
生态技术创新视角下工业产业的种群效应实证研究
2015-04-20 01:26万春
软科学 2015年3期
万春

摘要:以2001~2011年江西省的工业产业为样本,采用生态技术创新种群行为模型对种群效应进行实证分析,发现多数种群的个体数量已经逼近或超过区域创新资源决定的生态阈值,种群内部、种群之间由协同效应转向竞争效应,发出了环境过载的警示。
关键词:生态技术创新;工业产业;种群效应
DOI:10.13956/j.ss.1001-8409.2015.03.10
中图分类号:F423.3;F4036 文献标识码:A文章编号:1001-8409(2015)03-0043-04
1引言
在技术创新驱动下,工业化空前提高了人类的生产力,但对生态环境造成了严重破坏。我国作为最大的发展中国家,工业化是不可避免的一个发展阶段,这就决定了富有生态属性的新型工业化是我国生态经济的基本内核。党的十七大和十七届三中、四中全会将转变增长方式、推进生态经济提到了国家战略的高度,而新型工业化是最重要的措施之一。2012年11月,党的十八大报告强调了以“实施创新驱动发展战略”为支撑,“坚持走中国特色新型工业化道路”。同时,欧美等国家的再工业化浪潮使国际产业竞争聚焦于生态技术创新。从而,研究工业产业的生态技术创新就显得非常必要。
技术创新理论由Jeseph Schumpeter于1912年提出后,成为国外研究的热点之一,但主要是追求经济效益的传统技术创新,虽可大幅度提升生产力,亦加剧了环境污染及资源消耗,致使技术创新的增长效应递减,面临增长极限[1,2]。解决的根本途径是将生态元素融入技术创新,依靠生态技术创新取得技术进步,1950~2003年间中亚国家新型工业化的重要经验就是籍此壮大产业[3,4]。在不同层次、规模上,生态技术创新的目标、影响不同,从而内在机理也不同[5]。应根据国家或地区的特性,从政治、经济及文化等多个维度协同推进,结合企业组织、制度的变革来实施生态技术创新[6,9]。
生态技术创新的特征包括:创造性,将新技术、新工艺应用于生产,实施组织、制度、管理上的创新;生态性,将生态因素融入生产的全部环节 [10,11]。生态技术创新受到技术创新、人类、环境等多种因素及相互关系的影响[12,13]。企业拥有的创新信息资源形成特定的信息生态位——包括信息识别能力、获取能力、整合能力等生态因子,使企业的技术创新能力不同[14]。企业与合作伙伴形成技术创新网络,这是类似生命体的“活性组织”[15]。生态技术创新的进化机理可归结为生态共生、生态平衡、生态重组三类 [16]。从本质上讲,技术创新系统作为社会生态大系统中的子系统,将生态价值嵌入技术创新,使技术创新在生态领域实现价值回归[17]。综合现有研究的观点,生态技术创新是指创新主体为了获取生态经济综合效益,遵循自然生态系统的运行规律,将生态环保理念融入经济活动链上的各环节,消除污染,促进物质、能量等的循环而实施的技术创新。
从国内外的研究来看:将生态学原理移植运用于经济领域探索生态技术创新是一种趋势,但由于涉及生态、经济、技术等复杂因素,目前尚缺乏成熟的研究,且现有研究主要关注单个主体的创新行为,而事实上这是集体性的种群行为。本文依据以前的研究成果,运用生态技术创新种群行为模型,以江西省工业产业为样本,通过计量实证方法,研究工业产业生态技术创新的种群效应,从中得到实施生态技术创新的启示。
2模型构建
生态学中,种群是指一定时间空间范围内同种生物个体的集合。种群内部以及种群之间存在密切联系:既相互依存于共同的环境,又对环境容量决定的资源展开争夺。将经济系统视为生态子系统,产业即为种群,企业即为种群个体。黄鲁成根据生态学从Logistic模型推导Votka-Volterra模型的思路,建立了技术创新种群行为模型[18]。本文在此基础上,放宽相关假设,构建种群个体数量的临界值指标,拓展生态技术创新种群间关系模型。假定:
(1)种群个体的均衡数量由边际法则决定。随着个体数量N增加,边际收益MR(N)递减,而边际成本MC(N)由于资源紧缺、价格上涨而递增。MR(N)=MC(N)决定最大均衡个体数量N*0=K。
(2)给定生态技术创新资源(简称“创新资源”),当种群个体数量为N时,平均占据创新资源N/K,个体数量增长率g与剩余创新资源(1-N/K) 成正比关系。N由1→K,每增加1个单位,创新资源平均消耗1/K。
21单一生态技术创新种群的内部关系模型
在时间t期,以gt表示种群增长率,N(t)表示种群个体数量,α为与t-1期种群个体数量N(t-1)有关的增长率系数,根据生物种群的Logistic模型,有:
gt=dN(t)dt=αN(t-1){1-N(t-1)K} (1)
记dN(t)≈ΔN(t)=N(t)-N(t-1),dt≈Δt=t-(t-1)=1,有:
ΔN(t)==ρ1N(t-1)+ρ2N(t-1)2(2)
其中,ΔN(t)表示t期种群个体数量的增量;ρ1=α,一般ρ1>0,表征种群内部的协同效应;ρ2=-α/K,称为密度抑制系数或内部竞争系数,通常ρ2<0,表征种群内部的竞争效应。有:
ΔN(t)={ρ1+ρ2N(t-1)}N(t-1)(3)
(1){ρ1+ρ2N(t-1)}>0,有ΔN(t)>0。种群内部以协同效应为主,个体数量在创新资源所能支撑的最大数量之内,可维持增长趋势。①ρ1>0,ρ2>0;ρ1=0,ρ2>0;ρ1>0,ρ2=0。种群内部只有协同效应,还未产生竞争效应。②ρ1>0,ρ2<0,且ρ1>ρ2N(t-1);ρ1<0,ρ2>0,且ρ1<ρ2N(t-1)。种群内部已产生竞争效应,但比协同效应弱。
(2){ρ1+ρ2N(t-1)}<0,有ΔN(t)<0。种群内部以竞争效应为主,个体数量已超过创新资源可支撑的最大数量,在竞争淘汰中减少。ρ1<0,ρ2<0。种群内部为竞争效应,不存在协同效应。ρ1>0,ρ2<0,且ρ1<ρ2N(t-1);ρ1<0,ρ2>0,且ρ1>ρ2N(t-1)。种群内部竞争效应大于协同效应。
构造种群个体数量临界值指标N*=ρ1/ρ2,判别种群的内部关系:当个体数量少于N*,协同效应大于竞争效应;反之,竞争效应大于协同效应。
22两个不同生态技术创新种群之间的关系模型
对于两个生态技术创新种群1、2,它们各自类似而又独立的Logistic变化可用式(1)表示。基于Logistic模型,Lotka和Volterra提出了Lotka-Volterra种群间竞争模型,被广泛应用于现代生态学理论研究。基本思想是种群1、2对彼此的增长率产生影响,表现为:
N1 (t)=β12 N2 (t)(4)
N2 (t)=β21 N1 (t)(5)
其中,N1(t)、N2(t)为种群1、2在t期的个体数量。β12 为种群2对种群1的影响系数,指对于种群1给定的创新资源,一个种群2个体与β12 个种群1个体占用的资源相当:种群2增加1个单位,种群1会减少β12 个单位。类似地,β21 为种群1对种群2的影响系数。以K1、K2表示种群1、2在各自创新资源给定条件下的最大数量,t-1期种群1、2的个体数量为N1(t-1)、N2(t-1),占用种群1的创新资源分别为N1(t-1)/K1、β12 N2 (t-1)/K1 ,则种群1的Logistic模型可修正为:
dN1 (t)dt=α1 N1 (t-1)(1-N1 (t-1)K1 -β12 N2 (t-1)K1 )(6)
将式(4)代入作差分处理,得到:
ΔN1(t)=γ1N2(t-1)+γ2N2(t-1)2(7)
其中,γ1 =α1 β12 ,γ2=-2α1β212/K1。将式(7)变形为:
ΔN1(t)={γ1+γ2N2(t-1)}N2(t-1)(8)
类似前面分析,可以根据γ1、γ2的取值,判断种群1、2之间的关系。
(1){γ1+γ2N2(t-1)}>0,有ΔN1(t)>0。种群1、2之间以协同效应为主,种群2增加引起种群1增加。①γ1>0,γ2>0;γ1=0,γ2>0;γ1>0,γ2=0。种群1、2之间为协同效应,不存在竞争效应。②γ1>0,γ2<0,且γ1>γ2N2(t-1);γ1<0,γ2>0,且γ1<γ2N2(t-1)。种群1、2之间的协同效应大于竞争效应。
(2){γ1+γ2N2(t-1)}<0,有ΔN1(t)<0。种群1、2之间以竞争效应为主,二者为替代关系,种群2增加引起种群1减少。①γ1<0,γ2<0。种群1、2之间为竞争效应,不存在协同效应。②γ1>0,γ2<0,且γ1<γ2N2(t-1);γ1<0,γ2>0,且γ1>γ2N2(t-1)。种群1、2之间的竞争效应大于协同效应。
构造种群间个体数量临界值指标N*2=γ1/γ2,判别种群之间的关系:当种群2的个体数量超过临界值后,种群1、2之间的关系发生改变。
3计量模型与样本选取
31计量模型
由式(2)和式(7),分别构建计量模型,检验种群内部、种群之间的关系。
ΔN(t)=ρ0+ρ1N(t-1)+ρ2N(t-1)2+ε0(t) (9)
ΔN1(t)=γ0+γ1N2(t-1)+γ2N2(t-1)2+ε1(t)(10)
其中,ρ0、ρ1、ρ2、γ0、γ1、γ2为待估参数,ε0(t)、ε1(t)为残差项。
32样本选取
由于产业基础、资源禀赋的区域差异较大,我国尚未形成推进生态技术创新的统一模式,应从典型地区选择样本。江西省生态资源丰裕,持续实施了打造昌九工业走廊、壮大十大战略性新兴产业等发展战略,且《鄱阳湖生态经济区规划》于2009年获得国务院批复,建立了探索生态经济、实施生态技术创新、走新型工业化道路的国家级战略平台。考虑到规模以上工业企业的创新能力相对较强,直接关系到创新型国家目标的实现[19],故选取江西省2001~2011年间35个工业产业中规模以上企业为样本。按性质相近的原则分类,得到13个归并工业产业,视为13个种群,分别是采选N1、食品加工制造N2、纺织服装N3、木材及家具制造N4、印刷文体制造N5、石油化工N6、医药制造N7、非金属制品N8、金属制品N9、设备制造N10、电子N11、回收加工N12、能源及水的生产与供应N13。数据来源于历年《江西统计年鉴》,本文作了测算。
4实证分析
41单一生态技术创新种群内部关系的计量估计
按式(9),运用Eviews60软件进行计量处理,估计13个种群的ρ1、ρ2,详见表1。种群的内部关系有以下特点:
(1)采选N1、木材及家具加工制造N4、能源及水生产与供应N13等的内部为协同效应大于竞争效应,个体数量未达到临界值。原因在于:江西省的矿产、林业资源丰富,环境承载力强,种群N1、N4内部实施生态技术创新协作有较大空间;种群N13公共属性强、垄断程度高,个体数量有限,内部竞争效应受到制约。
(2)石油化工N6、医药制造N7的协同效应大于竞争效应,但部分年份个体数量超过临界值,种群内部关系逆转。N6、N7增长较快,2009年和2010年个体数量超过临界值,在竞争效应的淘汰机制下,2011年恢复至临界值以下。
(3)纺织服装N3前期协同效应大于竞争效应,后期则相反。N3属于行业竞争性强、准入门槛低的种群,个体数量增长惯性大,由2001年的217上升到2010年的961。但从2004年起,种群内部就以竞争效应为主,直至2011年,回调作用才显现,下降到777。
(4)食品加工制造N2、印刷文体制造N5、设备制造N10的竞争效应大于协同效应。从2001年开始,这些种群个体数量就已超过临界值,竞争效应明显,但在投资饥渴、市场容量扩大等因素的综合作用下,直到2011年才开始减少。
(5)非金属制品N8、金属制品N9、电子N11内部存在竞争效应。这些产业为典型的竞争性种群,个体数量分别从2001年的77、472、246,快速增加到2010年的1202、856、637,2011年开始回落。
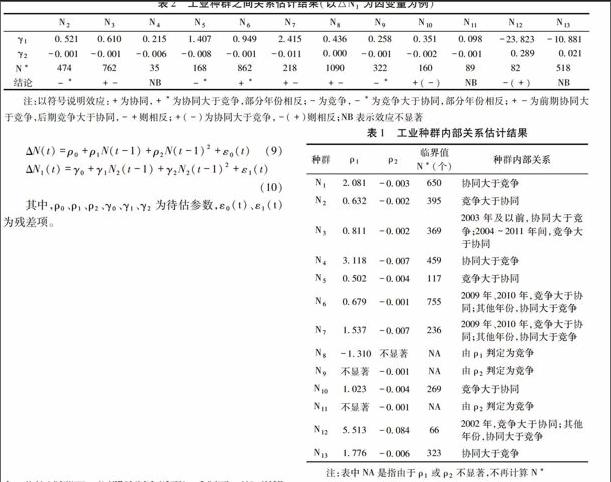
表1工业种群内部关系估计结果
种群ρ1ρ2临界值
N*(个)种群内部关系N12081-0003650协同大于竞争N20632-0002395竞争大于协同N30811-00023692003年及以前,协同大于竞争;2004~2011年间,竞争大于协同N43118-0007459协同大于竞争N50502-0004117竞争大于协同N60679-00017552009年、2010年,竞争大于协同;其他年份,协同大于竞争N71537-00072362009年、2010年,竞争大于协同;其他年份,协同大于竞争N8-1310不显著NA由ρ1判定为竞争N9不显著-0001NA由ρ2判定为竞争N101023-0004269竞争大于协同N11不显著-0001NA由ρ2判定为竞争N125513-0084662002年,竞争大于协同;其他年份,协同大于竞争N131776-0006323协同大于竞争注:表中NA是指由于ρ1或ρ2不显著,不再计算N*
42不同生态技术创新种群之间关系的计量估计
按式(10),估计13个种群的γ1、γ2。表2给出了△N1为因变量的处理结果。种群之间的关系有以下特点:
(1)协同效应与产业链衔接紧密度正相关,种群之间通过技术共生、产业互补等途径协作发展,产业链衔接越紧密,协同效应越强。采选N1与石油化工N6、设备制造N10之间,协同效应显著。
(2)竞争效应与种群个体数量正相关,种群个体数量越多,对资金、人才等的争夺越激烈,竞争效应越强。采选N1与食品加工制造N2之间,N2与纺织服装N3之间,均为竞争效应。
(3)当产业链衔接紧密度、种群个体数量等发生变化,种群之间的效应随之而变,通常前期以协同效应为主,后期以竞争效应为主。采选N1与纺织服装N3之间,印刷文体制造N5与医药制造N7之间,均呈现这种规律。
(4)当个体数量未超过临界值,由于跨种群生态技术创新水平不高,资源综合利用率低,产业链衔接紧密度不高的种群间的效应不显著。采选N1与木材及家具加工制造N4之间,食品加工制造N2与石油化工N6之间,效应均不显著。
(5)能源及水的生产与供应N13对其他种群具有支持作用,存在协同效应;但支持作用不足,表现出效应不显著的一面。N13与食品加工制造N2之间,协同效应大于竞争效应;与纺织服装N3之间,效应不显著。表2工业种群之间关系估计结果(以△N1为因变量为例)
N2N3N4N5N6N7N8N9N10N11N12N13γ10521 0610 0215 1407 0949 2415 0436 0258 0351 0098 -23823 -10881 γ2-0001 -0001-0006 -0008 -0001 -0011 0000 -0001 -0002 -0001 0289 0021N*474 762 35 168 862 218 1090 322 160 89 82 518 结论-*+-NB-*+*+-+--*+(-)NB-(+)NB注:以符号说明效应:+为协同,+*为协同大于竞争,部分年份相反;-为竞争,-*为竞争大于协同,部分年份相反;+-为前期协同大于竞争,后期竞争大于协同,-+则相反;+(-)为协同大于竞争,-(+)则相反;NB表示效应不显著5结论与启示
本文运用生态技术创新种群行为模型,基于江西省工业种群进行实证分析,得到如下研究结论:2001~2011年间,作为个体的工业企业数量快速增长,使种群内部、种群之间的关系呈现协同效应下降而竞争效应上升的趋势,反映了生态环境对粗放增长方式所产生的资源浪费、环境承载力透支等问题的警示。由于创新资源有限,种群内部关系直接取决于个体数量,很多种群的个体数量超过或逼近临界值,但竞争效应通过市场淘汰机制,使过多的个体趋向均衡,效果却于2011年才开始显现。种群之间的关系不仅取决于个体数量,还与种群之间的产业链衔接紧密度有关。一方面,江西省的工业种群个体数量普遍增加,种群间对区域创新资源的争夺使竞争效应上升;另一方面,生态技术创新相对滞后,产业契合度不高,协同效应不足。
本文揭示了生态技术创新种群的竞争、协同效应,其实际贡献在于,提供了实施生态技术创新的有益启示:①将仅注重单一创新主体创新行为的传统视野,拓展到区域资源决定的种群的更为宽广的范围;②提升生态技术创新水平,增进种群的协同效应。改进工艺,提高废弃物利用率,增强种群内的产业契合度,发展循环经济,开发循环链路,拓展互补共生渠道,强化耦合机制,提升种群间的产业紧密度;③调控个体数量,科学限制种群的竞争效应,提高生态准入门槛,改变当前盛行的盲目上马项目、片面追求依靠增加个体数量来提升种群集聚度的做法,科学配置创新资源,优化种群个体数量,使之趋于合理的均衡水平。
此外,本文的研究结论对政府实施公共投资形成公共种群的实践也具有较好的启迪意义:完善交通、电力等公共种群的发展模式,合理规划空间布局,增进与其他种群的协同效应,提升基础支持作用。
本文仅从个体数量来分析工业产业的生态技术创新种群效应,未涉及产业属性、技术耦合程度等因素,这既是本文的不足,也是进一步研究的方向。
参考文献:
[1]Carson R.Silent Spring [M].New York:Fawcett Crest Book,19621-15.
[2]Meadows DH,Meadows DL,Randers J,Behrens IIIWW.The Limits to Growth [M].New South Wales:Universe Books,197220-71.
[3]Sudakov VB.Ecological Aspects of Modern Technologies in Dam Construction [J].Hydrotechnical Construction,1994,28(11):635-636.
[4]STORM S,Naastepad CWM.Strategic Factors in Economic Development:East Asian Industrialization 1950-2003 [J].Development and Change,2005, 36(6):1059-1094.
[5]Machiba T.EcoInnovation for Enabling Resource Efficiency and Green Growth:Development of an Analytical Framework and Preliminary Analysis of Industry and Policy Practices [J].Int Econ Econ Policy,2010(7):357-370.
[6]Arthur PJM.Ecological Modernization and the Global Economy [J].Global Environmental Politics,2002,2(2):92-115.
[7]Johnstone N,Hascic I.Ecoinnovation,Policy and Globalization [R].OECD Observer,2007264-265.
[8]Ekins P.Ecoinnovation for Environmental Sustainability:Concepts,Progress and Policies [J].Int Econ Econ Policy,2010(7):267-290.
[9]Laperche B,Uzunidis D.EcoInnovation,Knowledge Capital and the Evolution of the Firm [J].The IUP Journal of Knowledge Management,2012(3):14-35.
[10]蔡宁,吕燕.产品导向型生态技术创新过程及其特征的研究 [J].生态经济,2000(12):16-19.
[11]荣诚.生态技术创新研究初探 [J].中国软科学,2004(5):159-160.
[12]吕玉辉,丁长青.技术创新生态与生态技术创新及其他 [J].科技管理研究,2007(3):33-34.
[13]薛桂波.基于软环境视角的生态技术创新分析 [J].科学管理研究,2010,28(3):47-50.
[14]陈浩义,王文彦.基于信息生态位视角对企业技术创新能力的认识 [J].科技进步与对策,2011,28(10):76-79.
[15]刘晓燕,阮平南,李非凡.基于关键成功因素的技术创新网络动态伙伴选择模型 [J].软科学,2014,28(7):25-28.
[16]汪志波.产业技术创新生态系统演化机理研究 [J].生产力研究,2012(3):192-194.
[17]易显飞.西方技术创新生态价值取向的历史变迁——从思维方式的视角看 [J].山西大学学报(哲学社会科学版),2013,36(2):13-17.
[18]黄鲁成.基于生态学的技术创新行为研究 [M].北京:科学出版社,200778-87.
[19]高霞.规模以上工业企业技术创新效率的行业分析 [J].软科学,2013,27(11):58-65.
(责任编辑:冉春红)
