
无患子物候及开花结果特性1)
2015-04-03 11:31高媛贾黎明苏淑钗戴丽莉翁震郭有枝翁学煌
东北林业大学学报 2015年6期
高媛 贾黎明 苏淑钗 戴丽莉 翁震 郭有枝 翁学煌
(省部共建森林培育与保护教育部重点实验室(北京林业大学),北京,100083) (福建源华林业生物科技有限公司)
随着石油、煤炭、天然气等储量的日益减少,全球能源发展趋势已逐步由多元化结构向可再生能源转化,以实现对化石燃料的逐步替代[1]。当前,世界上众多国家对生物柴油、燃料乙醇、固体颗粒燃料、沼气等生物质能源已进行了开发与利用[2-3],如欧美国家大力发展生物质能,北欧各国大力发展木材发电,德国大力发展沼气。在林业生物质能上美国、瑞典、加拿大、巴西等围绕杨树、柳树、桉树等开展了纤维素醇类的研究工作;欧盟、印度、马来西亚等主要以油菜籽、向日葵、棕榈等作为生产生物柴油的原料进行研发[4-12]。针对我国的特殊国情,考虑到耕地面积的红线问题,林业生物质能源成为了我国生物质能源主要的发展方向[13-14]。
无患子(Sapindus mukorossi Gaertn)为无患子科(Sapindaceae)无患子属(Sapindus. L)落叶大乔木,又名肥皂树、洗手果等[15],是我国特有的木本油料树种。主要产自我国东部、南部、西南部,为低山、丘陵及石灰岩山地常见树种[16]。因为其分布广泛,且种仁含油率高达40%以上,成为我国“林油一体化”产业发展中的重要生物质能源树种之一,可用来提取油脂,制造天然润滑油和生物柴油[17]。同时,无患子果皮中富含皂苷,是优良的洗涤化妆品原料[18];根、果还可作中药材等,是集生物质能源、化工和医药于一体的多功能原料树种。根据《全国林业生物质能源发展规划(2011—2020 年)》,计划至2020 年建设无患子能源林基地共25 万hm2,发展前景十分看好[19]。国内对无患子的研究较少,通过CNKI 检索发现从1990 年至今多为无患子油脂、皂苷提取等工艺及产品等的研究;与原料林培育相关的多着重于无患子不同种源的形态、油脂、皂苷、生理特征等变化的对比研究以及播种育苗、植苗造林、截干造林等栽培技术措施研究。国外SCI 检索发现其主要侧重于油脂基因分析、各类型新皂苷提取工艺、表面活性剂、微体繁殖、生物医药等研究。而对其生物、生态学特性缺乏较为深入的研究,针对无患子原料林培育中的树体管理、花果调控、水肥管理等方面也无实质性进展,因此在解决原料林培育中的落花落果现象,构建科学合理的树体管理、花果调控等技术体系时困难极大[20-21]。为此,笔者对无患子的生长节律、开花结果习性、枝类组成等主要生物学特性进行了调查研究,旨在为进一步开发和利用无患子提供理论依据,指导花果调控及树体管理等,并为无患子生物柴油原料林的高效培育奠定基础。
1 研究区概况
研究地点在福建省三明市建宁县,位于东经116°35′~117°04′,北纬26°30′~27°06′,海拔在280~1 858 m,是我国发展无患子生物柴油原料林的主要基地。研究地点属中亚热带海洋性季风气候,又兼有大陆性山地气候特点,年平均气温17.0 ℃,年平均降水量1 950 mm,多集中在春夏两季。常年相对湿度84%,平均日照时间为1 721 h。成土母岩为变质岩、岩浆岩和沉积岩3 大类,红壤面积占78.3%、黄壤占17.8%、紫色土占3.6%、草甸土占0.3%。质地多为轻壤、中壤,肥力较高,具有良好的水湿条件,适宜林木生长[22]。
研究林分属福建源华林业生物科技有限公司,为2009 年开始营造的无患子原料林,现有林分5 200 hm2,其中山地造林5 200 hm2,四旁种植0.67 hm2。造林地土壤多为黄壤或黄红壤。采用梯田结合穴状的整地方法,穴规格为50 cm×40 cm×40 cm。均为植苗造林,种源均来自福建、浙江及江西三省,一般选取2 年生的苗木种植,栽植密度为630 ~825株/hm2,苗木类型有播种苗、嫁接苗、移植苗等。在管理方面,每年修剪1 次、施肥1 次、除草1 次、病虫害防治3~5 次。
2 材料与方法
2.1 试验材料
在试验地选取具有代表性的30 株健康的无患子植株进行生物学特性观察,其种源来自浙江省天台县,现已在福建省建宁县种植6 a。株行距为3.5 m×3.5 m,树高为(4.4±1)m,冠幅为(4±1)m。每株树上选取3 个花序(果序)(树体N—E—S 方向上,中部的外侧顶生花(果)序)、3 个结果枝(树体N—E—S 方向上,中上部当年生并着生花序的枝条)、3个营养枝(树体N—E—S 方向上,中部当年生未形成花序的枝条)进行标记追踪调查,果实成熟期时每株树摘取30 个果实(4 个方向上的果序随机摘取)进行性状检测。
2.2 试验方法
物候期观测:调查时间为2013 年1 月至2014年11 月。物候期观测包括芽膨胀期、芽展开期、抽梢期、初花期、盛花期、末花期、子房膨大期、果实形成期、果实成熟期、落叶期等,采用目测法,每5 ~10 d 观测并详细记录1 次[23]。
开花、结果习性调查:开花、结果习性调查包括开花性状和结实性状等的调查[24]。开花性状调查包括花期发育变化及可孕花比例[25];结果性状调查包括坐果率、果枝组成情况(结果枝、营养枝及童期枝)、落花落果情况及果实性状的分析等[26]。调查初花期、盛花期、末花期、初果期、果实膨大期及果实成熟期所标记顶生花序及30 个侧生花序上的花数(不可孕花和可孕花分开统计)及果数,用卷尺测量花序、结果枝、营养枝的长度等,用游标卡尺测量结果枝、营养枝和果实的直径[27]。各花期雌雄蕊变化采用显微镜进行观测并拍照。对整个生长季进行追踪调查,观测花序和果实的生长进程及数量动态变化。雌雄比=可孕花数量/不可孕花数量,落花(果)率=((前一时期坐果(花)数-后一时期坐果(花)数)/前一时期坐果(花)数量)×100%,坐果率=(结果数量/总开花数量)×100%。
光照度、气象因子调查:用ST-85 型照度计,所显示的数值乘以100 即为照度值(单位:lx)。采果前期(林内叶面积指数ILA=2.16)测定所标记果序的光照度(树冠东面果序),测试时间于晴天的上午10:00,空地光照度于90 000 lx 以上,以选出同期坐果率最佳的光照度值。气温、湿度及降雨量等气象因子来源于福建省建宁县气象局所提供的实时测定数据。
数据处理:采用Microsoft Excel 2007 和SPSS Statistics 20 统计分析软件进行数据计算处理,结果枝组分类采用K—均值聚类法进行聚类。
3 结果与分析
3.1 物候期
对2013—2014 年福建建宁无患子进行了2 a 的物候期观测(表1)。2013 年无患子萌动于2 月10日,在持续10 d 出现大于10 ℃的日均温后;其萌芽于2 月20 日,日均温达到11 ℃以上;3 月中旬树木全面抽梢,此时当地近10 d 的平均温度达到14.2℃;4 月5 日开始长出小花序时平均温度已达18℃,平均湿度高达90%,10 d 内的平均日降雨量达12.1 mm;4 月中旬,花序长出80%,相近10 d 内均无降雨;4 月20 日叶片转色基本完成,颜色由浅转深;5 月12 日可明显看见绿色小花苞,此时日均温达22℃,之后便进入当地的连续性降雨期;5 月19 日进入初花期,全树有5%的花开放,一周后进入盛花期,从初花期到末花期,花期约15 d,在经过约一周的授粉后在6 月10 日进入初果期。在各花期时,建宁最高日降雨量可达43.1 mm,对授粉及坐果影响极大;6 月27 日进入果实膨大期,此后日均温逐步上升大于27 ℃;9 月1 日果实开始转色,进入初熟期,当地温度逐渐下降,林内日均温达25 ℃;10 月中旬果实进入成熟期。
2014 年,由于当地温度一直较低,树木休眠期较长。连续性高温出现在2 月23 日以后,有8 d 日均温大于10 ℃,3 月5 日才开始进入萌动期。在花序抽生出之前,其物候期晚于2013 年10 ~15 d,花序抽生于4 月20 日,均温达到19.7 ℃,而在出现花苞并进入初花期后,物候与2013 年极其相近,此时日均温大于22 ℃。由于2014 年5—6 月花期的(降雨量708 mm)当地雨水较2013 年(降雨量337 mm)多,最高一次降雨量达99 mm,雨水过多不仅会影响花粉活力及受精能力,使坐果不稳,而且使初花期时间比2013 年长,但从盛花期到果实膨大期两年物候则相近。果实初熟比2013 年迟25 d,由于2013 年9月温度大于26 ℃,而2014 年9 月温度下降到25 ℃以下,10 月温度回升26 ℃以上,果实才开始转色,一直到11 月初才成熟。本研究证明了福建建宁无患子连续两年的重要物候期极其相近,在连续出现8~10 d 日均温大于10 ℃后开始萌动,在大于18 ℃后开始抽生出花序,在大于22 ℃后进入初花期。对于2013 年萌动前活动积温为156.4 ℃,2014 年的活动积温为103.2 ℃,所以2014 年萌动期推迟;2014年9 月温度下降,也使成熟期比2013 年推迟,曾有研究证明[28]温度及降雨量对物候影响较大,本研究为之后根据物候期开展的研究奠定基础。
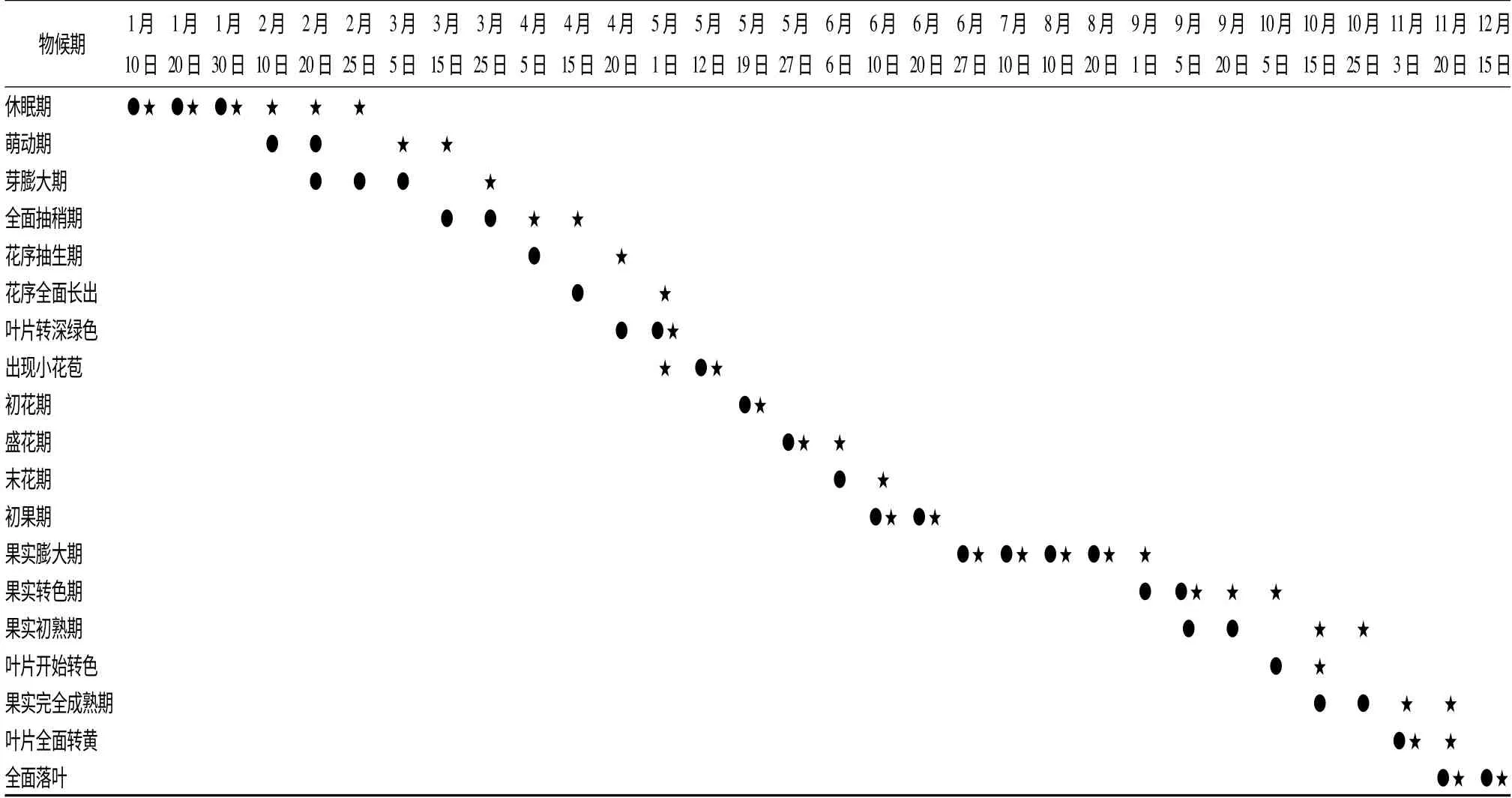
表1 2013 年1 月—2014 年12 月福建省建宁县无患子物候期
3.2 无患子开花、结果习性
无患子花为杂性,与同属无患子科的文冠果相似,属两性花,并有可孕花(雌)与不可孕花(雄)之分[29](见图1A、B)。可孕花具有三心皮(极少数为四心皮)组成的复雌蕊和8 枚不育的雄蕊,其花蕾后期发育过程中,花丝不再生长,各时期长短一致(见图1C、D、E),明显短于子房,花药不开裂,而子房却迅速膨大(见图1F),发育正常。不可孕花有8 枚可育的雄蕊,不同时期花丝持续伸长,花药开裂,正常散粉,到末期花丝逐渐萎蔫弯曲(见图1G、H、I),雌蕊很早退化、大小不变、逐渐萎缩(见图1J、K、L)。
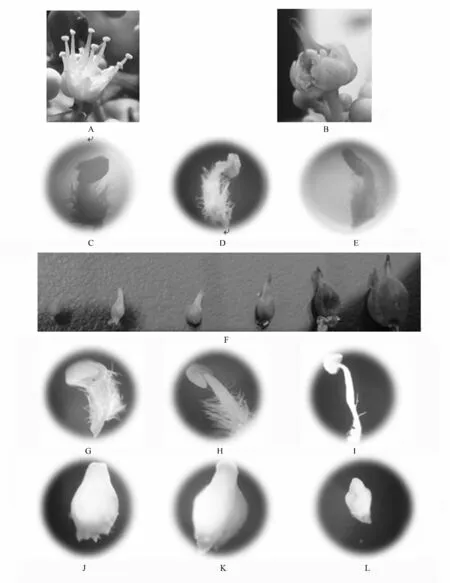
图1 可孕花及不可孕花各时期的雄蕊和子房变化
无患子的花在各时期的雌雄比例不一。开花顺序一般有两种情况,一种是不可孕花先开一部分,3~4 d 后先开的不可孕花凋落,可孕花开,2 d 后剩余的不可孕花全部开放(雄先行);第二种是可孕花先开1~2 d 后,不可孕花逐渐开放(雌先行)。同一花序上同时有可孕花和不可孕花,多为顶生花序,少数为侧生花序,侧生花序相对顶生偏小,但雌雄比与顶生相近。在初花期的雌雄花比例为5 ∶7,在盛花期的雌雄花比例为3 ∶8,末花期的雌雄花比例为25 ∶3。可孕花在3 个时期占全树花的比例分别为41%、27%和89%,不可孕花占59%、73%和11%(见图2)。一般以盛花期来说,可孕花占30%左右,不可孕花占70%左右。可孕花和不孕花在开花过程中形态及生理特征上存在一些差别,其各花期的花药、花丝、花瓣、子房等形态变化和开花动态见表2。

图2 无患子的开花动态
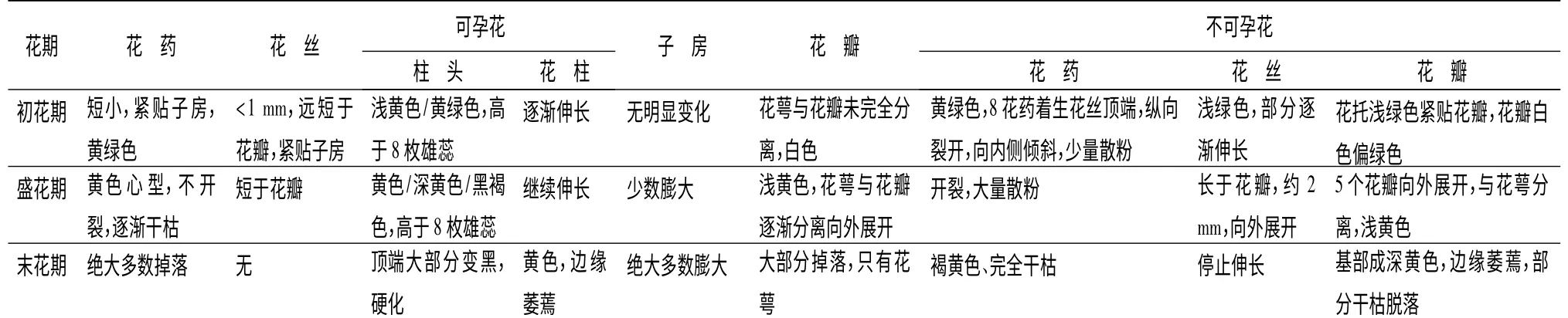
表2 无患子可孕花与不孕花形态特征和开花动态的比较
无患子花序似圆锥形,在盛花期的平均长度和宽度分别为(26.92±2.2)cm 和(26.40±1.6)cm。结果母枝上一般会有1~8 个结果枝不等,但当年能结果的不超过4 个,最多出现2 ~3 个结果枝,说明无患子顶端优势明显,在修剪时即可减掉多余的竞争枝,保证结果枝营养充分。无患子花序平均质量(77±5.3)g,一个花序最多结果100 个左右,平均结果17 个,每个花序的最高产量可达400 g,部分花序也有不结果的现象出现。
无患子为蒴果,在果实膨大期,复雌蕊子房多形成3 个分离小果,3 个果同时膨大,随着逐渐成熟,其中1~2 个果萎蔫,以供给一个果实营养;少数情况下会3 个果全成熟;极少数情况下出现4 心皮的复雌蕊出现4 个果完全成熟的情况(见图3)。成熟后,果实平均纵径、横径、侧径分别为(19.69±1.8)mm、(21.98±2.2)mm、(18.50±0.6)mm,果的最高质量可达7.5 g,平均质量4.5 g。偏大的无患子果多为心脏形,偏小的为近圆形。果皮厚度为(2.40±0.6)mm,其成熟前光滑、偏绿色,成熟后较粗糙、为黄色。果皮及种子平均质量分别为3.11 和1.75 g,各占整粒果实质量的69%和31%。种子平均纵径、横径分别为(14.85±0.3)mm、(14.38±0.8)mm,均为近圆形,且黑色光滑。
3.3 落花落果情况
无患子的开花数在初花期到盛花期间迅速增加,不可孕花数量在盛花期达到最高点,完成授粉后则迅速脱落,在盛花期到末花期间全部落完,同时伴随着少量的未能完成授粉受精的可孕花脱落(图4);可孕花在末花期到初果期间大部分脱落,整个花序的1/2 可孕花全部落下(表3)。坐果后从初果期到果实膨大期这段时期内落果率最高(表3),之后保持平稳状态,所以在初果期到膨大期期间采取保果措施可大幅度提高坐果率。

图3 无患子果实生长及分果败育情况
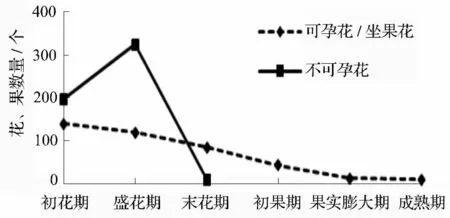
图4 无患子主要时期落花落果的变化
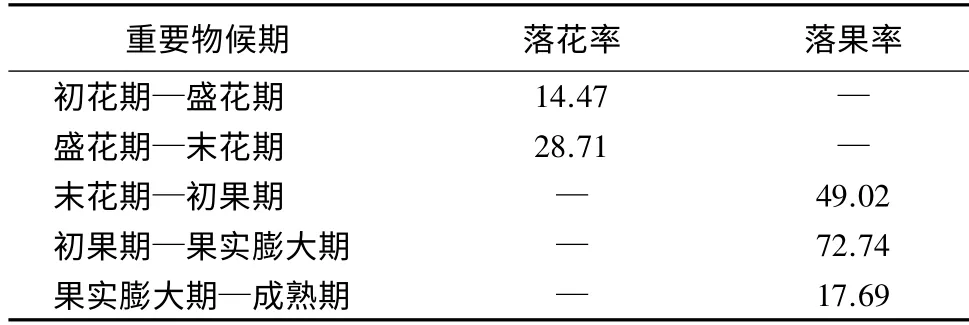
表3 无患子动态落果率 %
3.4 结果枝分类及对比
对无患子的结果枝进行分类,可知何种类型的结果枝可使无患子达到高产量的目的。笔者对90个当年结果枝进行测量并分析其长度与基径的相关性,采用K 均值聚类法进行了结果枝组的分类(表4)。将结果枝分成3 类,短果枝(10 ~34 cm)、中长果枝(35 ~56 cm)及长果枝(57 ~82 cm),其中不同长度的结果枝对应了相应基径,对其产量进行分析,长果枝的产量达到了极其显著水平。即说明了无患子是长枝和壮枝结果的树种。

表4 结果枝分类后产量间的对比
3.5 结果母枝基径与产量的关系
对无患子30 株大树的结果母枝进行调查,根据图4 可知结果母枝基径在<15 mm 时产量较低,多为零产,随着结果母枝基径变大,产量也随之上升,说明头年结果枝截除时需多留壮枝并进行短截以培养当年的结果枝,弱枝细枝可疏除。
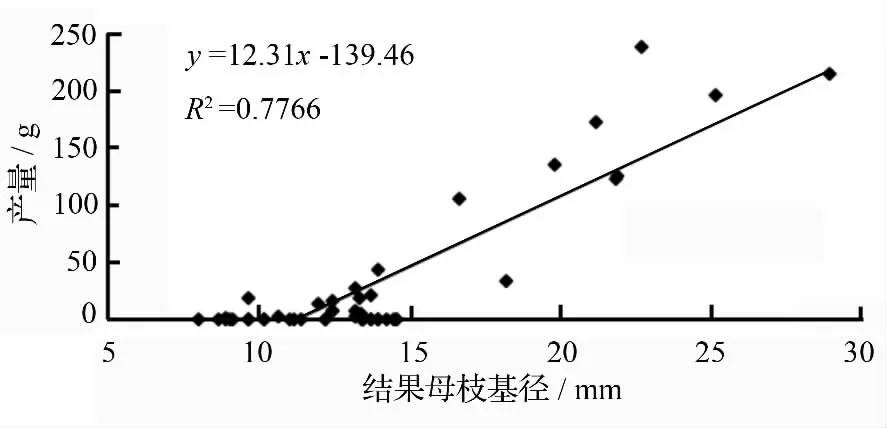
图5 结果母枝基径与产量关系
3.6 花序光照度与坐果率的关系
光照不够,影响产量,但光照过高,树体内部空,未形成叶幕(叶片在树冠内形成的叶群体),也不能较好的进行光合作用。所以有一个适合的光度值,在调节时,尽量在这个光照度之间即可。由图6 可知光照度(22.4~244)×100 lx,坐果率在0~29.09%;光照度(274~480)×100 lx,坐果率在27.27%~44.78%;光照度(524~625)×100 lx,坐果率为35.58%~78.1%;光照度(635~706)×100 lx,坐果率为16.18%~43.73%;光照度(729~861)×100 lx,坐果率7.02%~16.38%。无患子的若光照度调节好,坐果率可高达78.1%。
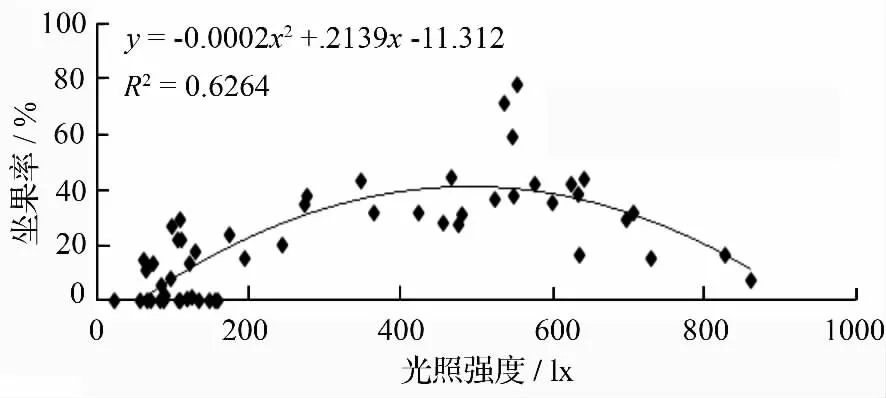
图6 光照强度与坐果率的关系
4 结论与讨论
无患子休眠期一般为40 ~65 d,在福建建宁的春季进入萌动期前需经历8~10 d 日均温大于10 ℃的天气,之后进入萌芽期。其花序抽生时日均温在18 ℃以上,花苞展开时日均温在22 ℃以上。刁松锋等[30]采用同为浙江天台种源种植于天台县的无患子开展了物候研究,其花序抽生于4 月20 日,发育约25 d,物候与笔者结果相近。花期分为初花期、盛花期、末花期等3 个时期,一般在花序抽生30~45 d 后进入初花期,花期持续15 ~20 d;果期分为初果期、果实膨大期、果实成熟期等3 个重要时期,花败后直接进入初果期,一直到果实成熟需(140±5)d。无患子落叶在果实成熟后20 ~45 d,之后便进入休眠期。连续2 a 的重要物候期几乎相近,重要时期相差±5 d;各重要物候期的气象因子(温度、湿度、降雨量等)指示一致,说明无患子的生长与环境变化存在重要关系。研究证明不同地区、不同年份文冠果的物候期有很大差异,即使在同一地区的相同生存环境下每年不同个体间的物候也有较大差异,均会对坐果率产生一定影响[31]。Blionis[32]等人研究证明决定植物开花物候的主要因素是温度,对相同植物而言,积温高的地方会更早开花。这种现象解释了福建建宁无患子开花物候较浙江天台[30]早5 ~10 d 的结论。也可证明由于2014 年(103.2 ℃)萌动前活动积温低于2013 年(156.4 ℃),所以其物候推迟。无患子在开展与物候期相关的各项科研工作时,可通过温度和降雨量的变化来预判其重要物候期。
无患子的花分为雄蕊败育的可孕花及雌蕊败育的不可孕花,两种花的花药与子房从花芽分化时期就形成差异。其开花过程包括初蕾—膨大—露白—待开—开放—转色—萎蔫—花落—坐果等,果园内不同树的开花期相距±3 d,同一株树也存在花期不遇的情况,所以授粉率低,是无患子坐果率低的原因之一。在盛花期其可孕花、不可孕花比例为3 ∶8,若进行及时调控,可增大可孕花的比例。
当地无患子的初花期正遇上当地雨季,对其开花授粉影响极大,可通过微气候调节、花果调控或基因筛选等措施提早或延迟花期。建宁当地种植黄花梨通过调节需冷量来使其提前开花;Raúl 等[33]使用乙烯延迟杏的花期等,有相应效果;Roux 等[34]对拟南芥的早花基因型进行筛选的方法效果显著,这些均值得借鉴。无患子落花落果现象明显,尤其在末花期及果实膨大期的落花落果率分别高达49%及73%,分析其原因是末花期大多数落不可孕花,使落花数量增大,但对果实产量不造成影响;果实膨大期为营养需求期,为给顶端果实提供营养,较多果实不能膨大则落果,使得落果率增大,尤其此段时期经一个子房形成的3 个果多数情况下会败育两个,若未得到及时补充会对后期无患子稳果造成更大影响。所以在开展花果调控等技术时,应及时采用微量元素补充、激素调控等措施进行稳果,可大幅度提升坐果率且可避免发生果实败育的现象。
无患子的结果枝组分为分成短果枝(10 ~34 cm)、中长果枝(35~56 cm)及长果枝(57 ~82 cm),其中长果枝的产量是其他的4 ~8 倍,显著高于其他结果枝,且越粗的结果母枝产量越高,说明在休眠季修剪时,应留短而粗结果母枝来培养次年或当年结果枝,需尽量疏掉短小和瘦弱的一年生枝。在选取嫁接枝条时也要选择当年生的长果枝,以保证成活率及产量。
光照对坐果率有着重要作用[35]。光照过高过低均对产量影响较大,所以无患子花序光照度在(524 ~625)×100 lx 时,坐果率最高,可以依据此光照度来指导修剪。依照适合无患子的开心形树形进行整形,在修剪时尽量使每个花序在同等条件下的光照度调节到此范围内。在生产中应保持合理的树冠间距,保留合理的结果母枝数量,使无患子树得到足够的光照度,达到优质高产的目标。
[1] 《国际石油经济》编辑部.洞察中国和印度的能源前景:国际能源署《世界能源展望2007》要点介绍[J].国际石油经济,2007,15(11):6-11.
[2] 韩德奇,徐国英,徐会林,等.生物柴油的现状与发展前景[J].国际石油经济,2002,10(5):34-36.
[3] 黄忠水,纪威,李淑艳,等.国外生物柴油的应用[J].节能与环保,2003,30(1):34-35.
[4] 田春龙,郭斌,刘春朝.能源植物研究现状和展望[J].生物加工过程,2005,3(1):15-19.
[5] 李军,吴平治,李美茹,等.能源植物的研究进展及其发展趋势[J].自然杂志,2007,29(1):22-25.
[6] 杨志斌,李德安.能源植物制备生物柴油的研究现状及发展趋势[J].湖北林业科技,2009,14(4):40-42.
[7] Makkar H P S,Becker K,Sporer F,et al. Studies on nutritive potential and toxic constituents of different provenances of Jatropha curcas[J]. Journal of Agricultural and Food Chemistry,1997,45(8):3152-3157.
[8] SujathaM,Mukta N. Morphogenesis and plant regeneration from tissue cultures of Jatropha curcas[J]. Plant Cell,Tissue and Organ Culture,1996,44(2):135-141.
[9] Zhang Y,Wang Y,Jiang L,et al. Aquaporin JcPIP2 is involved in drought responses in Jatropha curcas[J]. Acta Biochimica Etbiophysica Sinica,2007,39(10):787-794.
[10] Zhang Shuzhen,Yang Benpeng,Feng Cuilian,et al. Expression of the Grifola frondosa trehalose synthase gene and improvement of drought-tolerance in sugarcane (Saccharum officinarum L.)[J]. Journal of Integrative Plant Biology,2006,48(4):453-459.
[11] Yao Wei,Yu Aili,Xu Jingsheng,et al. Analysis and identification for transgenic sugarcane of ScMV-CP gene[J]. Molecular Plant Breeding,2003,2(1):13-18.
[12] Borrero M A V,Pereira J T V,Miranda E E. An environmental management method for sugar cane alcohol production in Brazil[J]. Biomass and Bioenergy,2003,25(3):287-299.
[13] Chhetri A B,Tango M S,Budge S M,et al. Non-edible plant oils as new sources for biodiesel production[J]. International Journal of Molecular Sciences,2008,9(2):169-180.
[14] Yang J,Dai G,Ma L,et al. Forest-based bioenergy in China:Status,opportunities,and challenges[J]. Renewable and Sustainable Energy Reviews,2013,18(3):478-485.
[15] 中国植物志编辑委员会.中国植物志:47 卷[M].北京:科学出版社,1998.
[16] 范理璋.无患子育苗技术[J].林业实用技术,2006,12(1):18-19.
[17] 陈小勤,黄林平,罗清连.滨海地区无患子芽苗切根移栽技术研究[J].林业实用技术,2003,9(3):10-11.
[18] 孙洁如,陈孔常,周鸣方,等.无患子表面活性物及其复配体系的性质研究[J].日用化学工业,2002,32(4):16-18.
[19] 黄素梅,王敬文,杜孟浩,等.无患子的研究现状及其开发利用[J].林业科技开发,2009,23(6):1-5.
[20] 孙颖,马桂珍.文冠果的生物学特性及其综合开发应用[J].现代农业,2010(1):4-5.
[21] 张谦,刘延刚,杨自强,等.文冠果的生物学特性和经济价值及其开发利用前景[J].农业科技通讯,2012(10):202-203.
[22] 李雪琴.建宁县无患子能源林引种育苗技术[J].绿色科技,2013(11):44-45.
[23] 李荣平,周广胜,张慧玲.植物物候研究进展[J].应用生态学报,2006,17(3):541-544.
[24] Barbola I F,Laroca S,Almeida M C,et al. Floral biology of Stachytarpheta maximiliani Scham. (Verbenaceae)and its floral visitors[J]. Revista Brasileira de Entomologia,2006,50(4):498-504.
[25] 戴攀峰,谭敦炎.雪莲的开花生物学特性及其生态适应意义[J].植物生态学报,2011,35(1):56-65.
[26] 严婷,郭军战,徐龙光.文冠果优良单株开花生物学特性研究[J].北方园艺,2014(5):58-62.
[27] 侯元凯,黄琳,周忠惠.文冠果果实性状相关性研究[J].林业科学研究,2011,24(3):395-398.
[28] 王连喜,陈怀亮,李琪,等.植物物候与气候研究进展[J].生态学报,2010,35(2):447-454.
[29] 王晋华,李凤兰,高荣孚.文冠果花性别分化及花药内淀粉动态[J].北京林业大学学报,1992,14(3):54-60.
[30] 刁松锋.无患子花果性状多样性及果实发育规律研究[D].北京:中国林业科学研究院,2014.
[31] 马利苹,王力华,阴黎明,等.乌丹地区文冠果生物学特性及物候观测[J].应用生态学报,2008,19(12):2583-2587.
[32] Blionis G J,Halley J M,Vokou D. Flowering phenology of Campanula on Mt Olynipos,Greece[J]. Ecography,2001,24(6):696-706.
[33] Grijalva C R L,Martínez D G,Macías D R,et al. Effect of ethephon on almond bloom delay,yield,and nut quality under warm climate conditions in northwestern mexico[J]. Chilean Journal of Agricultural Research,2011,71(1):34-38.
[34] Roux F,Touzet P,Cuguen J,et al. How to be early flowering:an evolutionary perspective[J]. Trends in Plant Science,2006,11(8):375-381.
[35] 李冬生,杨福新,孙万河.光照对日本栗树结实的影响[J].农业科技与装备,2009(5):21-22.
猜你喜欢
传感器世界(2022年7期)2022-10-12
湖北农业科学(2022年12期)2022-07-21
气象水文海洋仪器(2022年2期)2022-07-08
农产品市场周刊(2022年4期)2022-05-21
农产品市场周刊(2022年4期)2022-03-23
中南林业科技大学学报(2022年1期)2022-02-23
奥秘(创新大赛)(2021年2期)2021-04-07
山东林业科技(2019年2期)2019-06-03
江苏农业科学(2018年15期)2018-09-10
天然产物研究与开发(2018年7期)2018-08-21
