
基于单片段代换系的水稻抽穗期QTL上位性互作分析
2015-05-06 14:58刘书旖等
山东农业科学 2015年3期
关键词:水稻
刘书旖等


摘要:抽穗期是水稻(Oryza sativa)重要的农艺性状,它决定着水稻品种的地区和季节适应性。本试验利用以华粳籼74为受体亲本发展的带有抽穗期基因的15个单片段代换系为试验材料,通过两两杂交,筛选出15个不同亲本组合的双片段聚合系,结合分子标记辅助选择进行水稻抽穗期QTL鉴定,并研究QTL的聚合及其互作关系。
关键词:水稻;抽穗期基因;单片段代换系;基因聚合系;互作分析
中图分类号:S511.035.3文献标识号:A文章编号:1001-4942(2015)03-0001-05
Epistasis Interaction Analysis of QTL for Heading
Date in Rice using Single Segment Substitution Lines
Liu Shuyi1,Zhang Hua2,Liu Xu2,Xuan Ning2,Yang Yongyi2,Li Jun2,Li Guangxian2,Yao Fangyin2*
(1.Shandong Normal University,Jinan 250014,China;
2. Bio-Tech Research Center,Shandong Academy of Agricultural Sciences,Jinan 250100,China)
AbstractHeading date is an important agronomic trait in rice(Oryza sativa),which decides the variety adaptability of cultivation areas and seasons. Taking 15 single segment substitution lines which developed from Huajingxian 74 and carryed heading date gene as materials,after crossing them in pairs,15 double segment pyramiding lines were selected to identify the quantitative trait loci(QTL) for heading date by marker-assisted selections,and the pyramiding and interacting effects of QTL were also studied.
Key wordsRice;Heading date gene;Single segment substitution lines;Genic pyramiding lines;Interaction analysis
抽穗期(Heading date, HD)决定着水稻品种的地区和季节适应性,是水稻最重要的农艺性状之一,对品种的高产稳定起着不可忽视的作用。水稻早熟基因的发现不仅解决了早熟与丰产难以兼顾的问题,而且对克服籼粳亚种间F超亲迟熟有着重要作用[1]。因此,对水稻抽穗期基因定位、克隆、互作分析等方面的研究,对于提高农业生产具有重要的理论意义和应用价值。抽穗期长短主要由品种的感光性(Photoperiod sensitivity, PS)、感温性(Temperature sensitivity, TS)和基本营养生长性(Basic vegetative growth, BVG)决定[2]。抽穗期的遗传由主效基因和微效基因共同控制,分析方法以分子标记和统计分析方法为基础[3]。近年来,水稻抽穗期QTL定位、克隆等研究取得了重大进展,在多个群体中定位了大量水稻抽穗期QTL[4]。目前,国内外定位了700多个抽穗期QTL(http//www.gramene.org/qtl/index.html),分布在水稻12条染色体上,其中第3、6、7染色体上较多,尤其是第3染色体上共定位了102个QTLs,而第10染色体上最少,只有22个。存在于不同位点上的QTL被检测到的概率取决于自身的效应大小,效应较大的QTL被检测到的概率较大[5]。
染色体单片段代换系(Single segment substitution lines, SSSLs)是采用连续回交的方法,结合分子标记辅助选择,将供体亲本的染色体片段导入轮回亲本建立起来的一套近等基因系。单片段代换系只有代换片段与受体亲本不同,其它遗传背景与受体亲本完全一致,对代换区段中的QTL进行分析时遗传背景干扰很小,因此,单片段代换系是进行上位性研究理想的试验材料。不少学者利用单片段代换系材料对QTL进行了鉴定和精细定位,并克隆了一些重要性状的QTL[6]。水稻抽穗期受主效基因与微效多基因控制,基因之间存在互作[7~9]。以染色体单片段代换系为试验材料,通过两两杂交与连续自交,构建双片段纯合体株系,可以进行水稻抽穗期QTL的上位性互作分析。
1材料与方法
1.1试验材料
本试验所用15个单片段代换系均以华粳籼74为受体亲本,代换片段来源于不同的供体,具体情况见表1。
1.2试验方法
1.2.1双片段纯合体筛选利用不同的单片段代换系配制杂交组合,将选育出的杂合双片段聚合系在自然条件下进行自交。根据华粳籼74与单片段代换系所在区间的引物多态性,进行SSR检测,根据基因型,筛选出已聚合为双片段纯合体的株系,为下一步试验做准备。
1.2.2双片段纯合体代换片段长度的检测采集筛选出的双片段纯合体嫩叶,采用TPS粗提法提取DNA。用代换片段区间上的多态性引物,以两亲本与华粳籼74为对照,检测双片段纯合体中两个代换片段的长度。endprint
1.3田间试验
将华粳籼74、选出的双片段纯合体及其亲本于2014年4月28日播种,6月1日插秧。6月下旬开始进行抽穗期调查,记录单株抽穗期,以每株主穗抽出1 cm作为该株始穗的标准,以播种到始穗的天数作为抽穗期,每隔2~3天调查一次。
2结果与分析
2.1QTL的鉴定
根据抽穗期的数据可以确定单片段代换系上QTL的存在。若t检验得出的阈值P≤10-6,则代换系抽穗期与华粳籼74存在显著差异,认为单片段代换系上有不同于华粳籼74效应的QTL存在[10]。
加性效应值=(纯合SSSL的表型值-对照的表型值)/2
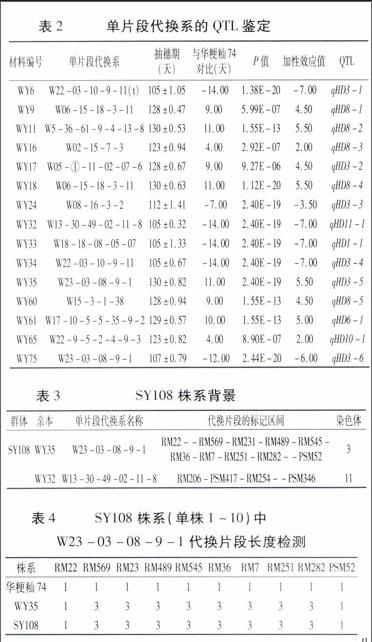
通过t检测可知,15个单片段代换系上均存在有不同于华粳籼74效应的QTL,并分别将其命名(表2)。
2.2双片段纯合体的筛选
2013年,通过对300个不同亲本组合的双片段杂合体株系自交获得的后代进行分子检测,共筛选出20个不同亲本组合的双片段纯合体株系。2014年种下筛选出的20个双片段纯合体株系,每个株系包括40个单株,取叶后提取DNA,对双片段纯合体中的两个代换片段长度进行检测。
2.3双片段纯合体代换片段长度的检测
利用代换片段内具有多态性的引物,对20个不同亲本组合的双片段纯合体代换片段长度进行检测。以2014年群体SY108为例,结果见表3。
由试验可得,SY108群体单株1~10含有与亲本WY35长度一致的代换片段,同理可验证,单株1~10含有与亲本WY32长度一致的代换片段。两个代换片段均与亲本代换片段长度相同的双片段纯合体株系共15个。
2.4QTL的互作分析
对筛选出的15个双片段纯合体株系进行抽穗期QTL互作分析。华粳籼74的抽穗期天数为119±1.01。
株系SY004(WY6/WY9聚合系)抽穗天数是119±0.67,与华粳籼74相当,而W22-03-10-9-11(t)上的QTL对其产生负加性效应,W06-15-18-3-11上的QTL对其产生正加性效应,W22-03-10-9-11(t)表现为早熟,W06-15-18-3-11表现为较晚熟,因此,qHD8-1对qHD3-1表现上位性。
株系SY016(WY11/WY17聚合系)抽穗天数是123±0.44,比华粳籼74晚抽穗,而W05-①-11-02-07-6和W5-36-61-9-4-13-8上的QTL均对其产生正加性效应,W05-①-11-02-07-6表现为较晚熟,W5-36-61-9-4-13-8表现为晚熟,因此,qHD3-2对qHD8-2表现上位性。
株系SY027(WY17/WY18聚合系)抽穗天数是127±0.38,比华粳籼74晚抽穗,W06-15-18-3-11和W05-①-11-02-07-6上的QTL均对其产生正加性效应,W06-15-18-3-11表现为晚熟,W05-①-11-02-07-6表现为较晚熟,因此,qHD3-2对qHD8-4表现上位性。
株系SY051(WY16/WY24聚合系)抽穗天数是112±0.89,比华粳籼74早抽穗,而W02-15-7-3上的QTL对其产生正加性效应,W08-16-3-2上的QTL对其产生负加性效应,说明qHD3-3对qHD8-3表现上位性。
株系SY053(WY18/WY24聚合系)抽穗天数是115±0.56,比华粳籼74早抽穗,而W06-15-18-3-11上的QTL对其产生正加性效应,W08-16-3-2上的QTL对其产生负加性效应,说明qHD3-3对qHD8-4表现上位性。
株系SY062(WY24/WY33聚合系)的抽穗天数是95±1.02,比华粳籼74早抽穗,而W08-16-3-2和W18-18-08-05-07上的QTL均对其产生负加性效应,W08-16-3-2表现为较早熟,W18-18-08-05-07表现为早熟,因此,qHD1-1对qHD3-3表现上位性。
株系SY069(WY24/WY34聚合系)抽穗天数是95±0.89,比华粳籼74早抽穗,而W08-16-3-2和W22-03-10-9-11上的QTL均对其产生负加性效应,W08-16-3-2表现为较早熟,W22-03-10-9-11表现为早熟,因此,qHD3-4对qHD3-3表现上位性。
株系SY070(WY24/WY65聚合系)抽穗天数是109±0.69,比华粳籼74早抽穗,而W08-16-3-2上的QTL对其产生负加性效应,W22-9-5-2-4-9-3上的QTL对其产生正加性效应,说明qHD3-3对qHD10-1表现上位性。
株系SY083(WY16/WY32聚合系)抽穗期天数是110±0.87,比华粳籼74早抽穗,而W02-15-7-3上的QTL对其产生正加性效应,W13-30-49-02-11-8上的QTL对其产生负加性效应,说明qHD11-1对qHD8-3表现上位性。
株系SY092(WY6/WY33聚合系)抽穗期天数是105±1.05,比华粳籼74早抽穗,而W22-03-10-9-11(t)与W18-18-08-05-07上的QTL均对其产生负加性效应,两亲本与聚合系抽穗期天数相同,因此,qHD3-1与qHD1-1没有叠加效应。
株系SY097(WY24/WY33聚合系)的抽穗天数是95±0.89,比华粳籼74早抽穗,而W08-16-3-2和W18-18-08-05-07上的QTL均对其产生负加性效应,W08-16-3-2表现为较早熟,W18-18-08-05-07表现为早熟,因此,qHD1-1对qHD3-3表现上位性。
株系SY106(WY34/WY60聚合系)抽穗期天数是105±1.21,比华粳籼74早抽穗,而W22-03-10-9-11上的QTL对其产生负加性效应,W15-3-1-38上的QTL对其产生正加性效应,说明qHD3-4对qHD8-5表现上位性。endprint
株系SY108(WY32/WY35聚合系)抽穗期天数是95±0.98,比华粳籼74早抽穗,而W13-30-49-02-11-8上的QTL对其产生负加性效应,W23-03-08-9-1上的QTL对其产生正加性效应,说明qHD11-1对qHD3-5表现上位性。
株系SY118(WY35/WY61聚合系)抽穗天数是132±0.36,比华粳籼74晚抽穗,而W23-03-08-9-1和W17-10-5-5-35-9-2上的QTL均对其产生正加性效应,W23-03-08-9-1和W17-10-5-5-35-9-2均表现为晚熟,聚合系表现为更晚熟,因此,qHD3-5和qHD6-1对抽穗期有叠加效果。
株系SY137(WY18/WY75聚合系)抽穗期天数是107±0.57,比华粳籼74早抽穗,而W06-15-18-3-11上的QTL对其产生正加性效应,W23-03-08-9-1上的QTL对其产生负加性效应,说明qHD3-6对qHD8-4表现上位性。
3结论
水稻具有高效的转化系统和高密度的遗传图谱和物理图谱[11,12]。本试验以田间调查数据与实验室分子标记为基础,从300组双片段杂合体株系中筛选出20个不同亲本组合的双片段纯合体,并对双片段纯合体中两个代换片段长度进行检测,与亲本代换片段长度相同的有15个。经上位性分析,得出双片段纯合体中两个相应QTL的上位性互作关系。该试验通过对水稻抽穗期QTL的互作分析,为其精细定位、分子标记辅助选择育种、基因克隆和转移、数量性状有利基因聚合等奠定基础。
致谢:本试验是在山东省农业科学院生物技术研究中心姚方印研究员实验室完成的。首先感谢姚方印老师、李广贤博士、张华博士、杨永义博士、柳絮助理研究员的悉心指导,同时感谢山东师范大学的支持,以及实验室同学们的关心帮助。
参考文献:
[1]
董春林, 孙业盈, 邓晓建. 水稻抽穗期基因研究进展[J]. 中国农学通报, 2005, 21(6): 459-462.
[2]魏祥进, 江玲, 徐俊锋, 等. 我国华北地区粳稻品种抽穗期遗传分析[J]. 中国水稻科学, 2009, 23(6): 596-600.
[3]邵迪, 李秋萍, 吴比, 等. 利用染色体片段代换系定位水稻主效抽穗期QTL[J]. 湖南农业大学学报, 2009, 35(4): 344-347.
[4]魏祥进, 徐俊峰, 江玲, 等. 我国水稻主栽品种抽穗期多样性的遗传分析[J]. 作物学报, 2012, 38(1): 10-22.
[5]Yano M, Sasaki T. Genetic and molecular dissection of quantitative traits in rice [J]. Plant Mol. Biol., 1997, 35: 145-153.
[6]张丽霞, 刘丕庆, 刘学义, 等. 染色体单片段代换系的构建及应用[J]. 分子植物育种, 2004, 2(3): 743-746.
[7]蔡俊迈, 周元昌, 李维明. 抽穗期基因间互作的遗传[J]. 福建农学院学报, 1988, 17(1): 1-9.
[8]Lin H X, Yamamoto T, Sasaki T, et al. Characterization and detection of epistatic interaction of 3 QTLs, Hd1, Hd2, and Hd3, controlling heading date in rice using nearly isogenic lines[J]. Theor. Appl. Genet., 2000, 101(1): 1021-1028.
[9]Yano M, Katayose Y, Ashikari M, et al. Hd1, a major photoperiod sensitivity quantitative trait locus in rice, is closely related to the Arabidopsis flowering time gene CONSTANTS[J]. The Plant Cell, 2000, 12(1): 2473-2483.
[10]姚方印. 基于SSSL的水稻抽穗期QTL的鉴定及上位性分析[D]. 广州:华南农业大学, 2006.
[11]Umehara Y, Inagaki A, Tanoue H, et al. Construction and characterization of a rice YAC library for physical mapping[J]. Mol. Breed., 1995, 1(1): 79-89.
[12]Harushima Y, Yano M, Shomura A, et al. A high-density rice genetic linkage map with 2275 markers using a single F2 population[J]. Genetics, 1998, 148(1): 479-494.endprint
猜你喜欢
世界热带农业信息(2021年7期)2021-08-11
格言·校园版(2021年30期)2021-07-06
好孩子画报(2021年3期)2021-04-02
农民致富之友(2020年8期)2020-05-11
农民致富之友(2020年1期)2020-02-11
农民致富之友(2019年30期)2019-10-21
农民致富之友(2019年23期)2019-08-16
农民致富之友(2019年14期)2019-05-24
新农村(2018年29期)2018-04-02
农产品市场周刊(2017年35期)2017-10-25
