
施肥与竞争生长对入侵植物黄顶菊和本地植物光合特性的影响
2015-07-01 23:34:23张晓亚陈凤敏殷吉林张风娟
生物安全学报 2015年3期
张晓亚, 陈凤敏, 陈 雪, 闫 静, 殷吉林, 张风娟*
1河北大学生命科学学院,河北 保定 071002; 2昌黎县第三中学,河北 秦皇岛 066600;3河北师范大学汇华学院,河北 石家庄 050091
施肥与竞争生长对入侵植物黄顶菊和本地植物光合特性的影响
张晓亚1+, 陈凤敏2+, 陈 雪3, 闫 静1, 殷吉林1, 张风娟1*
1河北大学生命科学学院,河北 保定 071002;2昌黎县第三中学,河北 秦皇岛 066600;3河北师范大学汇华学院,河北 石家庄 050091
【背景】黄顶菊具有较强的生态适应性和竞争能力,可压制或排挤本地物种,形成单优势种群,进而导致当地生物多样性下降。【方法】选取黄顶菊入侵域的4种伴生植物益母草、苘麻、马唐、反枝苋为研究对象,采用LI-6400便携式光合仪测定入侵植物黄顶菊与这4种植物竞争生长过程中的光合特性,并研究施肥对不同植物与黄顶菊竞争生长过程中光合速率、蒸腾速率和胞间CO2浓度的影响。【结果】竞争生长对黄顶菊净光合速率的影响与本地植物种类密切相关,施肥促进了黄顶菊的净光合速率;竞争生长显著抑制了益母草、马唐和反枝苋的净光合速率,促进了苘麻的净光合速率,施肥增强了这4种植物的净光合速率。竞争生长对益母草和苘麻蒸腾速率的影响显著,而对其他2种植物的影响较小;与苘麻、益母草和反枝苋竞争生长过程中黄顶菊的蒸腾速率显著降低;施肥显著降低了单种处理中黄顶菊的蒸腾速率。与益母草、苘麻和反枝苋竞争生长过程中黄顶菊的胞间CO2浓度明显降低,施肥处理对黄顶菊的CO2利用率有显著抑制作用。【结论与意义】施肥影响黄顶菊的光合特性,与不同本地植物竞争生长对黄顶菊光合特性的影响不同。本研究结果将为进一步揭示黄顶菊的入侵机制及其综合治理提供理论依据。
入侵生物; 黄顶菊; 光合特性; 施肥; 竞争生长
黄顶菊Flaveriabidentis(L.) Kuntze.属菊科堆心菊族黄顶菊属,原产于热带地区的南美洲,主要分布于西印度群岛、墨西哥和美国的南部,后来传播到非洲的埃及、南非,欧洲的英国、法国,澳洲的澳大利亚和亚洲的日本等地(刘全儒,2005)。2001年先后在我国河北衡水、廊坊,天津南开等地大量涌现。外来生物本身的生物学特性和强生态适应性是其成功入侵的主要原因(王俊峰等,2004; 闫淑君等,2006; 杨晴等,2014; Mariaetal.,2002)。黄顶菊喜生于荒地,具有喜光特性,且其是较为少见的双子叶C4植物(Powell,1978)。植物生物量的迅速累积与其光合特性密切相关(Bouman & Sylliboy,2012; Chenetal.,2013; Xing & Wu,2015),因而推测其强的入侵性与其特殊的光合特性和土壤肥力有关,但相关内容鲜有系统报道。本研究探究不同肥力条件下不同植物与黄顶菊竞争生长过程中的光合特性,为进一步揭示黄顶菊的入侵机制提供理论依据。
1 材料与方法
1.1 试验材料
试验材料分别为黄顶菊与其入侵域常见的4种本地草本植物益母草Leonurusartemisia(Laur.) S.Y.Hu、苘麻AbutilontheophrastiMedicus、马唐Digitariasanguinalis(L.) Scop.、反枝苋AmaranthusretroflexusL.。
1.2 试验处理
田间试验于2012年5月中旬至9月中旬在河北科技师范学院昌黎校区农场进行(北纬39°70′、东经119°17′),土质为潮土,pH 7.6,电导率 531.0 μS,有机质含量1.70%,水解性氮108.35 μg·g-1,有效磷 26.21 μg·g-1,速效钾90.65 μg·g-1。本试验采用随机区组试验设计,试验处理分别为黄顶菊单独种植(以下简称单种)、马唐单种、益母草单种、苘麻单种、反枝苋单种、黄顶菊与马唐混合种植(以下简称混种)、黄顶菊与益母草混种、黄顶菊与苘麻混种、黄顶菊与反枝苋混种;每个处理分为施氮肥和不施氮肥2个水平,肥料为尿素,添加量为30 g·m-2,于播种前一次性加入。每个处理种植面积为3.0 m×1.0 m,种植密度为80 株·m-2。混种处理中,先将2种植物的种子混合后播种,待植物长出幼苗,再进行间苗处理;每个小区植物共32株,混种比例为1∶1,随机排列,小区之间为宽度0.2 m的隔离带,每个处理5次重复。试验期间,及时人工拔除其他杂草。
1.3 测量方法
2012年7月,选择全天晴朗无云的天气,利用LI-6400便携式光合作用测定系统测定试验样株的光合生理指标。具体方法:选择样株外围的阳性功能叶片(基本不受其他叶片遮挡),分别对其光合作用进行多次平行测量,即各选择样株4株,从每样株中随机选择2片健康成熟、无病虫害的叶片进行测量(侯智勇等,2009; 莫凌等,2009; 叶万辉和冯惠玲,2000; 朱慧和马瑞君,2009),记录净光合速率、蒸腾速率、胞间CO2浓度等数据。
1.4 数据分析
采用Duncan′s新复极差法分析每个变量对每种植物的主效应及其交互效应,所有统计分析均由SPSS 19.0专业版软件包完成。
2 结果与分析
2.1 净光合速率
2.1.1 入侵域本地植物的净光合速率 在单种处理中,施肥对益母草、马唐的净光合速率无显著影响(P>0.05);在混种处理中,竞争生长显著抑制了益母草和马唐的净光合速率,施肥显著增强了益母草和马唐的净光合速率(P<0.05)(图1)。
单种处理中,施肥增强了苘麻的净光合速率(P<0.05);混种处理中,竞争生长显著促进了苘麻的净光合速率,且施肥增强了苘麻的净光合速率(P<0.05)。单种处理中,施肥降低了反枝苋的净光合速率(P<0.05);混种处理中,竞争生长显著抑制了反枝苋的净光合速率,但施肥增强了反枝苋的净光合速率(P<0.05)(图1)。
2.1.2 黄顶菊的净光合速率 单种处理中,施肥显著增强了黄顶菊的净光合速率。与单种无肥处理相比,与益母草混种增强了黄顶菊的净光合速率,施肥对混种处理中黄顶菊的净光合速率没有影响;与马唐混种显著降低了黄顶菊的净光合速率,施肥增强了黄顶菊的净光合速率,但与单种处理差异不显著;与苘麻混种显著降低了黄顶菊的净光合速率,施肥增强了黄顶菊的净光合速率;施肥显著降低了与反枝苋混种的黄顶菊的净光合速率(图2)。

图1 本地植物净光合速率Fig.1 Net photosynthetic rate of the four examined native weeds 数据为平均值±标准差,不同小写字母表示植物在不同处理下差异显著(P<0.05)。 The datas are means±SD. Small letters indicate significant difference of plant in different treatments (P<0.05).
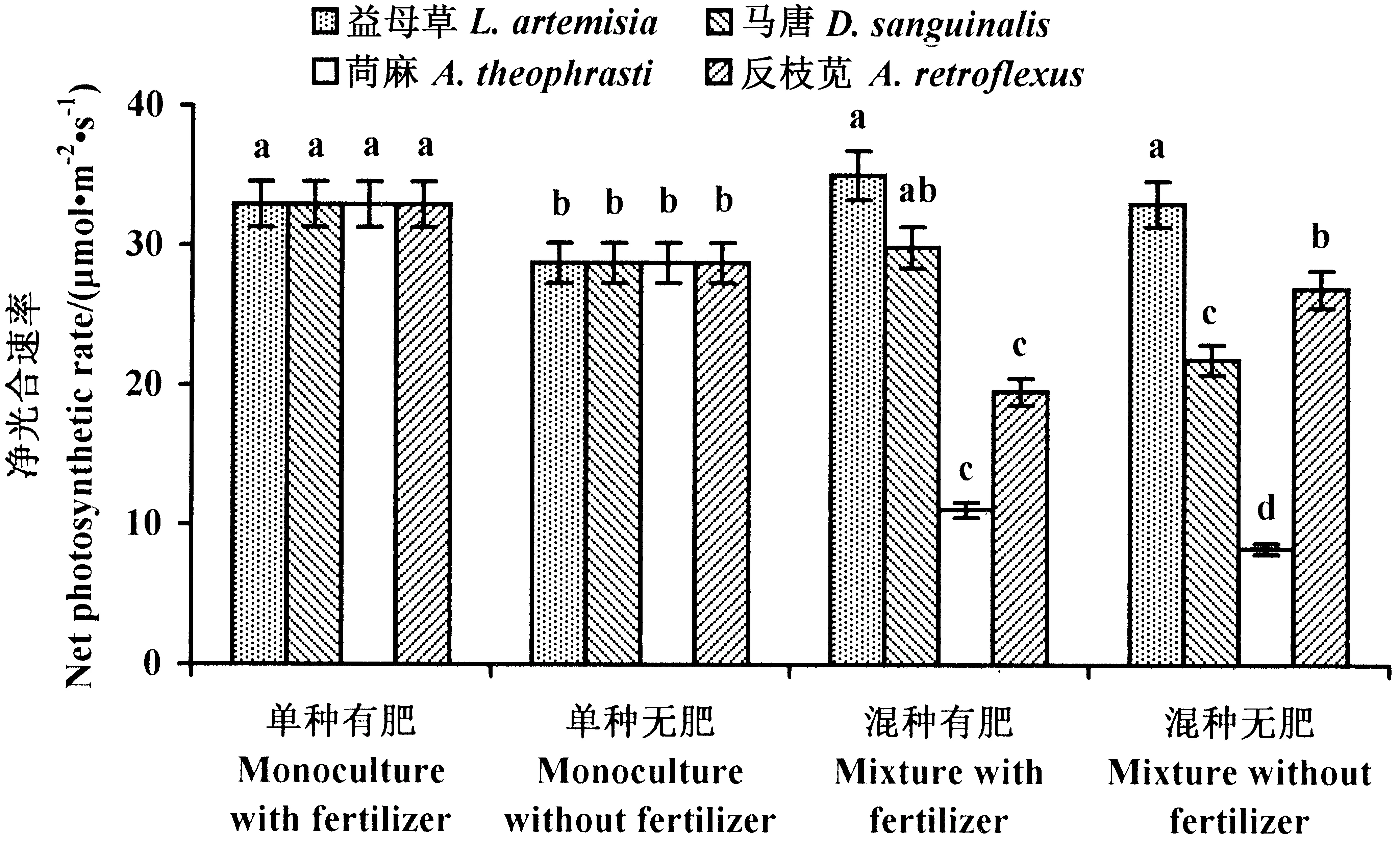
图2 黄顶菊净光合速率Fig.2 Net photosynthetic rate of the invasive palnt F. bidentis数据为平均值±标准差,不同小写字母表示植物在不同处理下差异显著(P<0.05)。 The datas are means±SD. Small letters indicate significant difference of plant in different treatments (P<0.05).
2.2 蒸腾速率
2.2.1 入侵域本地植物的蒸腾速率 与黄顶菊的竞争生长显著降低了益母草的蒸腾速率;单种处理中,施肥对益母草的蒸腾速率没有明显影响;混种处理中,施肥增强了益母草的蒸腾速率,但其显著低于单种有肥处理。施肥增强了单种处理中苘麻的蒸腾速率;与黄顶菊的竞争生长对苘麻的蒸腾速率没有影响。施肥和与黄顶菊竞争生长对马唐和反枝苋的蒸腾速率没有明显影响(图3)。
2.2.2 黄顶菊的蒸腾速率 单种处理中,施肥显著降低了黄顶菊的蒸腾速率。施肥促进了与益母草混种的黄顶菊的蒸腾速率,但对与马唐、苘麻和反枝苋混种的黄顶菊的蒸腾速率没有明显影响(图4)。
2.3 胞间CO2浓度
2.3.1 入侵域本地植物胞间CO2浓度 益母草在4种处理中的胞间CO2浓度均无显著差异(P>0.05),说明施肥与竞争生长对益母草胞间CO2浓度均无显著影响(图5)。
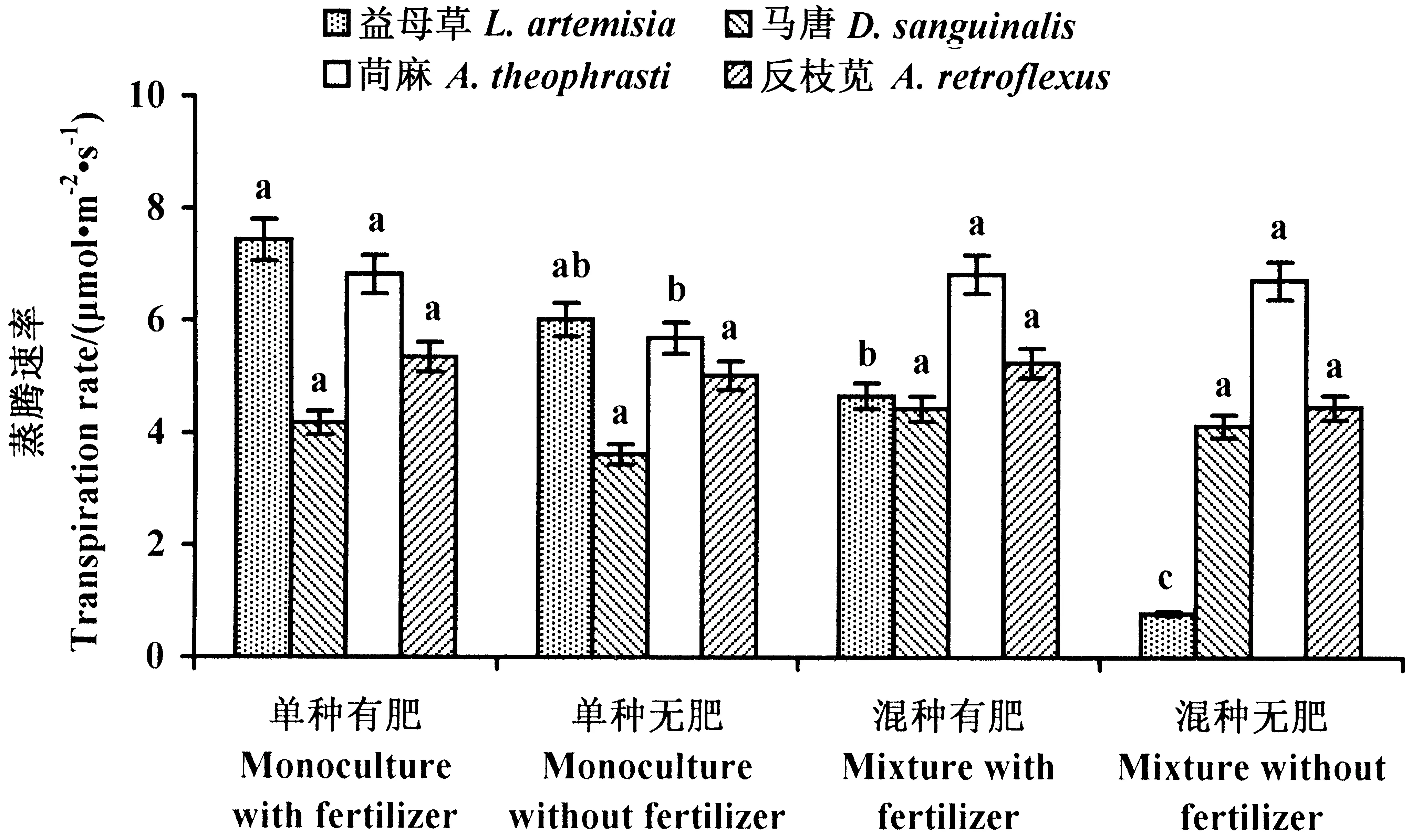
图3 本地植物蒸腾速率Fig.3 Transpiration rate of the four examined native weeds 数据为平均值±标准差,不同小写字母表示植物在不同处理下差异显著(P<0.05)。 The datas are means±SD. Small letters indicate significant difference of plant in different treatments (P<0.05).
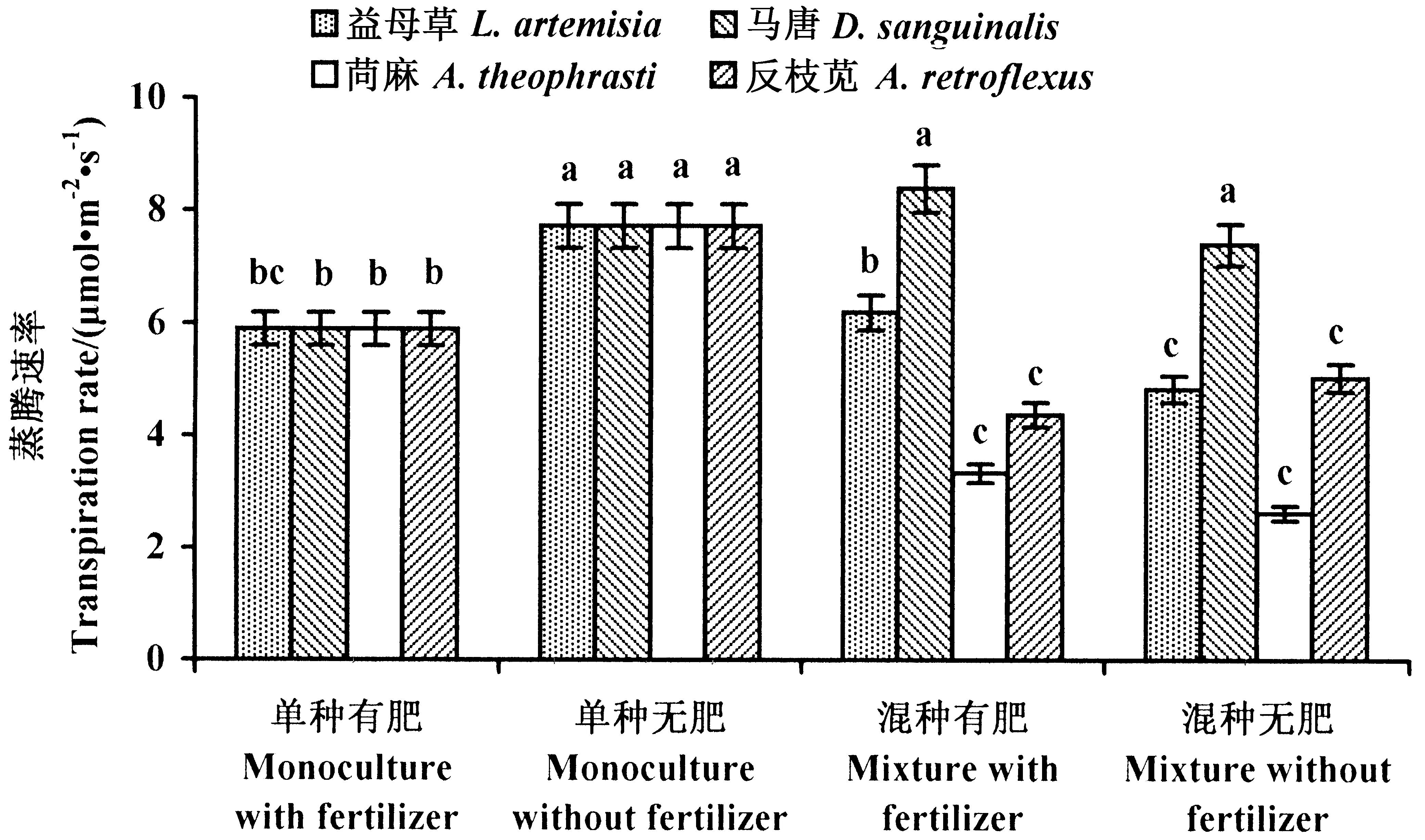
图4 黄顶菊蒸腾速率Fig.4 Transpiration rate of the invasive plant F. bidentis数据为平均值±标准差,不同小写字母表示植物在不同处理下差异显著(P<0.05)。 The datas are means±SD. Small letters indicate significant difference of plant in different treatments (P<0.05).
对马唐而言,单种处理下,施肥后的胞间CO2浓度显著高于未施肥处理(P<0.05),而在混种处理中,施肥与未施肥处理下胞间CO2浓度无显著差异(P>0.05);混种处理胞间CO2浓度均显著大于单种处理(P<0.05)。这说明混种处理显著抑制马唐的CO2利用率,而在单种处理下,无肥更有利于马唐对CO2的利用(图5)。
在苘麻的4种处理中,混种无肥处理胞间CO2浓度显著低于单种有肥和混种有肥处理(P<0.05),与单种无肥处理之间差异不显著(P>0.05),说明竞争生长对苘麻的CO2利用率没有影响(图5)。
反枝苋混种处理胞间CO2浓度显著高于单种处理(P<0.05),而施肥对于其单种与混种胞间CO2浓度的影响均不显著(P>0.05),说明混种对反枝苋的CO2利用率有显著抑制作用(图5)。
2.3.2 黄顶菊胞间CO2浓度 施肥显著抑制了黄顶菊对CO2的利用;与益母草、苘麻、反枝苋混种处理显著提高了黄顶菊对CO2的利用,而与马唐混种处理对黄顶菊CO2的利用率有显著抑制作用(图6)。
3 结论与讨论
外来生物成功入侵的因素有多方面,而外来植物自身的生理特性对于其种群的入侵、生存和扩展至关重要,有些外来物种具有比入侵域物种更强的光能利用力和光合响应机制,从而使其具有很强的入侵潜力(Baruch & Goldstein,1999; Durand & Goldstein,2001; Poorter,2001)。高的光合速率使入侵种能积累更多的碳用于生长和繁殖,为排挤本地种奠定了物质基础。植物的光合作用是一个非常复杂的生理过程,叶片光合速率与自身因素和环境因子密切相关。黄顶菊是双子叶植物中较为少见的C4植物(Powelletal.,1978) ,其入侵性与其光合特性有关(皇甫超河等,2009)。本研究发现,入侵域本地植物种类和土壤肥力是影响黄顶菊光合特性的主要因素。与益母草混种增强了黄顶菊的净光合速率,与马唐、苘麻和反枝苋混种显著降低了黄顶菊的净光合速率。吕远等(2011)研究发现,黄顶菊与反枝苋或苘麻混生时,黄顶菊的株高受到明显抑制,这可能与黄顶菊的光合速率降低密切相关。同时,单种处理中,施肥显著增强了黄顶菊的净光合速率、蒸腾速率和胞间CO2浓度。在黄顶菊与不同本地植物混种处理中,施肥对黄顶菊光合特性的影响与本地植物种类有关,如施肥增强了与马唐混种的黄顶菊的净光合速率,降低了与苘麻和反枝苋混种的黄顶菊的净光合速率,而对与益母草混种的黄顶菊的净光合速率没有影响。除了入侵域植物种类和土壤肥力等生态因子外,干旱、盐胁迫等环境因素也会影响黄顶菊的光合特性。如王秀彦等(2011)研究表明,干旱胁迫能降低黄顶菊叶片的净光合速率、蒸腾速率、气孔导度等生理指标,推迟净光合效率峰值出现的时间。
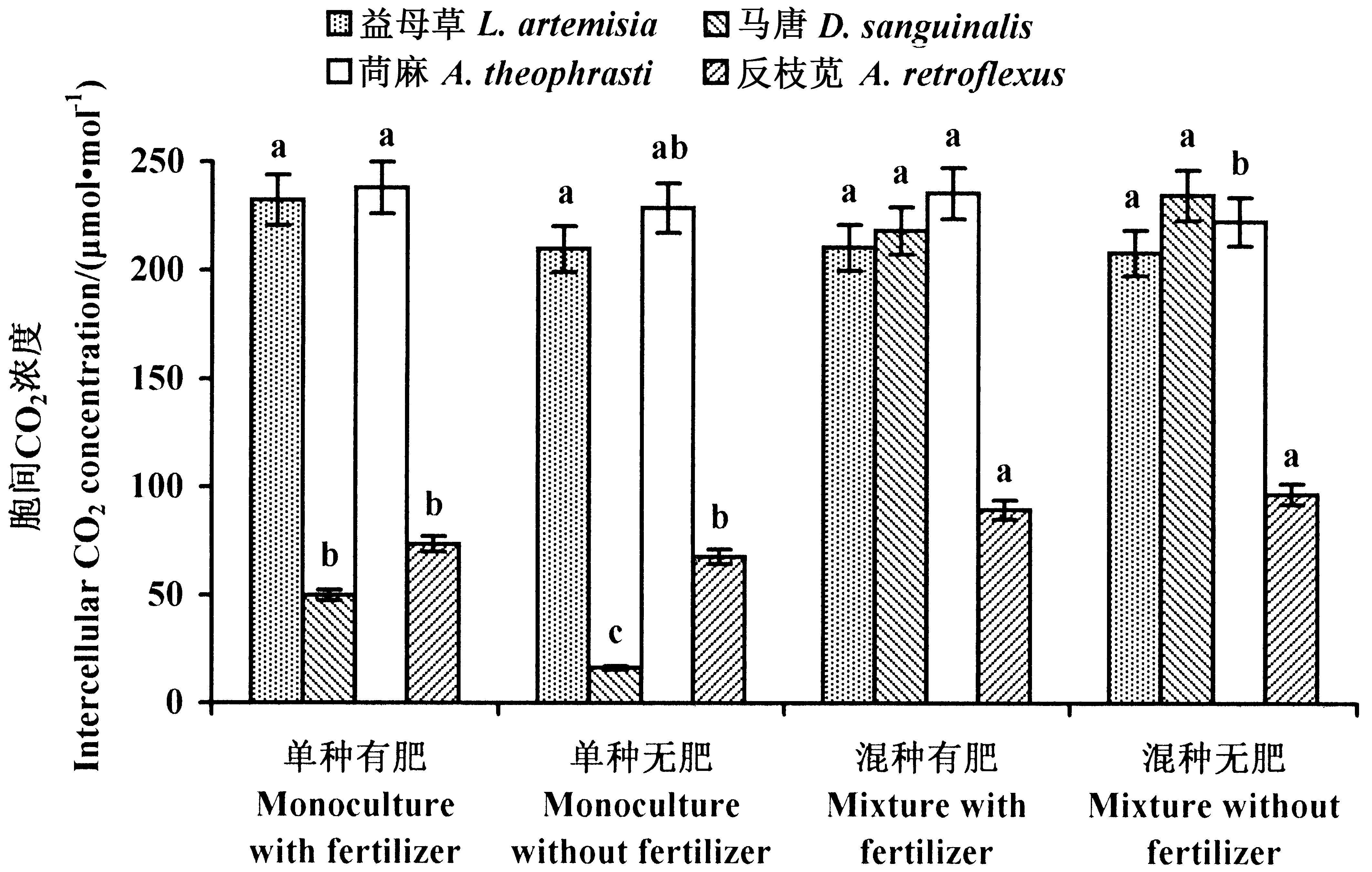
图5 本地植物胞间CO2浓度Fig.5 Intercellular CO2 concentration of the four examined native weeds 数据为平均值±标准差,不同小写字母表示植物在不同处理下差异显著(P<0.05)。 The datas are means±SD. Small letters indicate significant difference of plant in different treatments (P<0.05).
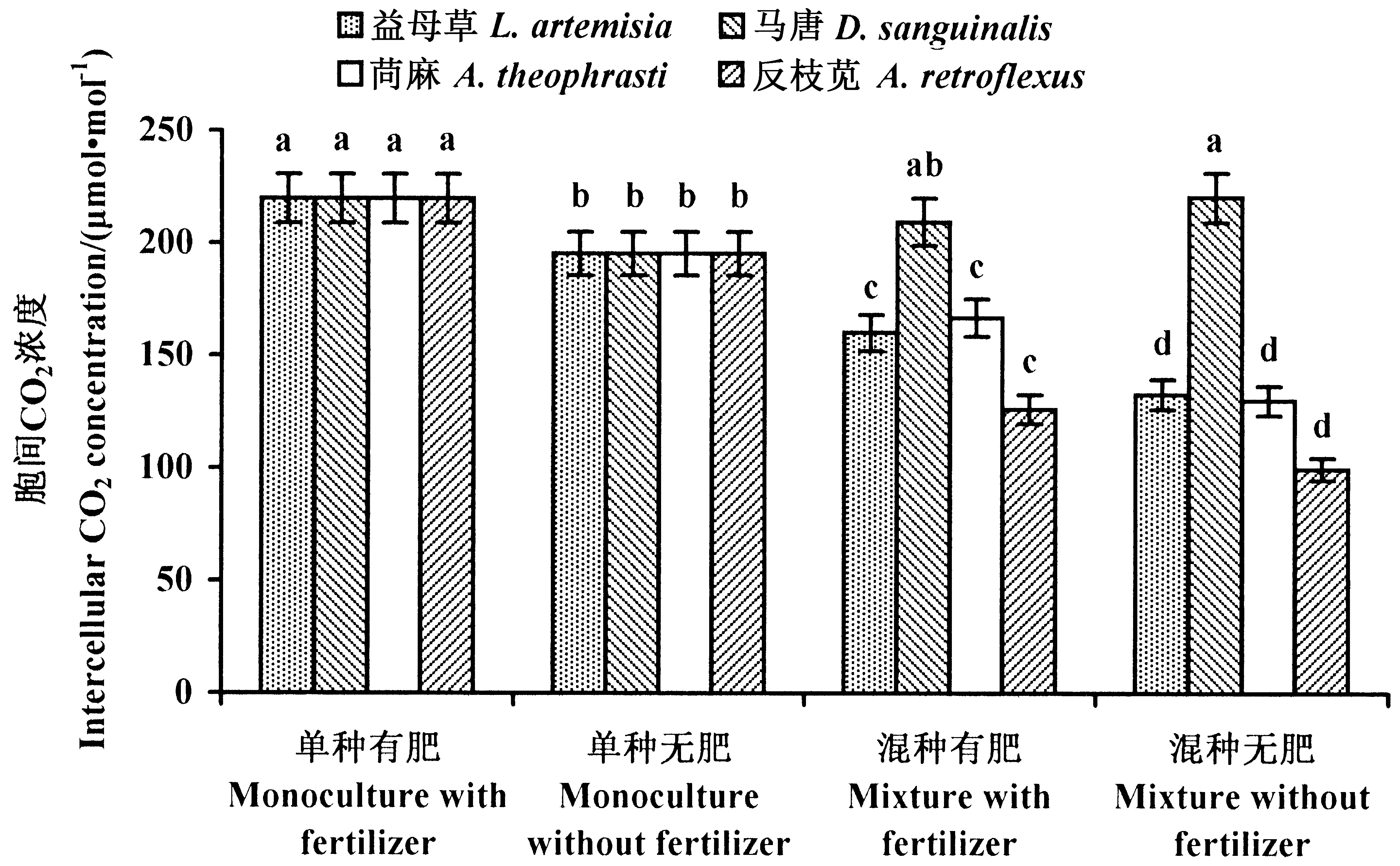
图6 黄顶菊胞间CO2浓度Fig.6 Intercellular CO2 concentration of the invasive plant F. bidentis数据为平均值±标准差,不同小写字母表示植物在不同处理下差异显著(P<0.05)。 The datas are means±SD. Small letters indicate significant difference of plant in different treatments(P<0.05).
本试验仅探讨了施肥和竞争生长条件下黄顶菊与其常见的主要伴生植物光合生理特征的差异,要全面了解黄顶菊的入侵机制,还需要进一步从繁育系统、抗逆性、种间竞争力等方面进行系统研究。
侯智勇, 洪伟, 李键, 林晗, 范海兰, 陈灿, 吴承祯. 2009. 不同桉树无性系光响应曲线研究. 福建林学院学报, 29(2): 114-119.
皇甫超河, 王志勇, 杨殿林. 2009. 外来入侵种黄顶菊及其伴生植物光合特性初步研究. 西北植物学报, 29(4): 781-788.
刘全儒. 2005. 中国菊科植物一新归化种——黄菊属. 植物分类学报, 43(2): 178-180.
吕远, 王贵启, 郑丽, 倪汉文. 2011. 入侵植物黄顶菊与本地植物的竞争. 生态学杂志, 30(4): 677-681.
莫凌, 唐文秀, 毛世忠, 黄仕训, 盘波, 骆文华. 2009. 珍稀濒危植物狭叶坡垒的光合特性. 福建林学院学报, 29(4): 357-361.
王俊峰, 冯玉龙, 梁红柱. 2004. 紫茎泽兰光合特性对生长环境光强的适应. 应用生态学报, 15(8): 1373-1377.
王秀彦, 阎海霞, 黄大庄. 2011. 干旱胁迫对黄顶菊光合特性的影响. 安徽农业科学, 12(3): 350-353, 358.
闫淑君, 洪伟, 吴承祯. 2006. 生物入侵对福建生态安全的影响. 福建林学院学报, 26(3): 275-280.
杨晴, 李婧实, 郭艾英, 齐艳玲, 李彦生, 张风娟. 2014. 遮阴和藜竞争对黄顶菊光合荧光和生长特性的影响. 应用生态学报, 25(9): 2536-2542.叶万辉, 冯惠玲. 2000. 外来入侵杂草薇甘菊及其伴生种基本光合特性的比较. 热带亚热带植物学报, 8(2): 139-146.朱慧, 马瑞君. 2009. 入侵植物马缨丹及其伴生种的光合特性. 生态学报, 29(5): 2701-2709.
Baruch Z and Goldstein G. 1999. Leaf construction cost, nutrient concentration, and net CO2assimilation of native and invasive species in Hawaii.Oecologia, 121: 183-192.
Bouman O T and Sylliboy J. 2012. Biomass allocation and photosynthetic capacity of willow (Salixspp.) bio-energy varieties.Forstarchiv, 83: 139-143.
Chen J W, Yang Z Q, Zhou P, Hai M R, Tang T X, Liang Y L and An T X. 2013. Biomass accumulation and partitioning, photosynthesis, and photosynthetic induction in field-grown maize (ZeamaysL.) under low- and high-nitrogen conditions.ActaPhysiologiaePlantarum, 35: 95-105.
Durand L A and Goldstein G. 2001. Photosynthes is, photoinhibition, and nitrogen use efficiency in native and invasive tree ferns in Hawaii.Oecologia, 126: 345-354.
Maria E, Alvarez M E and Cushman J H. 2002. Community-level consequences of a plant invasion: effects on three habitats in coastal California.EcologicalApplications, 12: 1434-1444.
Poorter L. 2001. Light dependent changes in biomass allocation and their importance for growth of rainforest trees species.FunctionalEcology, 15: 113-123.
Powell A M. 1978. Systematics ofFlavaria.AnnalsoftheMissouriBotanicalCanden, 65: 590-636.
Xing D K and Wu Y Y. 2015. Effects of low nutrition on photosynthetic capacity and accumulation of total N and P in three climber plant species.ChineseJournalofGeochemistry, 34: 115-122.
(责任编辑:杨郁霞)
The influence of fertilization and competition on photosynthetic characteristics of four native weeds and the invasive plantFlaveriabidentis
Xiao-ya ZHANG1+, Feng-min CHEN2+, Xue CHEN3, Jing YAN1, Ji-lin YIN1, Feng-juan ZHANG1*
1CollegeofLifeScience,HebeiUniversity,Baoding,Hebei071002,China;2TheThirdHighSchool,ChangliCounty,Qinhuangdao,Hebei066600,China;3HuihuaCollegeofHebeiNormalUniversity,Shijiazhuang,Hebei050091,China
【Background】Flaveriabidentishas strong ecological adaptability and competition ability. It can form a single dominant population through inhibiting the growth of native species, and then leads to loss of biodiversity. 【Method】F.bidentisand its 4 companion species,Leonurusartemisia,Abutilontheophrasti,DigitariasanguinalisandAmaranthusretroflexuswere studied. Photosynthetic characteristics ofF.bidentisand the four companion plant species were measured using LI-6400 portable photosynthesis system. The changes of photosynthesis rate (Pn), transpiration rate (Tr) and intercellular CO2concentration (Ci) during competitive growth between different plants andF.bidentiswere also studied. 【Result】The effect of competition on thePnofF.bidentiswas tightly related with the kinds of native weeds. Fertilization promoted itsPn. Competitive growth significantly inhibitedPnofL.artemisia,D.sanguinalisandA.retroflexusand promotedPnofA.theophrasti. Fertilization promotedPnof the four native weeds. Competitive growth had an obvious influence onTrofL.artemisiaandA.theophrasti, but it had less little effect onTrof the other two native weeds;TrofF.bidentiswas lower than that competed withA.theophrasti,A.retroflexusorL.artemisia. Fertilization significantly inhibitedTrinF.bidentisgrown in monoculture.CiofF.bidentiswas decreased obviously when it competed withL.artemisia,A.theophrastiandA.retroflexusrespectively. Fertilization addition significantly inhibitedCiofF.bidentisin monoculture treatment. 【Conclusion and significance】Fertilization affected the photosynthetic charateristics ofF.bidentis. The effect of different native plant species on the photosynthesis of the invasive plant species was different. The research results would help to discover the invasive mechanism and to provide the theoretical basis to control the invasive plant species.
invasive biology;Flaveriabidentis; photosynthetic characteristics; fertilization; competition
2015-06-11 接受日期(Accepted): 2015-07-08
国家自然科学基金(31040066、31171906); 河北省自然科学基金(C2015201021)
张晓亚, 女, 硕士研究生。 研究方向: 入侵生物学。 E-mail: xiaoyazhang2015@163.com。 陈凤敏, 女, 讲师。 研究方向: 生态学。 E-mail: 591348993@qq.com
10. 3969/j.issn.2095-1787.2015.03.007
+同等贡献作者(The two authors contributed equally to this work)
*通讯作者(Author for correspondence), E-mail: fengjuanzhang@126.com
猜你喜欢
植物保护(2024年3期)2024-06-08 07:17:41
民间故事选刊(2022年20期)2022-10-20 06:27:16
种子(2022年8期)2022-10-10 11:07:48
民间故事选刊·下(2022年10期)2022-05-30 15:51:26
江苏农业科学(2017年24期)2018-01-12 11:39:25
环境科技(2017年1期)2017-09-05 03:08:02
农业与技术(2017年6期)2017-04-21 00:01:27
农业研究与应用(2016年6期)2017-04-15 20:25:16
安徽农学通报(2015年8期)2015-07-02 01:42:09
成长·读写月刊(2015年4期)2015-06-18 21:13:37
