in Chinese).
[10] TANG X R(唐效蓉),LI W P(李午平),et al.Fertilizer effects on the plant growth and wood proportion of natural secondary forestry of Pinus massoniana[J].Journal of Central South Forestry University(中南林业科技大学学报).2006,26(1):12-18(in Chinese).
[11] MA K P(马克平),LIU Y M(刘玉明).Measurement of biotic community diversity[J].Chinese Biodiversity(生物多样性),1994,2(4):231-239(in Chinese)
[12] 张金屯.数量生态学[M].北京:科学出版社,2004.
[13] ZHANG K R(张克荣),LIU Y D(刘应迪),ZHU X W(朱晓文),et al.Community types and species diversity of Pinus massonianaforests of Yuelu Mountain,Changsha[J].Scientia Silvae Sinicae(林业科学).2011,47(4):86-94(in Chinese).
[14] LI Q(李 强),ZHOU D W(周道玮),CHEN X Y(陈笑莹).The accumulation,decomposition and ecological effects of above-ground litter in terrestrial ecosystem[J].Acta Ecologica Sinica(生态学报),2014,34(14):3 807-3 819(in Chinese).
[15] PING L(平 亮),XIE Z Q(谢宗强).Effects of introducing Eucalyptus on indigenous biodiversity[J].Chinese Journal of Applied Ecology(应用生态学报),2009,20(7):1 765-1 774(in Chinese).
[16] BAI G X(柏广新),ZHANG M J(张命军).Effect of thinning intensity on diversity of undergrowth plants in a hardwood forest in Changbai Mountains[J].Journal of Northeast Forestry University(东北林业大学学报),2011,39(12):27-29(in Chinese).
[17] YU L ZH(于立忠),ZHU J J(朱教君),KONG X W(孔祥文),et al.The effects of anthropogenic disturbances(thinning)on plant species diversity of Pinus koreansis plantations[J].Acta Ecologica Sinica(生态学报).2006,26(11):3 757-3 764(in Chinese).
[18] DUAN J(段 劼),MA L Y(马履一),JIA L M(贾黎明),et al.Effect of thinning on Platycladus orientalis plantation and the diversity of undergrowth vegetation[J].Acta Ecologica Sinica(生态学报),2010,30(6):1 431-1 441(in Chinese).
[19] BAO B(鲍 斌),DING G J(丁贵杰).Effects of thinning on growth and vegetation diversity of Pinus massoniana plantation[J].Journal of Central South University of Forestry &Technology(中南林业科技大学学报),2013,33(3):30-33(in Chinese).
[20] KANG B(康 冰),LIU SH R(刘世荣),et al.Soil physical and chemical characteristics under different vegetation restoration patterns in China south subtropical area[J].Chinese Journal of Applied Ecology(应用生态学报),2010,21(10):2 479-2 486(in Chinese).
[21] JIA ZH K(贾忠奎),WEN ZH Y(温志勇),JIA F(贾 芳),et al.Effect of thinning on water conservation of Platycladus orientalis plantation in Beijing mountain area[J].Journal of Soil and Water Conservation(水土保持学报),2012,26(1):62-66(in Chinese).
猜你喜欢
今日农业(2022年13期)2022-09-15 01:18:00
中成药(2021年5期)2021-07-21 08:38:46
今日中国·西班牙文版(2021年4期)2021-04-09 11:22:46
河北果树(2020年1期)2020-02-09 12:31:42
西部林业科学(2018年6期)2018-12-27 11:23:06
现代园艺(2018年2期)2018-03-15 08:01:03
广西林业科学(2016年1期)2016-03-20 05:33:02
广西林业科学(2016年4期)2016-03-16 05:44:58
科技视界(2014年25期)2014-12-25 06:40:04
理论观察(2014年2期)2014-04-29 07:15:45
