
山西不同居群赖草生产性能及营养价值评价
2015-04-10 07:30樊鹏鹏杜利霞霍亚迪董宽虎
草业科学 2015年8期
樊鹏鹏,杜利霞,霍亚迪,董宽虎
(山西农业大学动物科技学院,山西 太谷030801)
赖草(Leymus secalinus)属禾本科早熟禾亚科(Pooideae)小麦族赖草属,是一种根茎型旱中生禾草,无性繁殖能力强[1]。在山西各地均有零星生长,但主要分布在晋北地区,尤其在一些水分条件较好、盐渍化程度较轻的地区(河谷冲积平原荒地或水渠边沿)已生长成繁茂的株丛,并以强壮的根茎迅速繁衍,成为独立的优势群落[2-3]。赖草生境极其多样,在海拔500 ~4 700 m、从湿润的盐碱滩地和海滨滩地到干旱高温的沙土草原、荒漠化草原皆有生长,为草原、草甸的主要成分,具有广泛的适应性和较高的抗逆性[4]。此外,赖草在抽穗前为各种牲畜所喜食,也是一种饲用价值较高的优良牧草。
目前,对于赖草的研究从形态组织结构[5-8]、生理 特 性[9-12]、抗 旱 性[13-15]、种 子 抗 盐 性[16-17]、分子[18-20]等方面都有报道,但育成的赖草品种却很少。本试验以采集于山西省不同地区的8 个野生赖草居群为研究对象,进行人工栽培。通过对山西不同居群的赖草生产性能及营养价值的分析,进而评价野生赖草引种驯化的潜力及利用价值,为今后选育驯化野生赖草种质资源提供科学依据。
1 材料与方法
1.1 试验地自然概况
试验地设在山西农业大学动物科技学院试验站草业科学试验地(37°25' N、112°23' E,海拔799 m)内,土壤为石灰性褐土,pH 7.5。年平均气温9.5 ~10.5 ℃,最热月平均气温23.5 ~24.0 ℃,最冷月平均气温-6.5 ~-5.0 ℃,无霜期160 ~165 d,年平均降水量450 ~573 mm[21]。
1.2 试验材料
2010 年9 月27 日―10 月7 日分别从山西不同地区采集了8 个自然居群的赖草种子,各居群的地理位置与生境概况如表1 所示。
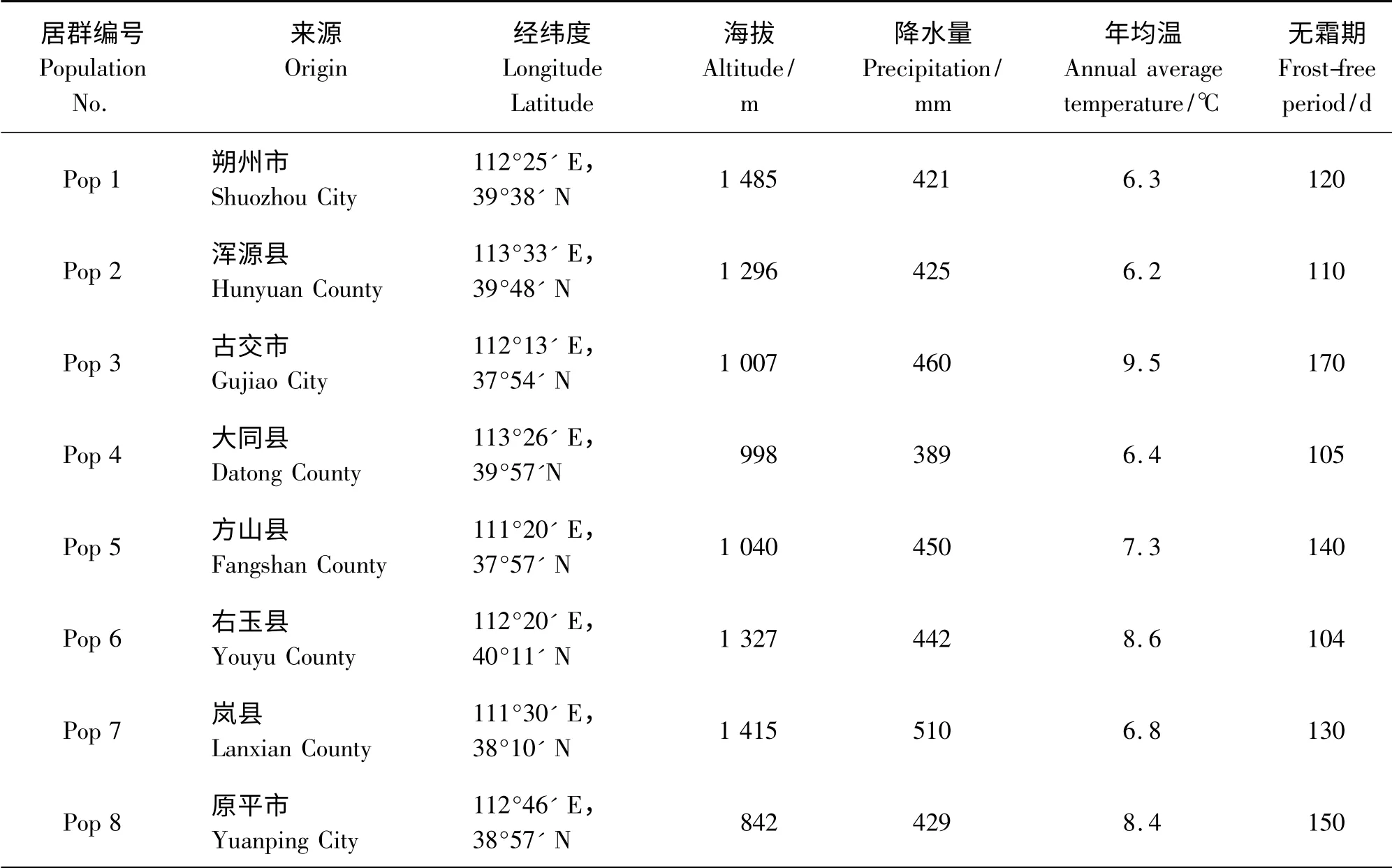
表1 赖草种子来源及所处生境条件Table 1 The source and habitat of L. secalinus
1.3 试验设计
采用完全随机区组设计,3 次重复,一共24 个小区,每个小区面积为3.6 m ×1.8 m,小区间间隔为100 cm。于2010 年4 月中旬播种,播种前后均不施肥,播种前深翻土地并耙平,人工开沟条播,行距30 cm,种植6 行,播深2.5 cm,播种量22.5 kg·hm-2。试验于2013 年6 月进行。
1.4 测定项目及方法
1.4.1 地上生物量测定 各小区于成熟期刈割,留茬高度3 ~4 cm。测定时剔除小区两侧边行及两端50 cm 之内的面积,剩余的全部刈割,之后立即称取鲜草重[22]。将草样105 ℃杀青30 min,65 ℃烘干2 ~3 d 之后,称干草重后推算各小区干草产量,同时对样品进行营养成分的测定。
1.4.2 种子繁殖性能测定 测定的指标有穗数、穗长、单株小穗数、单株小花数、饱满种子数。测定方法为每小区随机选取1 m×1 m 样方,数样方内的生殖枝数,后随机采集10 株生殖枝条,测定穗长计算平均值,同时计数每一枝条上的小穗数、小花数、饱满种子数,计算结实率、千粒重。结实率(%)=(单株饱满种子数/单株小花数)×100%;从成熟种子中随机测定200 粒种子的重量,重复3 次,计算千粒重[23]。
1.4.3 营养成分分析 烘干的样品粉碎后过40 目筛(0.425 mm)用于营养成分测定。按实验室常规分析方法[24]分析各样品中的粗蛋白质(Crude protein,CP)、粗灰分(Ash content,Ash)、钙(Calcium,Ca)、磷(Phosphorus,P)、中性洗涤纤维(Neutral Detergent Fiber,NDF)、酸性洗涤纤维(Acid Detergent Fiber,ADF)、粗脂肪(Ether extract,EE)。
1.5 数据统计分析
数据采用SAS 9.0 软件对所测数据进行单因素方差分析和主成分分析,用Duncan 法对各数据进行多重比较,平均值和标准误表示测定结果;采用Excel 2007 制图。
2 结果与分析
2.1 不同居群赖草的干草产量
不同居群赖草干草产量差异明显(图1),最高的是Pop 2,可达9.37 t·hm-2,其次是Pop 6,为7.62 t·hm-2;产量较低的是Pop 1 和Pop 5,分别为5.39 和6.44 t·hm-2。
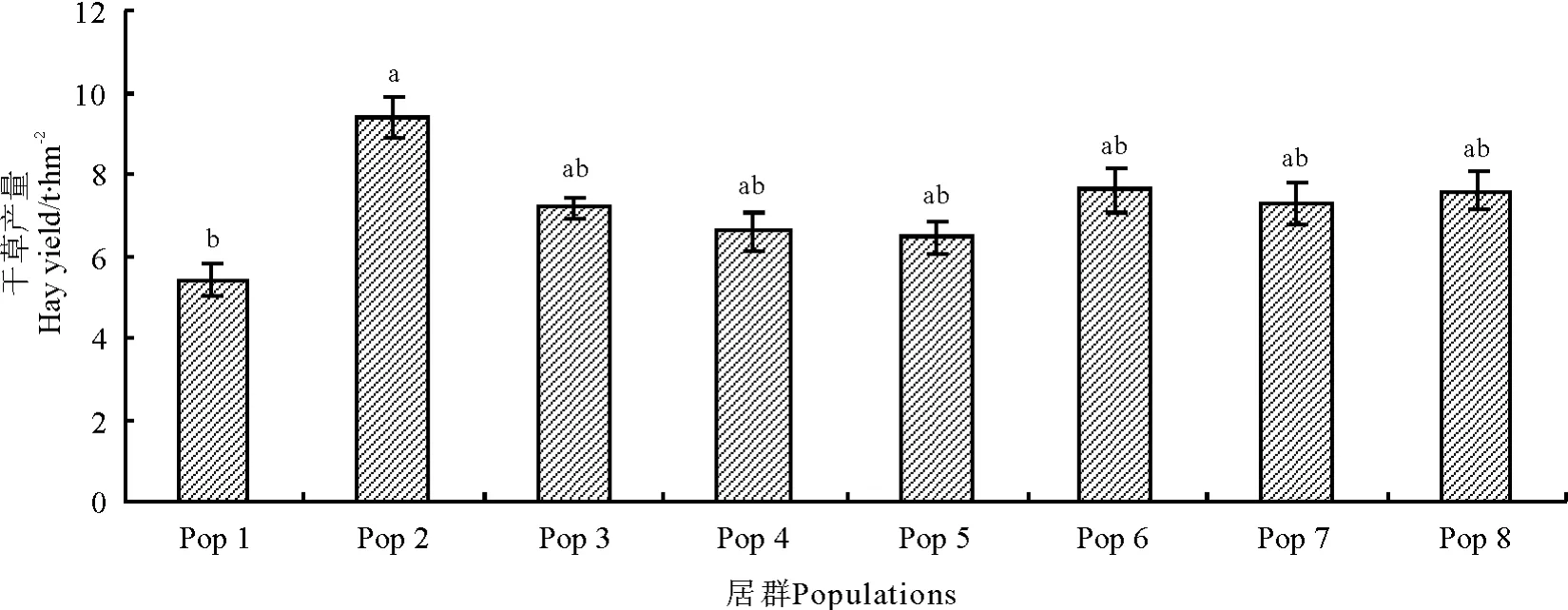
图1 不同居群赖草的平均干草产量比较Fig.1 Average hay yield of different L. secalinus populations
2.2 不同居群赖草的种子繁殖性状
Pop 2 的穗长明显长于其他居群,长21.21 cm,其次是Pop 6,长约19.86 cm;Pop 5 的赖草平均穗长最短,只有15.39 cm(表2)。各居群间生殖枝数、单株小穗数、单株小花数、饱满种子数均无显著差异(P >0.05)。Pop 7 的千粒重最大,为3.33 g,显著高于千粒重最小的Pop 5(P <0.05),其次为Pop 2、Pop 3。不同居群赖草结实率以Pop 2 最高,达63.02%,显著高于结实率最低的Pop 8。综合来看,Pop 2 的种子繁殖性状较好。
2.3 不同居群赖草的营养成分
CP 含量最高的是Pop 1,为10.99%,其次是Pop 2 和Pop 6,分别为9.93%和9.90%;含量最低的是Pop 3,只有6.64%,Pop 1 显著高于Pop 3 和Pop 5(P <0.05)。Pop 1 的Ca 含量也最高,可达3.65%,其次是Pop 2,为3.05%,Pop 4 的Ca 含量最低,只有1.70%。但Pop 4 的Ash 含量最高,为7.70%,其次是Pop 1、Pop 6 和Pop 2,含量分别为7.52%、7. 29%和6. 94%。P 含量最高的是Pop 4,约为0.19%;最小是Pop 2,只有0.12%(表3)。另外,Pop 6 的NDF、ADF 含量均最小,分别为62.01%和34.53%;且Pop 6 的EE 含量也较高,为4. 64%,与最高的Pop 8 差异不显著(P >0.05)。从各营养成分综合分析,Pop 6 的营养价值较高。
2.4 不同居群赖草种子繁殖性状的主成分分析
主成分分析表明(表4、表5),前3 个主成分特征值的累积贡献率达81.10%。第1 主成分的特征值为2.707,贡献率为38.68%;其中穗长的特征向量最大,为0.487,其次是饱满种子数(0.486)、千粒重(0.457);因此,可将第1 主成分称为穗长因子。第2 主成分的特征值是1.557,贡献率22.24%;影响第2 主成分的特征向量主要是单株小花数(-0.605),其次为结实率(0.463);因此,第2 主成分可以称为小花数因子。第3 主成分的特征值为1.413,贡献率为20.18%;生殖枝数在第3 主成分中的特征向量最大,为0.666,其次为单株小穗数(0.515);因此将第3 主成分称为生殖枝数因子。
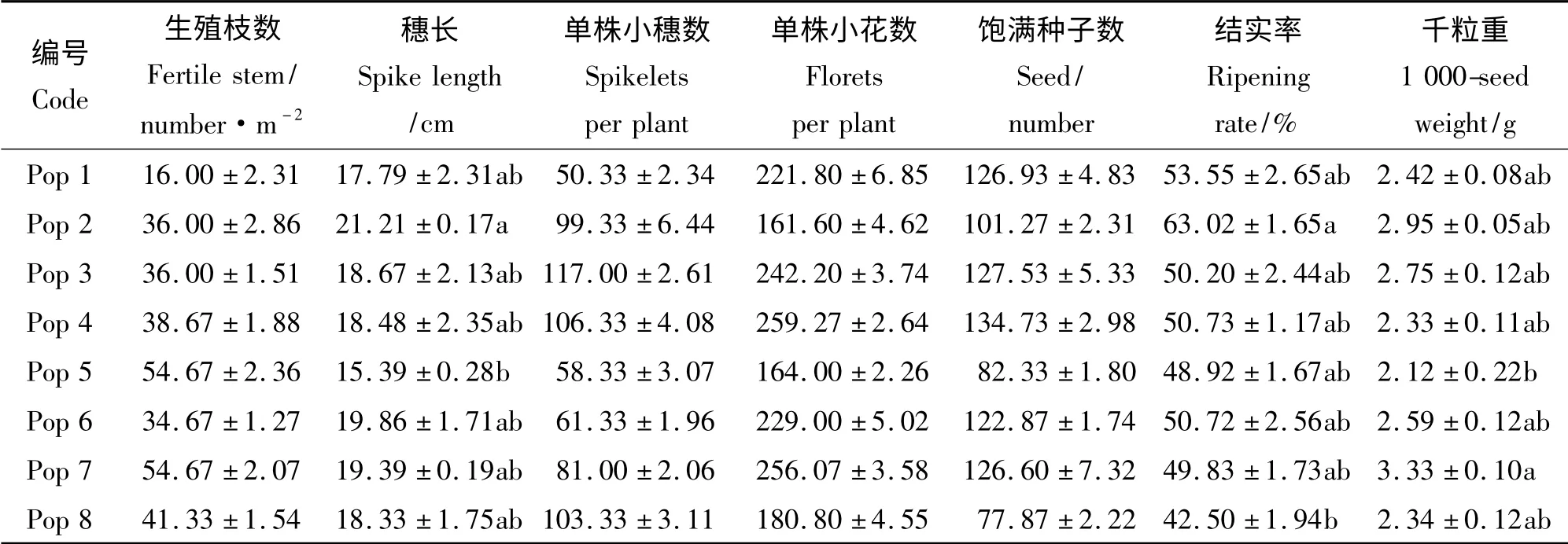
表2 不同居群赖草的种子繁殖性状Table 2 Seed reproduction traits of different L. secalinus populations
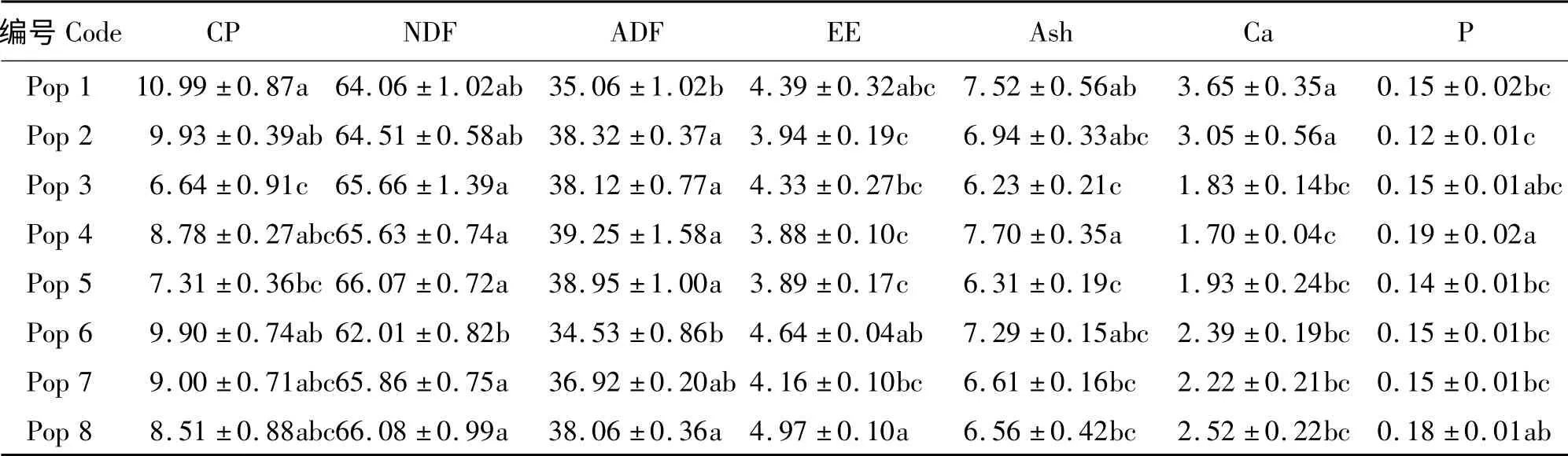
表3 不同居群赖草的营养成分Table 3 Nutrients of different L. secalinus populations %
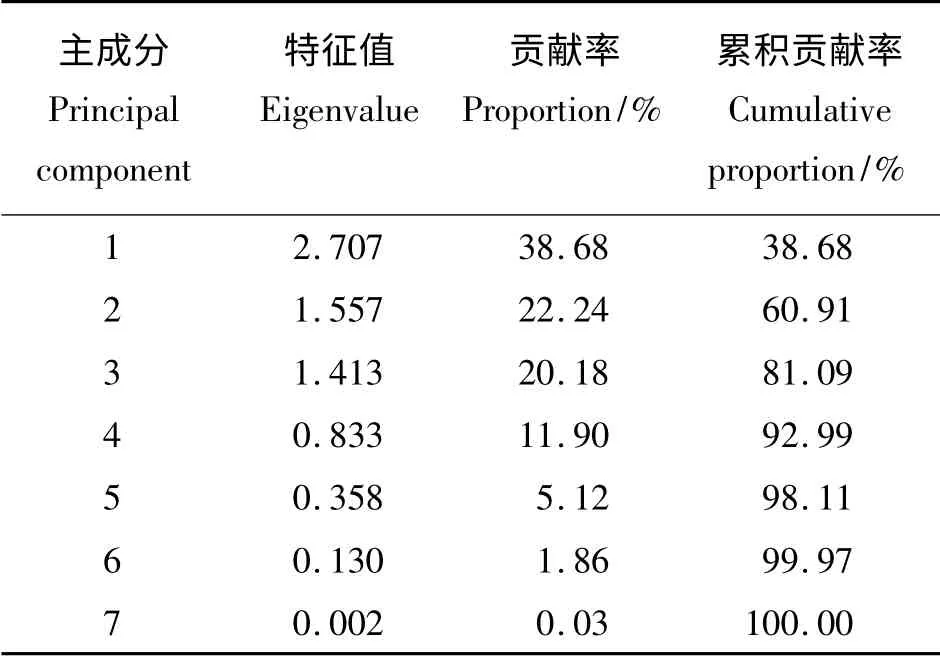
表4 7 个种子繁殖性状相关矩阵的特征值、贡献率和累积贡献率Table 4 Eigenvalue,proportion and cumulative proportion of the correlation matrix of 7 seed reproduction traits
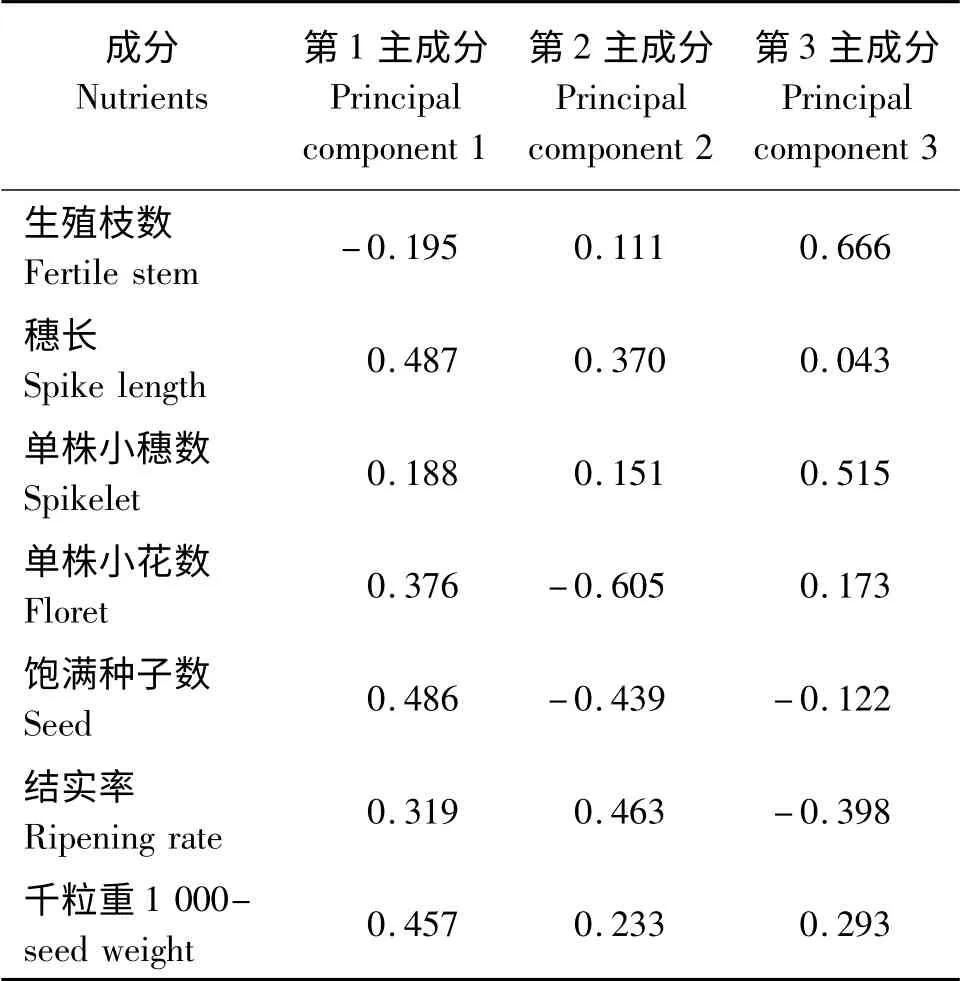
表5 选取的主成分对应的特征向量Table 5 Eigenvectors of selected principal components
2.5 不同居群赖草营养成分的主成分分析
主成分分析结果(表6 和表7)表明,前3 个主成分的特征值都大于1,累积贡献率达87.88%。第1 主成分的特征值为3.568,贡献率为50.97%,其中CP(0. 484)特征向量最高,其次为ADF(-0.464)、NDF(-0.445)、Ca(0.427),因此可将第1 主成分称为CP 因子。第2 主成分的特征值为1.455,贡献率为20.78%,特征值较大的有EE(0.696),因此第2主成分可以成为EE因子。第3 主成分的特征值为1.130,贡献率为16.13%,Ash(0.704)在第3 主成分中的特征向量最大,可将第3 主成分称为Ash 因子。对于特征向量中存在正负的现象,可能是因为性状之间有着相互制约的关系。
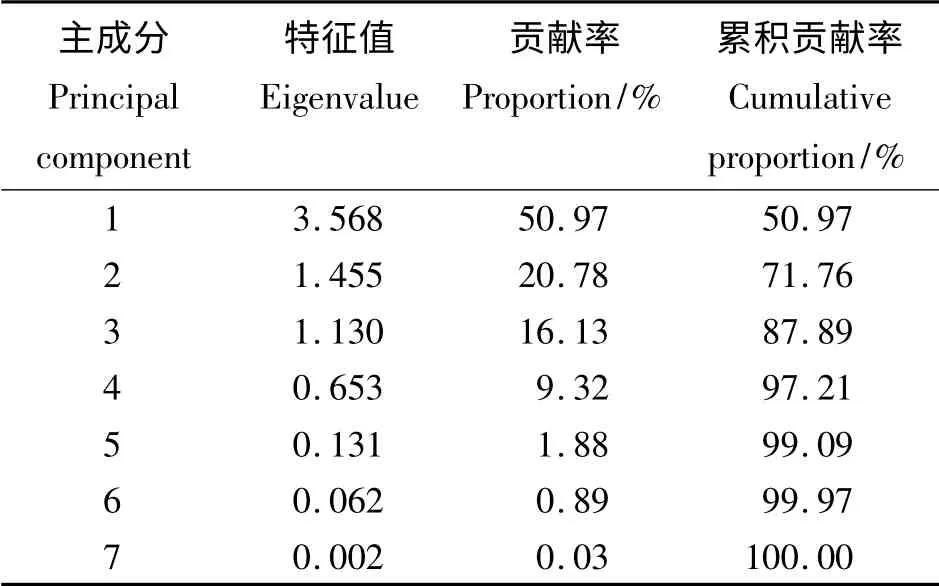
表6 7 个营养成分相关矩阵的特征值、贡献率和累积贡献率Table 6 Eigenvalue,proportion and cumulative proportion of the correlation matrix of 7 nutritional components
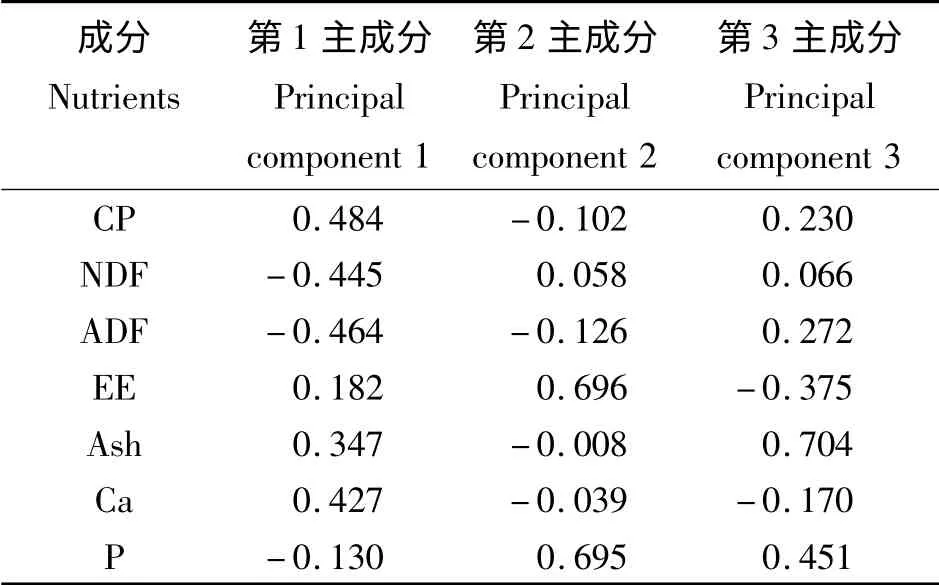
表7 选取的主成分对应的特征向量Table 7 Eigenvectors of selected principal components
3 讨论与结论
不同居群赖草的干草产量和种子繁殖性能均不同,这可能是因为不同赖草居群生长环境不同,在进化过程中为了适应不同的生存环境条件而产生的形态学多样性。浑源居群(Pop 2)赖草干草产量最大,单株小穗数、结实率和千粒重也相对较高,与此相反的是方山居群(Pop 5),该居群赖草干草产量较小,单株小穗数、结实率和千粒重也相对较低,这说明赖草的营养繁殖与种子繁殖有一定的相关性,与武路广等[23]对白羊草(Bothriochloa ischaemum)主要生产性能构成因子的研究结果一致。此外,本研究还发现8 个不同居群赖草穗长在15.39 ~21.21 cm,变化幅度不大,这与杨瑞武等[25]对赖草属11 个物种的形态学特征比较研究结果相同,表明赖草属物种的穗长在种内具有相对的稳定性。
新麦草(Psathyrostachys juncea)、羊草(Leymus chinensis)、垂穗披碱草(Elymus nutans)等禾本科牧草的营养成分分析[26]表明,新麦草在成熟期的CP含量为11.29%,EE 含量为4.93%,这两项指标均高于本研究采集的8 个赖草居群;Ash 含量为6.33%,仅高于本研究中古交和方山居群;Ca 含量只有1.33%,低于本研究的8 个赖草居群,表明赖草的矿物质含量要优于新麦草。桂荣等[27]对采集于内蒙古地区的赖草及羊草的营养动态进行了研究,发现成熟期赖草CP 含量只有7.19%,ADF 含量为48.69%,与本研究相比,CP 含量仅高于本研究的古交居群,但ADF 含量高于8 个居群中ADF 含量最高的大同居群,可见本研究采集的大部分居群赖草综合营养品质都要优于内蒙古地区的赖草,且适口性也要比内蒙古地区的赖草居群好,这可能是因为赖草生长环境不同而造成的差异;成熟期羊草CP含量为11.35%,ADF 含量仅37.00%,与本研究相比CP 含量要高于本研究的8 个赖草居群,ADF 含量仅高于朔州、右玉和岚县居群,这表明羊草的营养品质要优于本研究中的大部分赖草居群。马鸣[28]对垂穗披碱草营养成分分析研究得出,垂穗披碱草CP、EE、Ash 含量分别为6.60%、1.92%、4.78%,相比来说,这3 项营养指标均低于本研究的8 个赖草居群,说明赖草综合营养品质要优于垂穗披碱草。本研究只对成熟期赖草的生产性能和营养成分进行了研究,之后将对其他不同生育期的赖草作后续分析。
种子繁殖性状主成分分析表明,前3 个主成分因子中特征向量最大的分别为穗长、小花数和生殖枝数,说明这3 项指标值越大,种子繁殖性状越优异。对营养成分进行主成分分析,得到累积贡献率达87.89%的3 个主成分,这3 个主成分因子中特征向量最大的分别是粗蛋白、粗脂肪和粗灰分,粗蛋白是必不可少的营养物质,粗脂肪是提供热能的主要原料,粗灰分代表牧草中的矿物质,因此牧草中这些物质含量越高,牧草品质也就越好。
通过对8 个不同居群赖草成熟期的生产性能与营养价值综合分析,右玉居群表现最好,浑源居群较好,这两个居群可作为今后育种工作的重点选育对象。
[1] 蔡联炳,苏旭.国产赖草属的分类修订[J].植物研究,2007,27(6):651-660.
[2] 董宽虎.山西牧草种质资源[M].北京:中国农业科学技术出版社,2010:58-59.
[3] 钱崇澍,陈焕镛.中国植物志[M].北京:科学出版社,2004:162-163.
[4] 王丽焕,道理刚,肖冰雪,杨满业.赖草抗旱性研究进展[J].草业与畜牧,2007(11):1-4.
[5] 蔡联炳,张梅妞.国产赖草属的叶表皮特征与组群划分[J].植物研究,2005,25(4):400-405.
[6] 王六英,赵金花.赖草属(Leymus Hochst.)3 种牧草营养器官耐旱结构的研究[J].干旱区资源与环境,2001,15(5):23-27.
[7] Parminder K,Ivan W M,Steven R L,Bushman B S,Alvaro G H,Kim W R,Liu L,Mark A M.Gene expression polymorphisms and ESTs associated with gravitropic response of subterranean branch meristems and growth habit in Leymus wildryes[J].Plant Science,2008,175(3):330-338.
[8] 沙莉娜.赖草属植物的形态学、细胞学与分子系统学研究[D].成都:四川农业大学博士论文,2008.
[9] 杨瑞武,魏秀华,周永红,郑有良.赖草属植物醇溶蛋白的遗传多态性[J].云南植物研究,2004,26(1):103-110.
[10] 温涛,陈伟,史静,陈本建.温度对赖草种子萌发及幼苗生长的影响[J].青海草业,2013,22(3):2-5.
[11] 杜利霞,董宽虎,乔志宏,韩汝旦,娄世杰.NaCl 胁迫对赖草幼苗生理特性的影响[J].草原与草坪,2013,33(1):1-4.
[12] 杜利霞,董宽虎,乔志宏,韩汝旦,娄世杰.NaHCO3胁迫对赖草几种生理指标的影响[J].山西农业科学,2012,40(11):1160-1163.
[13] 易津,谷安琳,贾光宏,吴萧.赖草属牧草幼苗耐旱性生理基础的研究[J].干旱区资源与环境,2001,15(5):47-50.
[14] Liu S H,Kang Y H,Wan S Q,Wang Z C,Liang Z W,Sun X J.Water and salt regulation and its effects on Leymus chinensis growth under drip irrigation in saline-sodic soils of the Songnen Plain[J].Agricultural Water Management,2011,98(9):1469-1476.
[15] 叶煜辉,江明锋,陈艳,杨满业.赖草属植物的抗逆性研究进展与应用前景[J].生物学杂志,2009,26(4):54-57.
[16] 孙菊,杨允菲.盐胁迫对赖草种子萌发及其胚生长的影响[J].四川草原,2006(3):17-20.
[17] 白玉娥,易津,谷安琳,国振杰.八种根茎类禾草种子耐盐性研究[J].中国草地学报,2005,27(2):55-59.
[18] 高刚,苟学梅,王茜,张艳,李洋益,邓家彬,杨瑞武.赖草LsDREB2 转录因子基因的克隆与生物信息学分析[J].广西植物,2014,34:1-6.
[19] 史仁玖,郝岗平,赵茂林,杨清.多枝赖草Glutathione Reductase 基因克隆及胁迫表达分析[J].四川师范大学学报(自然科学版),2008,31(4):471-475.
[20] Fan X,Liu J,Sha L N,Sun G L,Hu Z Q,Zeng J,Kang H Y,Zhang H Q,Wang Y,Wang X L,Zhang L,Ding C B,Yang R W,Zheng Y L,Zhou Y H.Evolutionary pattern of rDNA following polyploidy in Leymus (Triticeae:Poaceae)[J].Molecular Phylogenetics and Evolution,2014,77:296-306.
[21] 邢毅,赵祥,董宽虎,史威威.不同居群达乌里胡枝子形态变异研究[J].草业学报,2008,17(4):26-31.
[22] 苏加楷,张文淑.规范牧草产量试验小区面积与测产方法[J].草地学报,2008,16(4):324-327.
[23] 武路广,霍梅俊,董宽虎.白羊草主要生产性能构成因子分析[J].中国草地学报,2013,35(4):61-65.
[24] 张丽英.饲料分析及饲料质量检测技术[M].北京:中国农业大学出版社,2003:35-41.
[25] 杨瑞武,周永红,郑有良,丁春邦.赖草属植物11 个物种的形态学特征比较研究[J].四川农业大学学报,2003,23(3):196-199.
[26] 王勇.新麦草新品系生物学特性及生产利用性能的研究[D].呼和浩特:内蒙古农业大学硕士论文,2005.
[27] 桂荣,那日苏,夏明,玉兰.主要栽培牧草营养动态及适宜利用时期[J].中国草地学报,1998(5):38-45.
[28] 马鸣.4 种禾本科牧草生产性能及营养价值研究[D].兰州:甘肃农业大学硕士论文,2008.
猜你喜欢
九江职业技术学院学报(2022年1期)2022-12-02
河北科技师范学院学报(2022年2期)2022-08-26
保定学院学报(2022年2期)2022-04-07
浙江中医药大学学报(2021年6期)2021-07-12
中国粮油学报(2020年12期)2021-01-09
军事运筹与系统工程(2020年2期)2020-11-16
许昌学院学报(2018年4期)2018-05-02
军事运筹与系统工程(2018年3期)2018-03-26
中华建设(2017年1期)2017-06-07
中亚信息(2016年10期)2016-02-13
