
1株黑鲷致病性假交替单胞菌的鉴定及毒力基因分析
2015-06-15 15:53乔毅沈辉万夕和戚原野王李宝
江苏农业科学 2015年4期
乔毅+沈辉+万夕和+戚原野+王李宝


摘要:从患有尾鳍基部溃烂病的人工养殖黑鲷(Acanthopagrus schlegel)病灶处分离到1株优势菌LSHD-1,经人工感染试验证实为引起黑鲷尾鳍基部溃烂病病原菌。细菌形态学观察、生理生化等表观、生物学试验、细菌16S rRNA基因同源性检索和系统发育学分析结果表明,LSHD-1与假交替单胞菌属(Pseudoalteromonas)的吻合程度最高,系统发育分析结果与杀鱼假交替单胞菌(Pseudoalteromonas piscicida)聚为1支。扩增细菌毒力基因,分析其潜在致病性,发现LSHD-1携带黏附侵袭位点基因(ail)、毒力活化因子(virF)、P4噬菌体整合酶基因(intB)、溶血素基因(hlyA)、丝氨酸蛋白酶基因(ahpA)、肠毒素基因(altA)、抗金属蛋白酶基因(ast)。药敏试验结果显示,LSHD-1对氧氟沙星、恩诺沙星、诺氟沙星、头孢曲松、氟苯尼考高度敏感,对磺胺异唑、强力霉素、青霉素G、四环素表现出较强耐药性。
关键词:黑鲷;假交替单胞菌;溃烂病;系统发育;毒力基因;药物敏感性
中图分类号: S941.42 文献标志码: A
文章编号:1002-1302(2015)04-0229-04
收稿日期:2015-01-04
基金项目:江苏省水产三新工程项目(编号:D2013-5-2) 。
作者简介:乔 毅(1987—),男,硕士研究生,主要从事水产养殖病害防治技术研究。 E-mail:qiaoyishou@yeah.net。
通信作者:万夕和,研究员,主要从事水产养殖病害防治技术研究。 E-mail:wxh1708@163.com。
假交替单胞菌属是1995年Gauthier等根据细菌16S rRNA序列发现有别于假单胞菌(Pseudomonas)和交替单胞菌(Alteromonas)的一个新属[1],目前已经发现30多个种。该菌广泛分布于海洋沉积物以及共附生在一些海洋动植物中。假交替单胞菌能产生胞外毒素、胞外多糖、胞外酶以及病毒活性物质等与致病相关的多种物质[2],是一类条件致病菌。研究表明,假交替单胞菌可感染海水养殖藻类和养殖动物等,引起藻类溶藻、绿斑、黄斑[3-5]等,养殖动物皮肤溃烂[6]等疾病。 黑鲷(Acanthopagrus schlegel)属于鲈形目鲈亚目鲷科鱼类,主要分布在西太平洋区,在我国沿海均有分布,是海水鱼养殖主要品种之一。近年来,江苏沿海地区黑鲷在养殖过程中经常发生皮肤溃烂,并出现不同程度的死亡,给黑鲷养殖产业带来严重威胁。2012年4月,江苏省海水增养殖技术及种苗中心人工养殖黑鲷过程中发生皮肤溃烂,并出现一定规模的死亡。本试验从患病鱼体分离到1株优势菌LSHD-1,对该菌进行了鉴定和毒力基因的研究,并进行了药物敏感性试验。
1 材料与方法
1.1 材料
试菌株LSHD-1,分离自江苏省海水增养殖技术及种苗中心的池塘养殖患皮肤溃烂病黑鲷,该菌株已通过回复感染证实为该病的病原菌。
2216E培养基、细菌微量生化鉴定管、水解酪蛋白琼脂(MH琼脂)、质控菌(大肠杆菌ATCC25922)及药敏试纸购自杭州天和微生物试剂有限公司;Premix Taq酶、DNA marker购自TaKaRa公司;引物(16S rRNA,毒力基因)由生工生物工程(上海)股份有限公司合成;其他试剂均为分析纯。
1.2 方法
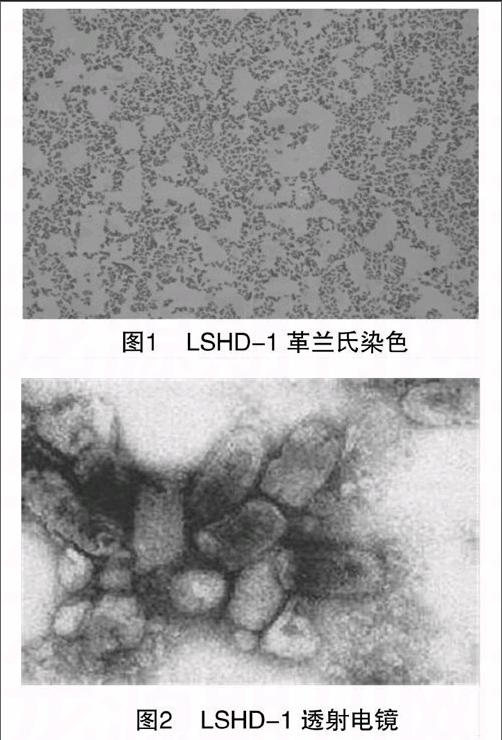
1.2.1 形态结构观察 用接种环挑取少量细菌在载玻片上涂片固定,革兰氏染色,油镜观察形态。挑取单菌落细菌用海水制成菌悬液,吸取1滴置于铜网膜上,通过醋酸铀染色,日立H-300型透射电子显微镜观察。
1.2.2 生理生化试验 参照《常见细菌系统鉴定手册》和《Bergeys Manual of Determinative Bacteriology》进行生理生化试验。
1.2.3 细菌16S rRNA基因扩增 挑取单菌落于装有 100 μL 灭菌去离子水的1.5 mL EP管中,煮沸10 min,4 ℃ 10 000 r/min离心5 min,上清即为聚合酶链反应(polymerase chain reaction,PCR)模板DNA,-20 ℃保存备用。采用通用引物扩增16S rRNA基因序列与测序,正向引物27F(5′-AGAGTTTGATCCTGGCTCAG-3′),反向引物1492R(5′-GGTT ACCTTGTTACGACTT-3′)。25 μL反应体系:12.5 μL Premix Taq酶,正、反向引物各1 μL,模板DNA 1 μL,ddH2O补足25 μL。反应条件:94 ℃5 min;94 ℃30 s,54 ℃30 s,72 ℃ 1.5 min,35个循环;72 ℃10 min。PCR产物经1%琼脂糖电泳(120 V,35 min)检测目的片段(约1 500 bp)后,送生工生物工程(上海)股份有限公司测序。
1.2.4 系统发育分析 利用National Center for Biotechnology Information(NCBI)数据库中blast对LSHD-1细菌16S rRNA序列进行相似性比较。使用ClustalX2.0软件与从GenBank数据库中获得序列相似性较高的序列进行多序列比对,通过MEGA4(molecular evolutionary genetics analysis,MEGA)软件采用邻接建树法构建系统发育树,并通过Bootstrap法(1 000次)检验。
1.2.5 毒力基因检测 通过PCR对LSHD-1菌黏附侵袭位点基因(ail)、耐热肠毒素基因(ystB)、黏附素基因(yadA)、毒力活化因子(virF)、P4噬菌体整合酶基因(intB)、气溶素基因(aerA)、溶血素基因(hlyA)、丝氨酸蛋白酶基因(ahpA)、肠毒素基因(altA)以及抗金属蛋白酶基因(ast)进行扩增。扩增条件:ail、ystB、yadA、virF、intB按照94 ℃预变性4 min;94 ℃ 变性45 s、退火45 s(退火温度见表1),72 ℃延伸40 s,35个循环;72 ℃终延伸5 min。aerA、hlyA、ahpA、altA、ast基因94 ℃预变性5 min;94 ℃变性30 s、退火30 s(退火温度见表1),72 ℃延伸30 s,35个循环;72 ℃终延伸10 min。PCR产物经1%琼脂糖电泳检测结果。endprint
表1 检测的毒力基因引物序列及反应条件
基因名称 引物序列(5′→3′) 长度
(bp) 退火温度
(℃)
Ail[7] F:TAATGTGTACGCTGCGAG;R:GACGTCTTACTTGCACTG; 351 50
ystB[7] F:CTTCAGATACTGGTGTCGCTGT;R:ATGCCTGACTAGAGCGATATCC 200 56
yadA[7] F:GGCAGAACAGCAGTCAGACATA;R:GGTGAGCATAGAGAATACGTCG 800 56
virF[7] F:GTACATTAGGCCAAGAGACG;R:GCAACATACCTCACAACACC 561 45
intB[7] F:TGCGCCATGCGGTCCATC;R:GGTGCATAAGATTCTCGG 714 50
aerA[8] F:CAAGAACAAGTTCAAGTGGCCA;R:ACGAAGGTGTGGTTCCAGT 309 57
hlyA[9] F:TGACAGGCAAGTAGAATAACGC;R:TGTCCGCTTTCCACTCCC 1 815 53
ahpA[10] F:GTTAGGCGTTGGCAATCTCG;R:CGCTGGAGTAGGAGGAACG 874 60
altA[11] F:ATCGTCAGCGACAGCTTCTT;R:CTCATCCCTTGGCTTGTTGT 442 58
ast[12] F:TGACCCAGTCCTGGCACGGC;R:GCTGATCGATCACCACCAGC 504 57
1.2.6 药物敏感性试验 选择磺胺异唑、强力霉素、氧氟沙星、青霉素G、恩诺沙星、新霉素、头孢曲松、四环素、红霉素、诺氟沙星、氟苯尼考药敏试纸片对LSHD-1进行敏感性试验,采用世界卫生组织推荐的K-B纸片琼脂扩散法进行,结果按敏感(S)、中敏(I)、耐药(R)报告[13]。
2 结果与分析
2.1 细菌形态学特征
LSHD-1菌落在2216E琼脂平板上为浅黄色,凸透镜状,表面光滑湿润有光泽,边缘完整,不透明,圆形。革兰氏染色阴性,呈短杆状(图1)。负染后电镜观察,菌体呈短杆状,极端单鞭毛(图2)。
2.2 生理生化特征
LSHD-1菌氧化酶试验阳性,L-鸟氨酸、L-精氨酸、L-赖氨酸和L-色氨酸反应阴性(表2),结合伯杰氏手册,该生理生化结果与假交替单胞菌基本符合。
2.3 分子生物学鉴定及系统发育学分析
PCR扩增LSHD-1菌16S rRNA基因获得1条清晰的条
表2 LSHD-1生理生化试验
反应(酶) 结果 反应(酶) 结果
鸟氨酸脱羧酶 - 吲哚(产生) -
精氨酸双水解酶 - N-乙酰-β-葡萄糖苷酶 +
赖氨酸脱羧酶 - β-半乳糖苷酶 -
脲酶 - 葡萄糖(产酸) -
L-阿拉伯糖醇(产酸) - 蔗糖(产酸) -
半乳糖羧盐(产酸) - L-阿拉伯糖(产酸) -
5酮基-葡萄糖酸(产酸) - D-阿拉伯醇(产酸) -
肢酶 - α-葡萄糖苷酶 +
酚红(产酸) - α-半乳糖苷酶 -
β-葡萄糖苷酶 + 海藻糖(产酸) -
甘露醇(产酸) - 鼠李糖(产酸) -
麦芽糖(产酸) - 肌醇(产酸) -
侧金盏花醇(产酸) - 纤维二糖(产酸) -
帕拉金糖(产酸) - 山梨醇(产酸) -
β-葡萄糖酸苷酶 - 麦芽糖苷酶 -
丙二酸 - L-天冬氨酸芳胺酶 +
带,经测序后发现为1 438个碱基的基因片段,将该序列在NCBI数据库进行blast分析对比,发现LSHD-1与假交替单胞菌杀鱼亚种的同源性最近,达99%。将测得序列与GenBank中已登录的16S rRNA序列进行匹配排列,构建系统进化树,结果LSHD-1与假交替单胞菌杀鱼亚种聚为1支(图3)。
2.4 毒力基因检测结果
PCR扩增LSHD-1细菌毒力基因,经电泳检测发现:altA、ail、intB、ahpA、ast、virF、hlyA基因(图4、图5)出现相应大小的条带,其中丝氨酸蛋白酶基因(ahpA)条带最为清晰,altA、intB、virF条带比较清晰,ail、ast、hlyA条带相对较弱。
2.5 药物敏感性试验
药敏试验结果表明,LSHD-1菌对氧氟沙星、恩诺沙星、头孢曲松、诺氟沙星、氟苯尼考敏感;对磺胺异唑、强力霉素、青霉素G、四环素表现出耐药;磺胺异唑和青霉素G对LSHD-1菌的抑菌效果较差(表3)。
3 讨论
假交替单胞菌是海洋中常见的细菌,随着海水养殖业的发展,假交替单胞菌受到广泛的关注和研究。目前,日本有关
于假交替单胞菌引起海带(Laminaria)孢子体红点病[14]和孔烂症[15]的报道,我国也有假交替单胞菌引起条斑紫菜(Porphyrae yezoensis)绿斑病[4]和黄斑病[5]的报道;Pujalte等从患病的海鲈体内分离到1株Pseudoalteromonas undina,试验发现其毒性较弱,但是在养殖环境恶化时引起金头鲷和海鲈患病[16];国内大量研究表明,假交替单胞菌可引起刺参(Apostichopus japonicus)腐皮病[17-19]、溃疡病[20]等;乔迁等报道假交替单胞菌引起七带石斑鱼(Epinephelus septemfasciatus)皮肤溃烂[21];白雪松等从群体死亡日本对虾(Penaeus japonicus)蚤状幼体中分离到1株致病菌,经鉴定为Pseudoalteromonas lipolytica[22]。本试验从患病黑鲷病灶处分离到1株优势菌LSHD-1, 革兰氏染色和透射电镜观察结果与假交替单胞菌endprint
表3 LSHD-1药物敏感性试验结果
药敏试纸
名称 药物含量
(μg/片)
抑菌圈大小(mm)
ATCC25922 LSHD-1
敏感性
磺胺异唑 300 24.6 0 R
强力霉素 30 18.4 8.0 R
氧氟沙星 5 31.5 23.4 S
青霉素G 10 15.7 0 R
恩诺沙星 15 38.0 26.6 S
新霉素 30 23.5 16.7 I
头孢曲松 30 34.0 27.9 S
四环素 30 21.5 13.5 R
红霉素 15 35.6 15.8 I
诺氟沙星 10 24.3 18.7 S
氟苯尼考 30 34.6 22.4 S
的特征基本一致;根据生理生化表型特征,LSHD-1菌与假交替单胞菌相似度最高;另外,LSHD-1细菌的16S rRNA基因序列在NCBI数据库中通过blast同源性检索,基因序列与假交替单胞菌杀鱼亚种的同源性最高,达到99%,从基因系统发育树来看,LSHD-1与假交替单胞菌杀鱼亚种聚为1支。综合LSHD-1的形态学特征、生理生化和分子生物学鉴定结果,认为LSHD-1为假交替单胞菌杀鱼亚种。
黑鲷在养殖过程中,相关报道过有拟态弧菌(Vibrio mimicus)引起的幼鱼腹水病[23]、哈维氏弧菌(Vibrio harveyi)[24]与迟缓爱德华氏菌(Edwardsiella tarda)[25]引起的大规模死亡。假交替单胞菌杀鱼亚种感染黑鲷的研究未见报道,毒力人工回感试验结果表明,该株菌为引起黑鲷尾鳍基部溃烂病的病原菌;通过扩增LSHD-1菌的毒力基因,结果检测到黏附侵袭位点基因(ail)、毒力活化因子(virF)、P4噬菌体整合酶基因(intB)、溶血素基因(hlyA)、丝氨酸蛋白酶基因(ahpA)、肠毒素基因(altA)、抗金属蛋白酶基因(ast)。ail基因能够编码对宿主产生高度黏附并保护细菌不被宿主免疫系统杀灭的小分子黏附蛋白[26],目前,ail基因在小肠耶尔森氏菌中研究较多[7,27-28],有学者甚至认为,可将ail基因作为小肠耶尔森氏菌致病性指示基因[29];ahpA基因是细菌重要的致病基因之一,其主要在病原入侵过程中通过活化毒力因子间接发挥作用[30],张璐等利用缺失突变法构建迟缓爱德华氏菌ahpA基因突变菌株,通过与野生菌株的侵染试验的比较,认为ahpA基因对迟缓爱德华氏菌前期侵染宿主有关[31],朱大玲等研究表明,ahpA基因阳性的嗜水气单胞菌(Aeromonas hydrophila)为有毒菌[32];hlyA基因是多数致病细菌的常见基因,关于其在致病过程中所起的作用,付乔芳等认为,hlyA基因与嗜水气单胞菌的致病力关系不大[33],但黄钧等发现携带hlyA基因的气单胞菌致病力增强[34];virF基因是编码毒力基因的调控因子[35],在细菌致病性中起着重要作用,altA、ast基因在已有报道中证实与致病性关系不大[33]。本试验还扩增到intB基因,与苟小兰等的试验结果[7]一致,说明LSHD-1染色体上存在高致病性毒力岛。综上可以推断,本试验分离的致病性杀鱼假交替单胞菌的致病力可能主要由ail、ahpA、virF、intB基因决定。毒力基因的量、毒力基因的表达机理以及毒力基因间的相互作用与致病性的关系非常复杂,引起黑鲷的致病机制尚需进一步研究。
LSHD-1药物敏感性试验结果与乔迁等的研究结论[21-22]基本一致,氧氟沙星、恩诺沙星、诺氟沙星、头孢曲松、氟苯尼考对LSHD-1有较好的抑菌效果;该菌对磺胺异唑、强力霉素、青霉素G、四环素表现出较强耐药性。但是总体药物敏感性比淡水菌敏感,这可能是因为海水养殖抗菌药物使用的频率不高,细菌在没有抗菌素环境压力下保持了较高的药物敏感性。该菌的药物敏感性试验结果可为防治该菌引起的水生动物疾病提供借鉴和参考。
参考文献:
[1]Gauthier G,Gauthier M,Christen R. Phylogenetic analysis of the genera Alteromonas,Shewanella and Moritella using genes coding for small-subunit rRNA sequences and division of the genus Alteromonas into two genera,Alteromonas (emended) and Pseudoalteromonas gen. nov.,and proposal of twelve new species combinations[J]. International Journal of Systematic Bacteriology,1995,45(4):755-761.
[2]席 宇,朱大恒,刘红涛,等. 假交替单胞菌及其胞外生物活性物质研究进展[J]. 微生物学通报,2005,32(3):108-112.
[3]黄 姿,李春强,于晓玲,等. 1株溶藻细菌(Pseudoalteromonas sp.)的分离鉴定及溶藻活性初探[J]. 海洋技术,2008,27(3):56-60.
[4]闫 咏,马家海,许 璞,等. 1 株引起条斑紫菜绿斑病的柠檬假交替单胞菌[J]. 中国水产科学,2002,9(4):353-358.
[5]王洪斌,李信书,夏亚明,等. 条斑紫菜丝状体黄斑病病原体分离鉴定及生物学特性研究[J]. 海洋环境科学,2011,30(3):361-364,408.endprint
[6]张 莹,姜国良,刘 云,等. 仿刺参“腐皮综合症”病灶处优势菌的分离鉴定及AHLs信号分子的检测[J]. 微生物学通报,2009,36(11):1664-1669.
[7]苟小兰,王 利. 小肠结肠炎耶尔森氏菌对黄颡鱼的致病性及毒力基因检测[J]. 水产科学,2013,32(5):293-296.
[8]Wang G H,Clark C G,Liu C Y,et al. Detection and characterization of the hemolysin genes in Aeromonas hydrophila and Aeromonas sobria by multiplex PCR[J]. Journal of Clinical Microbiology,2003,41(3):1048-1056.
[9]Heuzenroeder M W,Wong C Y F,Flower R L P. Distribution of two hemolytic toxin genes in clinical and environmental isolates of Aromonas spp. correlation with virulence in a suckling mouse model[J]. Federation of European Microbiological Societies Microbiology Letters,1999,174(2):131-136.
[10]储卫华,陆承平. 聚合酶链反应(PCR)法检测嗜水气单胞菌丝氨酸蛋白酶基因[J]. 中国兽医学报,2003,23(5):463-464.
[11]Nawaz M,Khan S A. Detection and characterization of virulence genes and integrons in Aeromonas veronii isolated from catfish[J]. Food Microbiology,2009,1:1-5.
[12]El-Barbary M I. Some clinical microbiological and molecular characteristics of Aeromonas hydrophila isolated from various naturally infected fishes[J]. Aquacuture International,2010,18(5):943-954.
[13]谭 瑶,赵 清,舒为群,等. K-B纸片扩散法药敏试验[J]. 检验医学与临床2010,7(20):2290-2291.
[14]Sawabe T,Makino H,Tatsumi M,et al. Pseudoalteromonas bacteriolytica sp.nov.,a marine bacterium that is the causative agent of red spot disease of Laminaria japonica[J]. International Journal of Systematic Bacteriology,1998,48:769-774.
[15]Sawabe T,Tanaka R,Lqbal M M,et al. Assignmen t of Alteromonas elyakovii KMM 162T and five strains isolated from spot-wounded fronds of Laminaria japonica to Pseudoalteromonas elyakovii comb. nov. and the description of the species[J]. International Journal of Systematic Bacteriology,2000,50:265-271.
[16]María J,Ariadna S B,María C M,et al. Occurrence and virulence of Pseudoalteromonas spp. in cultured gilthead sea bream (Sparus aurata L.) and European sea bass (Dicentrarchus labrax L.). Molecular and phenotypic characterisation of P. undina strain U58[J]. Aquaculture,2007,271:47-53.
[17]王印庚,方 波,张春云,等. 养殖刺参保苗期重大疾病“腐皮综合征”病原及其感染源分析[J]. 中国水产科学,2006,13(4):610-616.
[18]谢建军.养殖刺参(Apostichopus japonicus) 腐皮综合症主要致病菌快速检测技术研究[D]. 上海:上海水产大学,2007.
[19]葛 辉,何丽斌,林 琪,等. 福建海区网箱养殖刺参“腐皮综合症”病原分析与鉴定[J]. 福建水产,2012,34(2):105-109.
[20]孟庆国,吴刘记,吴信忠,等. 养殖刺参溃疡病病原学研究[J]. 水产科学,2006,25(12):635-639.
[21]乔 迁,韩娜娜,陈 超,等. 致病性假交替单胞菌的分离鉴定及药敏特性研究[J]. 安徽农业科学,2010,38(26):14224-14226,14232.
[22]白雪松,张晓君,比可然,等. 1株假交替单胞菌的分离与鉴定[J]. 江苏农业科学,2013,41(10):211-216.endprint
[23]王国良,郑天伦,金 珊,等. 黑鲷幼鱼腹水病病原菌[J]. 中国兽医学报,2003,23(1):33-34.
[24]潘晓艺,沈锦玉,尹 文,等. 哈维氏弧菌黑鲷分离株BK-1培养条件优化研究[J]. 浙江海洋学院学报:自然科学版,2005,24(3):240-243.
[25]闰茂仓,王雪鹏,丁 雷. 养殖黑鲷迟缓爱德华菌的分离鉴定[J]. 山东畜牧兽医,2010,31(4):19,22.
[26]Yamashits S,Lukacik P,Barnard T J,et al. Structural insights into Ail-mediated adhesion in Yersinia pestis[J]. Structure,2011,19(11):1672-1682.
[27]王 军,李跃旗,赵 蕊,等. 致病性小肠结肠炎耶氏菌DNA多态性[J]. 解放军医学杂志,2002,27(1):46-47.
[28]Balak R,Murali H S,Batra H V. Cloning expression and characterization of attachment-invasion locus protein(Ail) of Yersinia enterocolitica and its utilization in rapid detection by immunoassays[J]. Lett Appl Microbio1,2010,50(2):131-137.
[29]Huang Y,Wang X,Cui Z,et al. Possible use of ail and foxA polymorphisms for detecting pathogenic Yersinia enterocolitica[J]. BMC Microbiology,2010(10):211-217.
[30]Vipond R. Defined deletion mutants demonstrate that the major secreted toxin are not essential for the virulence of Aeromonas salmonicida[J]. Infection and Immunity,1988,66(5):1990-1998.
[31]张 璐,张书仁,孙铂光. 丝氨酸蛋白酶Pic在迟缓爱德华氏菌侵染鱼类中的作用[J]. 山东农业科学,2014,46(7):15-19.
[32]朱大玲,李爱华,汪建国,等. 嗜水气单胞菌毒力与毒力基因分布的相关性[J]. 中山大学学报:自然科学版,2006,45(1):82-85.
[33]付乔芳,邱军强,胡 鲲,等. 嗜水气单胞菌国内分离株的毒力因子分布与致病性相关性分析[J]. 生物学杂志,2011,28(6):53-57.
[34]黄 钧,黄艳华,胡大胜,等. 黄沙鳖白底板病病原菌的分离鉴定及6 种毒力基因检测[J]. 水生生物学报,2013,37(5):844-854.
[35]Lambert-Rouvroit C,Sluiters C,Cornelis G R.Role of the transcriptional activator,VirF,and temperature in the expression of the pYV plasmid genes of Yersinia enterocolitica[J]. Mol Micmbio1,1992,6(3):395-409.endprint
