
不同强度干旱胁迫对刺槐幼苗生理生化特性的影响
2015-05-22 06:01
中南林业科技大学学报 2015年8期
(娄底职业技术学院,湖南 娄底 417000)
刺槐Robinia pseudoacacia,又名洋槐,属豆科蝶形花亚科刺槐属的落叶乔木。其萌芽力和根蘖性强,生长迅速,树冠高大宽阔,枝叶浓郁,叶色鲜绿,每年4~6月花开季节绿白相映,素雅而芳香,已经成为我国南北各地广泛栽培的优良庭荫树种与行道树绿化树种;同时,因其适应性强,耐干旱贫瘠,更是立地条件差、环境污染重的工矿区绿化及荒山荒地绿化的先锋树种。目前对刺槐的研究主要集中在造林技术、生态系统的影响等方面[1-2],但有关刺槐抗旱生理方面研究较少[3-4]。本研究通过采用盆栽人工控水方法,对一年生刺槐实生苗进行4种不同强度持续干旱胁迫模拟试验,旨在分析探讨刺槐幼苗对不同强度干旱胁迫的生理生化指标变化特性与适应机制,其结果可为刺槐在城乡园林绿化中的科学节水灌溉与合理利用及解决生长发育中的水分供需矛盾提供理论指导与技术支撑。
1 材料与方法
1.1 试验材料及处理方法
供试材料为1 a刺槐实生苗。试验地点设在娄底职业技术学院农林实训基地。2012年3月28日将长势基本一致80 株1 a刺槐实生苗稍带土球上盆到塑料花盆中培育,每盆1株。盆土为混合基质(园土∶有机肥∶沙土=40%∶40%∶20%)。2012年6月28日(即盆栽正常养护3个月后)从按统一规格标准从中选择60盆(每盆1株)浇透水供试验备用。
1.2 试验设计
通过随机区组设计,将供试的60盆刺槐苗随机分为4个处理组,依次为:CK(对照)、LS(轻度干旱)、MS(中度干旱)、HS(重度干旱),其土壤相对含水量分别为75~80%、55~60%、35~40%、15~20%,每组各设3个重复。试验全程土壤相对含水量采用人工称重法控制,每天傍晚对所有供试盆栽苗称重1次,按计算的补水量补充当天消耗的水分(为扣除盆栽过程中的自然蒸发失水,试验设置了没有栽培植物的相同盆土作为参照),使土壤含水量始终控制在设定范围内。
1.3 测定项目及方法[5]
可溶性糖(SS)含量采用蒽酮法测定;游离脯氨酸(Pro)含量采用酸性茚三酮显色法测定;丙二醛(MDA)含量采用硫代巴比妥酸法(TBA)的测定;超氧化物歧化酶(SOD)活性采用氮蓝四唑(NBT)还原法测定;过氧化物酶(POD)活性采用愈创木酚显色法测定。于测定当天上午8:00~9:00,选取各处理组相同部位功能叶数片进行,每个指标均重复测定3次。
1.4 数据处理
采用Excel 2003与DPS V6.55统计分析软件进行数据处理与分析。
2 结果与分析
2.1 不同强度干旱胁迫对刺槐叶片可溶性糖(SS)含量与游离脯氨酸(Pro)含量的影响
2.1.1 对可溶性糖(SS)含量的影响
如图1-A所示,随着干旱胁迫强度加大,刺槐叶片中可SS含量不断增加。其中LS(轻度干旱)条件下只比CK(对照)高出4.12%,与CK差异不显著;而MS(中度干旱)和HS(重度干旱)条件下SS含量增加明显,分别比CK高出74.23%、128.43%,与CK差异显著。
2.1.2 对游离脯氨酸(Pro)含量的影响
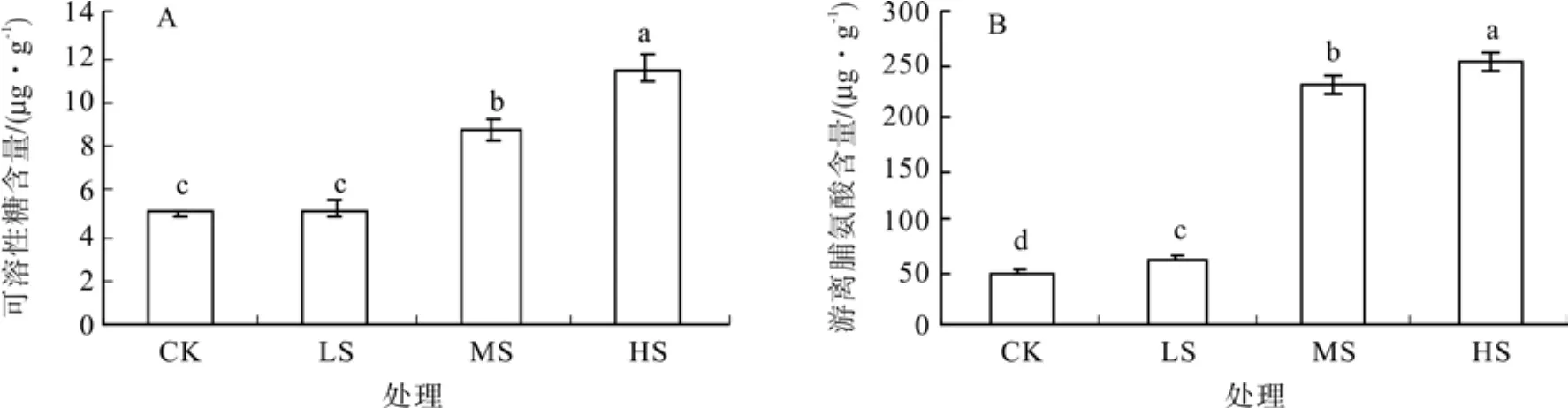
图1 不同强度干旱胁迫对可溶性糖含量与游离脯氨酸含量的影响Fig.1 Effects of different drought stress on soluble sugar content and free proline content
如图1-B所示,随着干旱胁迫强度的加大,刺槐叶片中Pro含量明显增加。HS条件下达到最高值252.47 ug/g,是CK的5.01倍,LS、MS时也分别达到CK的1.30倍、4.62倍;且CK、LS、MS、HS4种不同胁迫处理条件下两两之间Pro含量也都差异显著。
2.2 不同强度干旱胁迫对刺槐叶片相对电导率(REC)和丙二醛(MDA)含量的影响
2.2.1 对相对电导率(REC)的影响
由图2-A可知,随着干旱胁迫强度的加大,刺槐幼苗叶片REC值不断上升。其中,LS时REC稍高于CK但两者差异不显著,但在MS和HS条件下其值REC急剧增加,分别达到CK的3.06倍、4.04倍,均与CK差异显著。
2.2.2 对丙二醛(MDA)含量的影响
由图2-B可知,刺槐幼苗叶片的MDA含量表现为HS>MS>LS。其中LS条件下只比CK增加0.07 umol/g,两者差异不显著,而水分胁迫达到MS和HS条件时MDA的产生量分别是CK(对照)的2.44倍、4.39倍,与CK的差异显著。
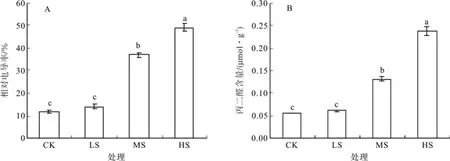
图2 不同强度干旱胁迫对相对电导率和丙二醛(MDA)含量的影响Fig.2 Effects of different drought stress on relative conductivity and malondialdehyde (MDA) content
2.3 不同强度干旱胁迫对刺槐叶片超氧物歧化酶(SOD)活性和过氧化物酶(POD)的影响
2.3.1 对超氧物歧化酶(SOD)活性的影响
由图3-A可看出,随着干旱胁迫强度的加大,刺槐幼苗叶片的SOD活性呈现出先不断增强后急剧减弱的变化特点。从CK到LS、MS其叶片的POD活性不断增强,且3种条件下任意两者之间差异均达到显著水平,随着干旱胁迫强度的进一步加大(HS条件下),SOD活性已经明显下降,但与CK相比,只低了1.72 u/g·min,两者差异不显著。
2.3.2 对过氧化物酶(POD)活性的影响
由图3-B可看出,随着干旱胁迫强度的加大,刺槐幼苗叶片的POD活性也呈现出先增强后减弱的变化特点,同时增强的速度与下降的速度均比较快。其中LS、MS时叶片的POD活性分别是CK的2.11倍、2.82倍,与CK差异明显;而HS与LS、MS相比,POD活性显著减弱而差异显著,但HS与CK相比,其活性仍然显著高于CK,达到CK的1.68倍,这说明在严重干旱胁迫条件下刺槐能够通过增强POD活性来应对干旱对其造成的伤害。

图3 不同强度干旱胁迫对超氧物歧化酶(SOD)和过氧化物酶(POD) 活性的影响Fig.3 Effects of different drought stress on superoxide dismutase (SOD) activity and peroxidase (POD)
3 结论与讨论
渗透调节是植物适应干旱胁迫的重要生理机制,而可溶性糖(SS)与脯氨酸(Pro)作为主要的渗透调节物质,大量研究表明,干旱胁迫条件下植物叶片SS和Pro含量通常会显著增加[6-7]。本研究也发现,随干旱胁迫强度的不断加大,刺槐叶片中SS和Pro含量不断增加,特别是在重度干旱(HS,15~20%的土壤相对含水量)胁迫时含量分别达到CK的2.28倍、5.01倍,这说明在严重干旱条件下,刺槐能够通过自身代谢活动诱导植物体内累积较多的SS与Pro来增加细胞溶质浓度,降低细胞渗透势,维持膨压,以增强植物的吸水保水能力,提高抗旱适应性与抗旱能力[8]。这与梁静等[9]研究酸枣叶对干旱胁迫的响应结果一致。
膜系统通常被认为是受干旱伤害的最初和关键部位[10],即当植物受到干旱胁迫时,往往首先表现细胞质膜受损而引起膜透性增大,导致细胞内电解质大量外渗。而相对电导率(REC)就是反映细胞膜透性的大小与膜系统稳定性的重要指标。REC值越大,表示细胞膜受损害的程度越重。而丙二醛( MDA)是细胞膜发生膜脂过氧化反应的产物之一,MDA含量的高低在一定程度上能反应脂膜过氧化的程度[11]。本研究中,REC与MDA含量的变化规律相类似,均表现为LS(轻度干旱)与对照差异不显著,而MS(中度干旱)和HS(重度干旱)均显著高于CK(对照),说明细胞膜透性变化与膜脂过氧化反应是缓慢渐进的,只有当干旱胁迫超出其能承受范围后才会引起细胞膜结构与功能受到严重伤害,导致膜透性增大与膜脂过氧化程度加重。
植物为保护自身免受伤害而形成了一套相应的抗氧化保护系统,其中超氧物歧化酶(SOD)和过氧化物酶(POD)在植物的抗氧化胁迫中起着极为重要的作用[12]。本研究中,随着干旱胁迫强度的加大,刺槐幼苗叶片的POD、SOD活性表现先增强后减弱,说明在LS(轻度干旱)、MS(中度干旱)胁迫下,刺槐叶片通过增强这两种保护酶活性来有效清除活性氧可能对其内部组织造成的伤害,但当干旱胁迫强度超出其自我适应限度,保护酶活性就会受到抑制而导致清除能力明显减弱[13],与刘球等[14]对红椿无性系幼苗研究结果一致。
综上所述,刺槐幼苗能够通过积累渗透调节物质可溶性糖SS与脯氨酸Pro、改变膜透性与MDA含量以及变化POD、SOD活性等生理生化特性来适应与应对不同强度干旱胁迫对其生命活动与生长造成的影响,表现出较强的抗旱耐旱潜力。
[1] 吕蒙蒙.刺槐丛枝菌根真菌提高土壤营养和结构稳定性的研究[D].西北农林科技大学,2013.
[2] 陈亚南,马露莎,张向茹,等.黄土高原刺槐叶片生态化学计量学特征[J].水土保持研究,2014,21(3):57-61,67.
[3] 赵丽英,杨建伟,张二芹.环境因子对盆栽刺槐苗木生理生长变化影响的通径分析[J].林业科学,2010,46(4):140-145.
[4] 徐发良.高温胁迫对刺槐幼苗生理指标的影响[J].山西林业,2014, (1):30-31.
[5] 高俊凤.植物生理学实验指导[M].北京:高等教育出版社,2006.
[6] 董 蕾,陈 博,李吉跃,等.5个楸树无性系抗旱性的综合评价[J].中南林业科技大学学报,2014,34(4):30-35.
[7] 潘 昕,谢德新,邱 权,等.干旱胁迫对两种速生树种苗木生理指标的影响[J].中南林业科技大学学报,2013,33(10):84-88.
[8] 阎秀峰,李 晶,祖元刚.干旱胁迫对红松幼苗保护酶活性及脂质过氧化作用的影响[J].生态学报,1999,19(6):850-854.
[9] 梁 静,魏学智.酸枣叶对土壤水分的生理生化响应[J].植物研究,2010,30(5):549-555.
[10] 杨鑫光,傅 华,李晓东.干旱胁迫对霸王水分生理特征及细胞膜透性的影响[J].西北植物学报, 2009, 29(10):2076-2083.
[11] 包 卓,孟祥英,张晓松,等.干旱胁迫对5种园林绿化植物生理生化的影响[J].经济林研究,2010,28(3):46-50.
[12] 谷文众,刘 杨,谷振军.干旱胁迫对金盏菊膜脂过氧化及保护酶活性的影响[J].经济林研究,2009,27(3):79-81.
[13] 肖姣娣.火棘幼苗生理生化特性对干旱胁迫的响应[J].东北林业大学学报,2014,42(11):14-16,27.
[14] 刘 球,陈彩霞,吴际友,等.红椿无性系幼苗叶片抗氧化酶指标对干旱胁迫的响应[J].中南林业科技大学学报,2013,33(13): 73-76,82.
猜你喜欢
中老年保健(2022年3期)2022-11-21
安徽化工(2021年3期)2021-05-29
现代园艺(2020年2期)2020-12-22
棉花学报(2020年3期)2020-08-08
音乐研究(2019年5期)2019-11-22
科学与财富(2019年15期)2019-10-21
湖南饲料(2019年4期)2019-10-17
江苏农业学报(2019年1期)2019-09-10
小溪流(故事作文)(2018年10期)2018-12-28
现代园艺(2018年2期)2018-03-15
