
不同生态区糜子种质资源的遗传多样性分析
2015-04-19 03:17连帅王瑞云马跃敏刘笑瑜季煦
山西农业大学学报(自然科学版) 2015年3期
连帅,王瑞云,2,*,马跃敏,刘笑瑜,季煦
(1.山西农业大学 农学院,山西 太谷 030801; 2.农业部黄土高原作物基因资源与种质创制重点实验室,山西 太原 030031)
不同生态区糜子种质资源的遗传多样性分析
连帅1,王瑞云1,2,*,马跃敏1,刘笑瑜1,季煦1
(1.山西农业大学 农学院,山西 太谷 030801; 2.农业部黄土高原作物基因资源与种质创制重点实验室,山西 太原 030031)
以5个糜子栽培生态区的40份糜子种质资源为试材,利用5个糜子特异性SSR标记对试材进行遗传多样性分析,旨在为糜子核心种质的构建和筛选提供一定的理论基础。结果表明:40份材料中共检测出15个等位基因变异,平均每个位点3个; 40份材料间的遗传相似系数为0.692~0.726; 聚类分析将试材分为5类,分别位于东北春糜子区、华北夏糜子区、北方春糜子区、黄土高原春夏糜子区和南方秋冬糜子区。同时,测定了以上试材的形态和生理指标(株高、主穗长和叶绿素含量),发现试材间存在多样性:(1)40个糜子材料的旗叶叶绿素含量介于27.23~43.10 g·L-1之间,倒二叶叶绿素含量介于17.00~41.17 g·L-1之间,其中185号材料(红糜子)叶绿素含量均最大。(2)40份材料的株高介于119.73~273.97 cm之间;其中,60号材料(镇原大糜子)最高,为273.97 cm,247号材料(黑糜子)最低,为119.73 cm;(3)主穗长介于17.00~66.07 cm;其中257号材料(黑糜子)最长,为66.07 cm,265号材料(鹅头)最短,为17.00 cm。通过对糜子材料部分形态、生理指标及5对糜子特异性SSR标记分析了遗传多样性,从聚类结果得出,SSR标记出的糜子多态性表现出了一定的地域性。
糜子;亲缘关系;SSR标记;聚类分析;多样性分析
糜子(PanicummiliaceumL.)是禾本科黍属一年生草本植物,耐温喜光,为短日照C4植物,生育期短,适合在各种土壤上生长,尤其是在贫瘠的沙壤土上,自然灾害发生的年份还可以作为救灾作物[1]。作为一种古老的农作物,糜子种植已有10 000多年的历史[2],起源于中国,是干旱和半干旱地区的主要粮食作物[3]。全世界糜子栽培面积约550~600万hm2,俄罗斯、乌克兰和中国栽培面积最大[1]。我国共收集到糜子种质资源10 200份,国家储备8 849份[4]。我国糜子资源广布于全国各省,分别属于7个糜子栽培区(东北春糜子区、华北夏糜子区、北方春糜子区、黄土高原春夏糜子区、西北春夏糜子区、青藏高原春糜子区和南方秋冬糜子区)[5]。糜子种质资源的多样性为生产和育种提供了更大空间。
有关糜子的多样性研究(形态、解剖、生理生化及分子遗传等方面)已经取得长足进展。胡兴雨等[6]对国家种质资源库中收集保存的8 016份黍稷种质资源的株高、千粒重、生育期等11个农艺性状进行主成分分析和聚类分析,将其分成5大组群,各个组群都有一定的形态学特征,为有效利用这些资源提供了理论依据。王纶等[7]采用反复干旱法,对山西省有代表性的500份黍稷种质资源进行了抗旱性鉴定评价,筛选出9份一级高度抗旱的种质。对一级高度抗旱的种质又进行了田间抗旱生理指标气孔导度、蒸腾速率、光合速率的测定和实验室离体叶片含水量的测定,进一步验证了反复干旱法对大批量黍稷种质资源抗旱性鉴定。王瑞云等[8]以6个糜子栽培生态区的18个糜子种质为材料,调查了糜子6个生育时期的8项形态、解剖及生理指标,发现糜子耐旱性因生态区的不同而不同,宁夏的“八升半”为较耐旱种质,山西的黄粮黍较不耐旱。慕芳等[9]通过对糜子叶表面结构的扫描电镜观察,发现其叶上下表面均有较厚的角质膜和蜡质,叶表面分布有3种类型的表皮毛。上述叶的结构特点反映出糜子抗旱性的特征。近年来,随着分子遗传学和基因工程的迅速发展,分子标记技术被广泛用于作物品种鉴定、纯度鉴定、亲缘关系鉴定、杂种优势和遗传多样性分析等研究领域。糜子中已有AFLP[10,11]、RAPD[12]和ISSR[13]标记构建及运用的研究。Karam等[10,11]分别用AFLP及银染AFLP标记检测糜子遗传多样性,但12份材料均来源于美国和加拿大,没有中国的。Lagler等[13]采用9个草莓属植物的ISSR标记检测了21份匈牙利糜子,发现只有7个引物可扩增出DNA片段,共检测到15个ISSR等位基因。但是关于从糜子基因组中分离微卫星序列或基于微卫星标记体系的构建方面的相关信息仍很缺乏[14]。2008年,Hu等[15]选取了水稻、小麦、大麦和燕麦的46个SSR标记分析了来源于中国不同生态区的118份糜子材料的遗传变异。2010年,Cho等[13]通过构建糜子基因组DNA的富SSR文库,发展了25个多态性微卫星标记,并对其稳定性进行了鉴定。Hunt等通过验证Cho等为糜子开发的25个SSR标记,发现其中只有16个标记稳定性较好,并且已利用这些标记分析了包括33份中国材料在内的来源于欧亚大陆的98份糜子的遗传多样性[16]。本研究利用5对糜子特异性引物分析来源于中国的40份糜子资源的遗传多样性,旨在为糜子核心种质的构建和筛选提供一定的理论依据。
1 材料与方法
1.1 试验材料
以中国农业科学院作物科学研究所提供的40个糜子种质资源(表1、图1)为试材。
1.2 试验方法
1.2.1 株高和主穗长的测定
用游标卡尺分别测量不同糜子材料的株高和主穗长度。
1.2.2 叶绿素含量的测定
用SPAD-502叶绿素测定仪测旗叶和倒二叶的叶绿素含量。
1.2.3 PCR扩增及等位基因检测
选取5对糜子特异性SSR引物[3](表2)进行PCR扩增。CTAB法提取糜子叶片DNA。PCR体系(20.2 μL)包括ddH2O 13.6 μL ,10×Taq Buffer 2 μL,dNTP 1 μL,上下游引物各0.4 μL,Taq DNA 聚合酶0.2 μL和模板DNA 3 μL。PCR 程序为: 94 ℃预变性5 min;94 ℃ 30 s,53 ℃退火 45 s,72 ℃延伸30 s,共30 个循环;72 ℃延伸10 min;4 ℃保存。扩增产物采用6%变性聚丙烯酰胺凝胶电泳检测。
1.3 数据分析
采用Microsoft Excel、SAS统计软件,运用方差分析处理数据,采用单因素方差分析(One-way ANOVA)比较不同生态区种质间的差异性,结果用平均值±标准误表示。

表1 供试糜子种质及其来源Table 1 The accessions and origin of broomcorn millet

图1 40个糜子材料地理分布图Fig.1 The geographical distribution of broomcorn millet germplasm

表2 SSR引物详情Table 2 SSR primers for details
在电泳胶板上,相同位置出现带的记为‘1’,无带则记为 ‘0’,将所有SSR引物所扩增带的结果输入数据表,利用NTSYS-pc软件统计分析,计算遗传距离(GD),采用非加权平均法(UPGMA)构建树状图。
2 结果与分析
2.1 不同糜子种质形态、生理指标的多样性分析
测定了40份糜子材料的4项形态和生理指标(表3)。由表3可以看出,不同糜子材料的株高、主穗长、旗叶叶绿素含量和倒二叶叶绿素含量都有显著差异(P<0.05)。40份材料的株高介于119.73~273.97 cm之间;其中,60号材料最高,为273.97 cm,247号材料最低,为119.73 cm;40份材料的主穗长介于17.00~66.07 cm之间;其中,257号材料最长,为66.07 cm,265号材料最短,为17.00 cm。就旗叶叶绿素含量而言,40个糜子材料的叶绿素含量介于27.23~43.10 g·L-1之间,其中185号最大,62号最小。就倒二叶叶绿素含量而言,40个糜子材料的叶绿素含量介于27.93~41.17 g·L-1,其中185号最大,62号最小。

表3 不同糜子种质形态和生理指标比较Table 3 The comparision of morphological and index plysiological of broomcorn millets
续表3
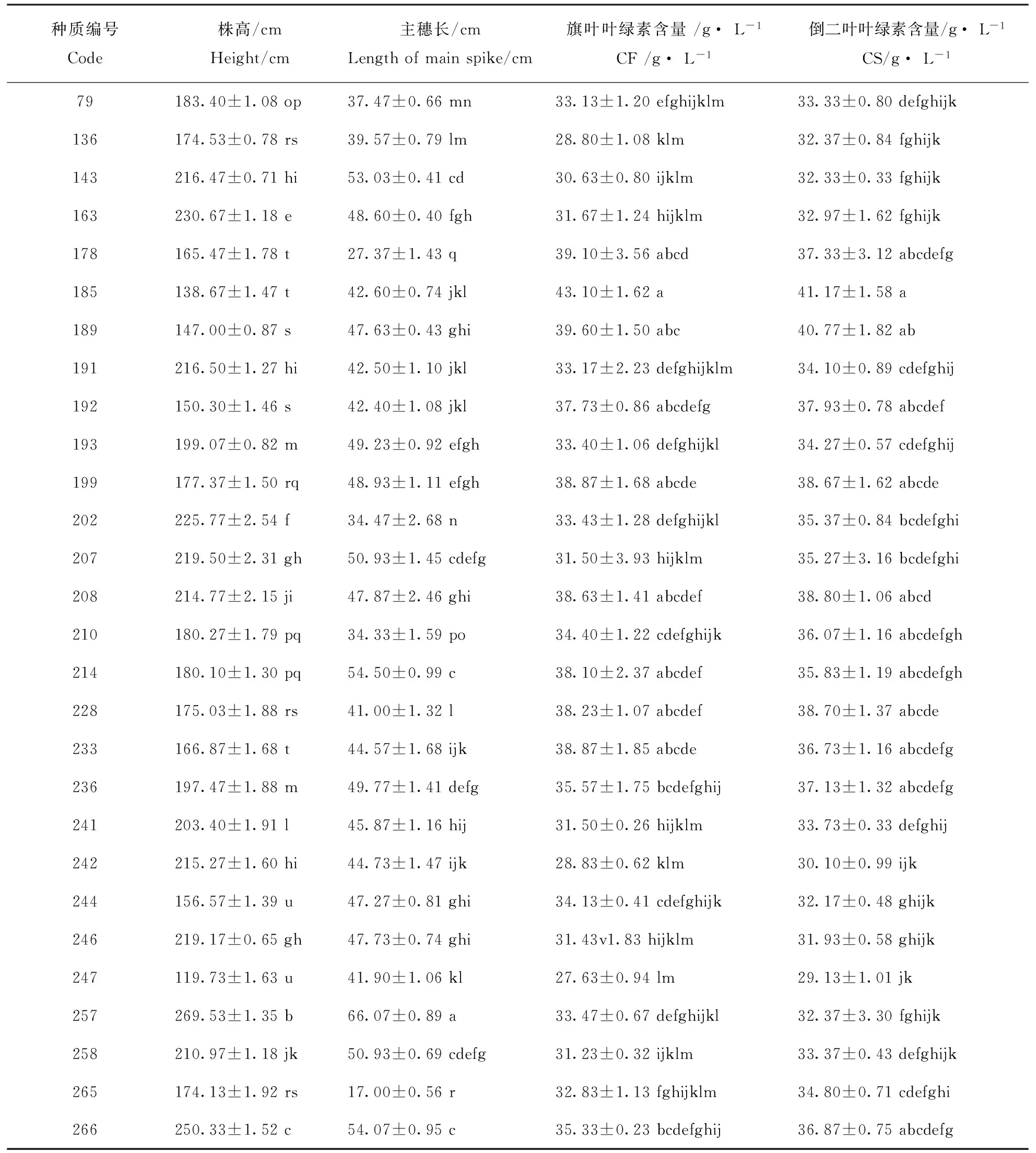
种质编号Code株高/cmHeight/cm主穗长/cmLengthofmainspike/cm旗叶叶绿素含量/g·L-1CF/g·L-1倒二叶叶绿素含量/g·L-1CS/g·L-179183.40±1.08op37.47±0.66mn33.13±1.20efghijklm33.33±0.80defghijk136174.53±0.78rs39.57±0.79lm28.80±1.08klm32.37±0.84fghijk143216.47±0.71hi53.03±0.41cd30.63±0.80ijklm32.33±0.33fghijk163230.67±1.18e48.60±0.40fgh31.67±1.24hijklm32.97±1.62fghijk178165.47±1.78t27.37±1.43q39.10±3.56abcd37.33±3.12abcdefg185138.67±1.47t42.60±0.74jkl43.10±1.62a41.17±1.58a189147.00±0.87s47.63±0.43ghi39.60±1.50abc40.77±1.82ab191216.50±1.27hi42.50±1.10jkl33.17±2.23defghijklm34.10±0.89cdefghij192150.30±1.46s42.40±1.08jkl37.73±0.86abcdefg37.93±0.78abcdef193199.07±0.82m49.23±0.92efgh33.40±1.06defghijkl34.27±0.57cdefghij199177.37±1.50rq48.93±1.11efgh38.87±1.68abcde38.67±1.62abcde202225.77±2.54f34.47±2.68n33.43±1.28defghijkl35.37±0.84bcdefghi207219.50±2.31gh50.93±1.45cdefg31.50±3.93hijklm35.27±3.16bcdefghi208214.77±2.15ji47.87±2.46ghi38.63±1.41abcdef38.80±1.06abcd210180.27±1.79pq34.33±1.59po34.40±1.22cdefghijk36.07±1.16abcdefgh214180.10±1.30pq54.50±0.99c38.10±2.37abcdef35.83±1.19abcdefgh228175.03±1.88rs41.00±1.32l38.23±1.07abcdef38.70±1.37abcde233166.87±1.68t44.57±1.68ijk38.87±1.85abcde36.73±1.16abcdefg236197.47±1.88m49.77±1.41defg35.57±1.75bcdefghij37.13±1.32abcdefg241203.40±1.91l45.87±1.16hij31.50±0.26hijklm33.73±0.33defghij242215.27±1.60hi44.73±1.47ijk28.83±0.62klm30.10±0.99ijk244156.57±1.39u47.27±0.81ghi34.13±0.41cdefghijk32.17±0.48ghijk246219.17±0.65gh47.73±0.74ghi31.43v1.83hijklm31.93±0.58ghijk247119.73±1.63u41.90±1.06kl27.63±0.94lm29.13±1.01jk257269.53±1.35b66.07±0.89a33.47±0.67defghijkl32.37±3.30fghijk258210.97±1.18jk50.93±0.69cdefg31.23±0.32ijklm33.37±0.43defghijk265174.13±1.92rs17.00±0.56r32.83±1.13fghijklm34.80±0.71cdefghi266250.33±1.52c54.07±0.95c35.33±0.23bcdefghij36.87±0.75abcdefg
注:表内数据为平均值±标准误;每列数据右侧字母不同者表示差异显著(P<0.05)。
Note:Values are mean ± SD.Values within a column followed by a different letter are significantly different atP<0.05.
2.2 不同糜子种质遗传多样性分析
利用5对SSR引物分析了40份糜子材料的遗传多样性,结果见表4。由表4可以看出,PIC值介于0.30~0.61之间,引物GB-PaM-115(图2)和GB-PaM-134的PIC>0.5时,该引物为高度多态性信息引物;引物GB-PaM-121(图3)、GB-PaM-126和GB-PaM-145(0.25 表4 SSR引物多态性分析Table 4 Diversity assessment based on SSR primers 图2 引物GB-PaM-115扩增后的条带Fig.2 Polymorphism amplified by primer GB-PaM-115 图3 引物GB-PaM-121扩增后的条带Fig.3 Polymorphism amplified by primer GB-PaM-121 由图4可以看出,40个糜子种质分布于5大生态区,基本符合糜子栽培生态区,但也有例外。例如材料185号(红糜子)和191号(白糜子)栽培于东北春糜子区,但是聚类结果将其分在了黄土高原春夏糜子区,具体原因有待深入研究。由图4可以看出,在遗传相似系数为0.62处,将糜子种质分为两类:金守黍和狸黍子分布于南方秋冬糜子区;其它的糜子常种植于北方。在遗传相似系数为0.656处,将糜子种质分为三类:第一类种植于北方春糜子区、黄土高原春夏糜子区;第二类种植于东北春糜子区、华北夏糜子区;第三类种植于南方秋冬糜子区。在遗传相似系数为0.71处,将糜子种质分为五类:第一类的9个材料来源于内蒙、甘肃地区,属于北方春糜子区;第二类的11个材料来源于山西、陕西、宁夏和甘肃等地区,属于黄土高原春夏糜子区;第三类的13个材料来源于黑龙江、吉林和辽宁等地区,属于东北春糜子区;第四类的5个材料来源于河北,属于华北夏糜子区;第五类的2个材料分别来源于广东和安徽,属于南方秋冬糜子区。从聚类结果看,SSR标记出的糜子多态性表现出了一定的地域性。 我国黍稷种质资源的研究从农艺性状的分析评价、品质分析、抗逆性分析到细胞学鉴定,取得了很大成果。2008年,胡兴雨等[6]对8 016份黍稷种质资源进行了农艺性状主成分分析和聚类分析。分成5大组群,各个组群都有一定的形态学特征,为有效利用这些资源提供理论依据。2011年,Hunt[16]等利用16个糜子特异性标记分析了98份糜子的遗传多样性,但是其中仅包括33份中国材料。在品种鉴定及遗传多样性研究方面,SSR是一种很常用且普遍有效的分子标记,在实践研究中已成为一种重要手段。本试验利用Hunt等[16]验证过的5对糜子特异性SSR标记分析了40个中国糜子材料的遗传多样性,同时调查部分形态和生理指标,发现相似系数为0.71处可将40个材料分为五大类,糜子的多态性表现出了一定的地域性。 图4 40个糜子品种聚类图Fig.4 Dendrogram of broomcorn millet germplasm 目前,我们课题组已经收集到400份来源于中国不同生态区的糜子资源,正在设计和筛选更多的引物,从多层次、多角度分析中国糜子种质资源的多样性,期望对糜子核心种质的筛选和构建提供一定的理论基础。 [1]郭琦.中国糜子(PanicummiliaceumL.)种质资源遗传多样性分析[D].太谷:山西农业大学硕士学位论文,2013. [2] Lu H Y,Zhang J P,Liu K B.Earliest domestication of common millet (Panicummiliaceum) in East Asia extended to 10,000 years ago[J].PNAS,2009,18:7367-7372. [3]王瑞云,刘晓欢,申鸽子,等.5个糜子品种的形态指标差异分析[J].山西农业科学,2013,41(11):1167-1169. [4]乔治军.糜子产业发展现状与思路[J].作物杂志,2013(5):25-27. [5]柴岩,冯佰利,王宏岩.中国黄米食品[M].陕西:西北农林科技大学出版社,2012:118-119. [6]胡兴雨,陆平,贺建波,等.黍稷农艺性状的主成分分析与聚类分析[J].植物遗传资源学报,2008,9(4):492-496. [7]王纶,温琪汾,曹厉萍,等.黍稷抗旱种质筛选及抗旱机理研究[J].山西农业科学,2007,35(4):31-34. [8]王瑞云,季煦,刘晓欢,等.不同生态区糜子种质间叶片及光合特性的多样性分析[J].山西农业大学学报(自然科学版),2014,34(2):98-102. [9]慕芳,冯佰利,王鹏科,等.糜子叶表面抗旱结构的扫描电镜观察[J].河北农业科学,2010,14(11):65-67. [10]Karam D,Westra P,Niessen S J,et al.Assessment of silver-stained AFLP markers for studying DNA polymorphism in proso millet (PanicummiliaceumL.)[J].Revista Brasileira de Botanica,2006,29(4):609-615. [11]Karam D,Westra P,Nissen S J,et al.Genetic diversity among proso millet (PanicummiliaceumL.) biotypes assessed by AFLP technique[J].Planta Daninha,2004,22(2):167-174. [12]M’Ribu H K,Hilu K W.Detection of interspecific and intraspecific variation inPanicummilletsthrough random amplified polymorphic DNA[J].Theoretical and Applied Genetics,1994,88(3/4):412-416. [13]Cho Y I,Chung J W,Lee G A ,et al.Development and characterization of twenty-five new polymorphic microsatellite markers in proso millet (PanicummiliaceumL.)[J].Genes and Genomics,2010,32(3):267-273. [14]Lágler R,Gyulai G,Humphreys M,et al.Morphological and molecular analysis of common millet (P.miliaceum) cultivars compared to a DNA sample from the 15th century (Hungary)[J].Euphytica,2005,146: 77-85. [15]Hu X Y,Wang J F,Lu P,et al.Assessment of genetic diversity in broomcorn millet (PanicummiliaceumL.) using SSR markers[J].Journal of Genetics and Genomics,2009,36:491-500. [16]Hunt H V,Campana M G,Lawes M C,et al.Genetic diversity and phylogeography of broomcorn millet (PanicummiliaceumL.) across Eurasia[J].Molecular Ecology,2011,20(22): 4756-4771. (编辑:武英耀) Genetic Diversity of Broomcorn Millet (PanicummiliaceumL.) Germplasms of Different Ecotype Zone of China Lian Shuai1,Wang Ruiyun1,2,*,Ma Yuemin1,Liu Xiaoyu1,Ji Xu1 (1.CollegeofAgriculture,ShanxiAgriculturalUniversity,TaiguShanxi030801,China;2.KeyLaboratoryofCropGeneResourcesandGermplasmEnhancementonLoessPlateau,MinistryofAgriculture,TaiyuanShanxi030031,China) Using 40 broomcorn millet germplasm resources as material,the genetic diversity was analyzed based on 5 simple sequence repeat (SSR) markers,to provide certain theoretical basis for the construction of core germplasm corn millet and filtering.The results showed: 15 alleles were detected among the materials,3 alleles were the average per locus.Genetic similarity coefficient among 40 accessions ranged from 0.692~0.726,which were classified into five categories including North China Spring Millet area,Summer millet area,Northern spring millet area in the Loess Plateau,the Spring and Summer millet zone and the South autumn and winter millet area,respectively.Meanwhile,the morphology and physiological indexes (plant height,ear length and the chlorophyll content)of the material were assayed.The results showed : (1) The chlorophyll content of flag leaf of the 40 accessions ranged from 27.23 g·L-1to 43.10 g·L-1,the chlorophyll content of blade in the second leaf from top ranged from 27.93 g·L-1to 41.17 g·L-1; among them the accession of No.185 (Hong mizi) was the biggest.(2) The plant height of the 40 accessions were ranged from 119.73 cm to 273.97 cm,among them,Accession 60 and 247 were the highest and the lowest,respectively; (3) The panicle length of main spike among all the accessions were ranged from 17.00 cm to 66.07 cm,257 and 265 were the longest and the shortest,respectively.Analyzed the genetic diversity through some of the physiological indicators and five pairs of millet specific SSR markers of millet material.Derived from the clustering results,SSR markers out of millet polymorphism showed some regional. PanicummiliaceumL; Genetic relationship; SSR marker; Cluster analysis; Diversity analysis 2014-11-04 2014-12-16 连帅(1991-),男(汉),山西临汾人,硕士研究生,研究方向:植物分子育种 *通讯作者:王瑞云,副教授,博士,硕士生导师。Tel: 15234420135;E-mail: wry925@126 com 国家自然科学基金(31271791) S516 A 1671-8151(2015)03-0225-07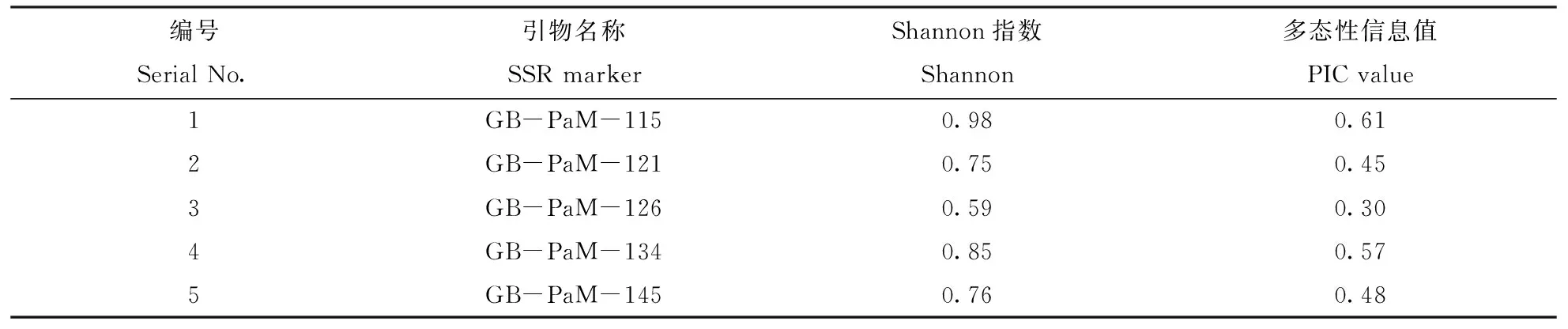


3 讨论与结论

猜你喜欢
今日农业(2022年13期)2022-09-15
中国粮油学报(2022年6期)2022-07-23
今日农业(2022年8期)2022-07-05
山西农业科学(2021年1期)2021-01-18
现代农村科技(2019年12期)2019-01-06
江西建材(2018年4期)2018-04-10
中国麻业科学(2018年6期)2018-04-09
西南农业学报(2016年5期)2016-05-17
西南农业学报(2016年6期)2016-04-16
广西林业科学(2016年3期)2016-03-16
