
Effects of the Amplitude and Frequency of Salinity Fluctuation on the Body Composition and Energy Budget of Juvenile Tongue Sole (Cynoglossus semilaevis)
2015-03-31 02:55:25SachinOnkarKHAIRNARTIANXiangliFANGZihengandDONGShuanglin
Sachin Onkar KHAIRNAR, TIAN Xiangli, FANG Ziheng, and DONG Shuanglin
Effects of the Amplitude and Frequency of Salinity Fluctuation on the Body Composition and Energy Budget of Juvenile Tongue Sole ()
Sachin Onkar KHAIRNAR, TIAN Xiangli*, FANG Ziheng, and DONG Shuanglin
,,,266003,
Effects of the amplitude (±2, ±4, ±6, and ±8) and frequency (2, 4, and 8d) of salinity fluctuation on the body composition and energy budget of juvenile tongue sole () were investigated in a 64-d experiment. Results showed that the amplitude and frequency of salinity fluctuation had significant interaction and both substantially affected the final weight and specific growth rate of juvenile tongue sole. The tongue sole exhibited better growth in treatments with moderate amplitude and frequency of salinity fluctuation (amplitude ±4–6; frequency 4–8d) than in other treatments and the control. In terms of energy budget, salinity fluctuation strongly affected the proportions of energy components, including those deposited for growth and lost in respiration, feces, and excretion. Moderately amplitude and frequency of salinity fluctuationg that favored the growth of tongue sole partitioned more energy for growth and less energy for metabolism than the constant and other amplitude and frequency of salinity fluctuation. Average energy budget for tongue sole at moderately fluctuating salinity was determined to be 100(food)=30.92(growth) +10.30(feces)+6.77(excretion)+52.01(respiration). Energetic advantage at moderately fluctuating salinity, including increased energy intake, high assimilation efficiency, reduced metabolism expenditure, and more energy partitioned into growth, might account for the enhancement of tongue sole growth. Commercial farmers are recommended to rear juvenile tongue sole with moderate salinity fluctuations for better growth performance of this species.
salinity fluctuations; fish body composition; energy budget; energy allocation;
1 Introduction
Tongue sole,Günther, 1873 (Actinopterygii, Cynoglossidae), is a rare fish species of great commercial value and native to China’s sea areas (Bohai Sea, Yellow Sea, and East China Sea), which inhabits coastal waters and estuaries (Masuda, 1984; Ma., 2005, 2007). Due to over-fishing, the wild population of tongue sole has decreased sharply,and its annual output was less than 1 ton (Jiang and Wan, 2005).In recent years, tongue sole has become an important species in China’s intensive indoor fish culture for its high nutritional and economic values (Ma., 2005, 2007).Most research of tongue sole hasfocused on the food habits and life history in Bohai Sea, as well as the breeding and embryonic development of this species (Dou, 1993,1995a,b; Du., 2004; Liu., 2005; Ma., 2005, 2007). Additionally, studies have investigated tongue sole its karyotype, genetic diversity(Liu., 2010; Zhuang., 2006; Liao., 2007; Liu., 2007, 2008), influence of water temperature and ration (Fang., 2010), and compensatory growth (Tian., 2010). However, more studies are appreciated for the quickly developing aquaculture of tongue sole as a newculture species.
Salinity is one of the most important abiotic factors, which affect the growth, body composition, and energy budget of aquatic organisms by exerting complex and wide-ranging biological effects (Brett and Groves, 1979; Gaumet., 1995; Kumlu., 2000; Boeuf and Payan, 2001). There are important commercial interests in determining the optimum salinity for each commercial fish species in intensive systems where the salinity can be altered to fit the species. A number of studies have been focused on the effect of constant salinity on aquatic organisms. It is generally accepted that aquatic organisms grow better at constant salinity rather than fluctuating salinity. Therefore, most aquatic organisms are cultured at constant salinity. The positive influence of fluctuating salinity on the growth of aquatic organisms has previously been reported in fish (Stroganov, 1962; Konstantinov and Martynova, 1993), but the associated effect on biochemical composition and energy budget of fish species is unclear. Regarding the biochemical compositions, protein, lipid, and ash contents of juvenile cobia () were found significantly affected by salinity (Chen., 2009); and protein, lipid, and moisture contents of juvenile starry flounder () were found significantly higher in low-salinity treat- ments than in other treatments (You., 2009).
All the energy acquired by animals through food ingestion is ultimately used in metabolic processes, deposited as new body tissues (growth or reproduction), or lost as waste products. Energy budgets are used to describe the partitioning of energy ingested into various components. Energy consumed in food () is primarily channeled into growth (), respiration (), feces (), and excretion (), following the expression=+++(Carfoot, 1987). Additionally, the energy values can be expressed as a ratio of absorbed or assimilated energy which helps to understand how energy is channeled to growth and which forms of energy are mainly consumed in a particular condition.
In this study, we examined the effects of the frequency and amplitude of salinity fluctuation on the biochemical composition and energy budget of tongue sole. Results were analyzed to reveal the energetic strategy of tongue sole, in order to provide reference data for promoting land-based culture of this species in brackish water.
2 Material and Methods
2.1 Fish Acclimation
Juvenile individuals of tongue sole () were provided by Yantai Mingbao Aquatic Product Co., Ltd. (Shandong, China) and transferred to the laboratory at the Aoshanwei Research Centre of Ocean University of China (Qingdao, China). Juvenile fish were acclimated to seawater (salinity 30) in 1000-L fiberglass tanks for atleast one week before experiment. During acclimation, the fish were fed to satiation twice a day at 07:00 and 18:00 with commercial pellets (Table 1; Guangzhou YuequnTechnology Co., Ltd, China).

Table 1 Energy content and chemical composition of commercial pellets for feeding juvenile tongue sole
Notes: Energy content (mean ± standard error) measured by PARR1281 Calorimeter (PARR Instrument Company, USA); protein content calculated as nitrogen content multiplying by 6.25 and nitrogen content determined with a Vario ELIII Elemental Analyser (Elementar, Germany); moisture content (mean ± standard error) determined by oven-drying at 70℃ to constant weight. Information of other chemical compositions provided by the manufacture (Guangzhou Yuequn Technology Co., Ltd, China).
2.2 Experimental Design and Management
The effect of salinity fluctuation on fish was studied at three frequencies and four amplitudes during a period of 64 days. The control treatment (S30±0) was maintained constant (30), and the experimental treatments (S30±2, S30±4, S30±6, and S30±8) were subjected to salinity fluctuation at the amplitudes of ±2,±4, ±6, and ±8, respectively (Fig.1). In each amplitude treatment, there were three frequency treatments (D2, D4, and D8) corresponding to 2-, 4-, and 8-d intervals of salinity changes, respectively. There were five replicates each treatment, and each replicate had six juvenile fish.
Fig.1 Example of salinity fluctuation treatment at the amplitude of ±6 and frequency of 4d (D4S30 ±6). The same fluctuation pattern was maintained at different amplitudes (±2, ±4, ±6, and ±8) and frequencies (2-, 4-, and 8-d).
At the end of acclimation, all fish were starved for 24h to evacuate the gut. Normal and healthy fish were selected and weighed, then assigned to 65 rectangular glass aquaria (55cm×30cm×35cm). Before weighing, fish were anaesthetized using MS-222 (60mgL−1) to reduce stress. Fifteen fish were sampled for estimation of the initial body composition and energy content of control treatment.
Initially on day 1, all fish were held at the control salinity (30) and subsequently salinity was changed as per experimental design. The fish were fed twice a day at 07:00 and 18:00 with commercial pellets described above. Before feeding, the feed of each aquarium was weighed (to 0.1mg). Uneaten feed in the tank was collected 30min after feeding by pipetting and then oven-dried at 75℃. Potential loss of uneaten food was determined by placing feed in water for 30min followed by collecting, drying, and weighing. The remaining proportion was calculated and used to adjust the amount of feed intake. For energy and nitrogen analyses, feces were collected by pipetting before and after feeding for 1h, oven-dried at 75℃, weighed, and homogenized, then stored at −20℃. During the experimental period, 30% of water in each aquarium was replaced every other day to ensure suitable water quality. Seawater used in the experiment was filtered with a sand filter. Low-salinity water was made by diluting sand-filtered seawater with fully aerated tap water, and high-salinity water made by adding sea salt to sand- filtered seawater. Moderate aeration was provided continuously to maintain the dissolved oxygen content (>6mgL−1); aeration was stopped 30min before fecal and uneaten food collection and resumed thereafter. Food consumption was estimated as the difference in dry weight between the amounts of food supplied and uneaten.
During the experiment, water temperature (18–22℃), pH (about 8.0), and ammonium-nitrogen content (<0.2mgL−1) were monitored daily and simulated natural photoperiod (14-h light: 10-h dark)was maintained. The experiment was conducted from September 27 to December 1, 2011. At the end of the 64-d experiment, all fish were weighed and sampled following 24-h starvation, then dried at 70℃.
2.3 Chemical Analysis
Moisture content of feed and fish was determined by oven-drying at 70℃ to constant weight. Ash content was determined by 10-h heating at 550℃ in a muffle furnace (SXL-1030, China) (Tian and Qin, 2003). Lipid content was measured with BUCHI Extraction System B-811 (BUCHI, Switzerland). Nitrogen content was determined with a Vario ELIII Elemental Analyzer (Elementar, Germany); and gross energy content measured by a PARR- 1281 Calorimeter (PARR Instrument Company, USA). Each sample had at least duplicate estimates. Protein was calculated as nitrogen content multiplying by 6.25.
2.4 Energy Determination and Energy Budget
Energy budget was calculated using the following equation (Carfoot, 1987):
,
whereis energy consumed;is energy for growth;is energy lost in feces;is energy loss as ammonia excretion; andis energy loss as respiration.,, andin the budget equation were calculated as follows:
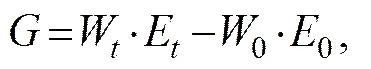
whereCandFare food intakes in terms of weight (g) and fecal production (g), respectively;IFandFEare energy contents of feed and feces (kJg−1), respectively;Wand0are the final and initial wet body weights (g) of fish, respectively; andEand0are the final and initial energy contents of fish body (kJg−1), respectively.was estimated based on a nitrogen budget equation (Elliott, 1976):
whereCis nitrogen consumed from food;Fis nitrogen lost in feces;Gis nitrogen deposited in animal body; and 24.83 is energy content of excreted ammonia (kJg−1).was calculated as the difference between energy consumption and the sum of energy allocated to growth, feces, and excretion:
Energy assimilated (), energy absorbed (), assimilation efficiency (1), net growth efficiency (2), and net respiratory efficiency (3) were estimated using the following equations (Cui, 1989):
2.5 Growth Rate Calculation and Data Analysis
Specific growth rate (SGR) in terms of weight (%d−1) was calculated as follows (Cui, 1989):
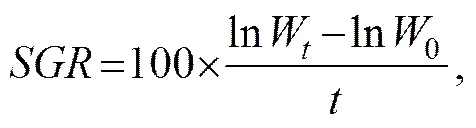
whereWand0are the final and initial wet body weights (g) of fish, respectively; andis the feeding duration (d).
Statistical analysis was performed with a statistical package (SPSS 16.0 for Windows, SPSS Inc., Richmond, CA, USA). The assumption of homogeneity of variances was tested for all data, and log-transformation was conducted if necessary. The interaction between the amplitude and frequency of salinity fluctuations, which affected the SGR rate, body composition, and energy budget of fish, was tested using two-way analysis of variance (ANOVA). Significant ANOVAs were followed by a Duncan’s multiple comparison to examine the differencebetween treatments.
3 Results
During the experiment, no mortality, health disturbance, or alteration in behavior of fish was observed in any treatments (0.05).
3.1 Growth
There was no siginificant difference in the intial body weight of tongue sole among all treatments (0.05, Table 2). The results of two-way ANOVA showed that both the frequency and amplitude of salinity fluctuation had significant effects on the final weight of juvenile fish (=10.366,0.01 and=40.401,0.01, respectively), and that there was significant interaction between these two parameters of salinity fluctuation (=3.906,0.01). Among all treatments, D8S30 ±6 obtained the maximum final body weight of fish, 19.42g (0.05).
The results of two-way ANOVA showed that the frequency and amplitude of salinity fluctuation significantly affected the SGR of fish (=206.126,0.01;=189.750,0.01), with significant interaction between the two parameters of salinity fluctuation (=4.614,0.05). The treatments with moderate salinity fluctuation (amplitude ±4–6; frequency 4–8d) produced significantly higher SGR of fish than the control did (0.05), whereas the other treatments mostly failed to improve the SGR of fish compared with the control (0.05).
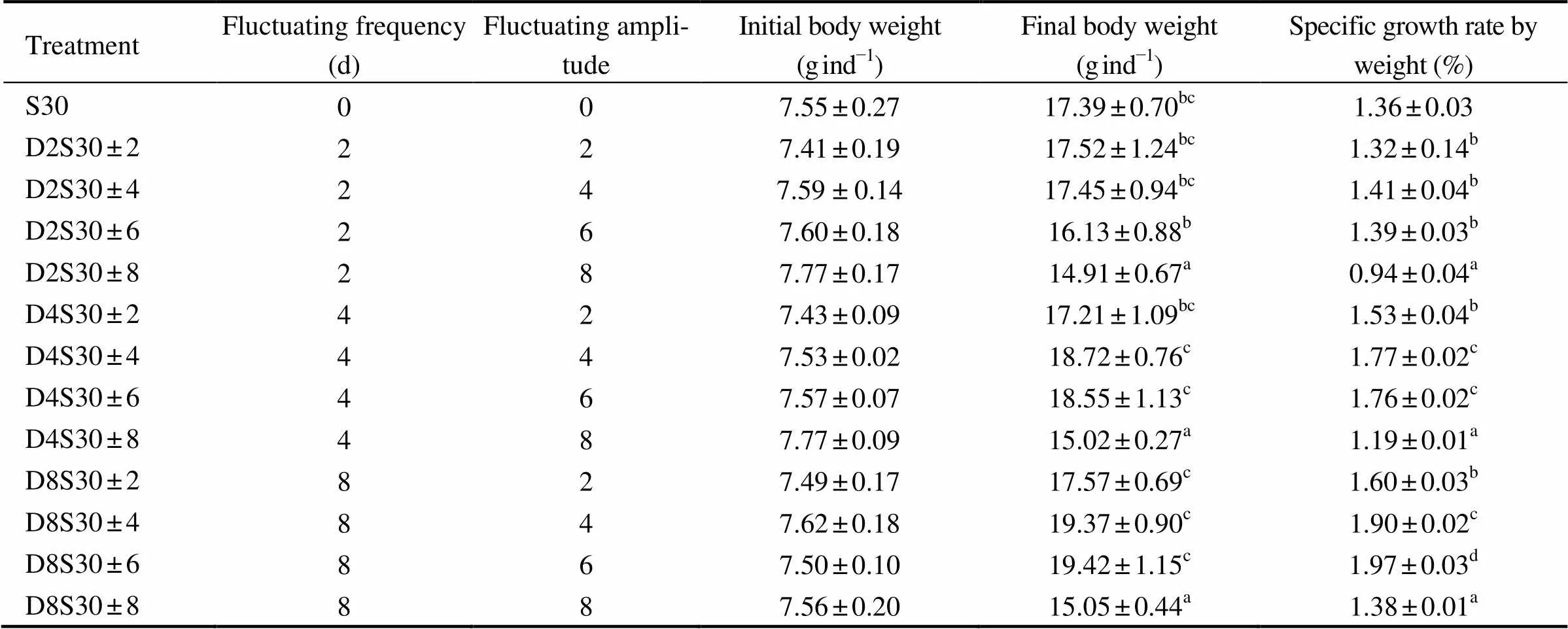
Table 2 Growth performance of tongue sole (C. semilaevis) at different frequencies and amplitudes of salinity fluctuation for 64d (mean ± standard error)
Note: Different superscripts in the same column mean significant difference between treatments (Duncan’s multiple comparisons,<0.05).
3.2 Body Composition
The content of crude protein, lipid and ash in tongue sole ranged from 22.63% to 24.95%, 3.87% to 5.35% and of 2.04% to 3.25% dry weight, respectively. They were significantly affected by the frequency and amplitude of salinity fluctuation (0.01). And there was significant interaction between the two parameters of salinity fluctuation (0.01 for protein and lipid contents;0.05 for ash content). Moisture content of juvenile fish ranged from 77.32% to 83.21%, which significantly affected by the frequency and amplitude of salinity fluctuation (0.05 and0.01, respectively). However, no significant interaction was found between the frequency and amplitude of salinity fluctuations (0.05) (Table 2).
Generally, the content of crude protein in fish showed increases with increasing frequency and amplitude (up to ±6) of salinity fluctuation, while lipid content of juvenile fish tended to decrease with increasing amplitude of salinity fluctuation (except for ±8). Salinity fluctuation at high frequency and large amplitude (D2S30±8) deposited significantly more lipid in fish than other treatments (0.05). Ash content of juvenile fish decreased with increasing frequency of salinity fluctuation. However, there was no significant difference in ash content of fish between the treatments with salinity fluctuation at amplitudes of ±4 and ±6 and frequencies of 4 and 8d (0.05). For moisture content of juvenile fish, there was a trend to decrease with increasing amplitude (except for ±8) of salinity fluctuations at each frequency.
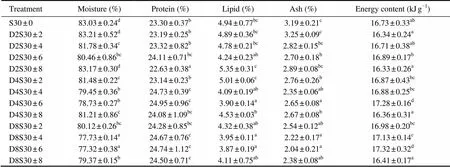
Table 3 Biochemical compositions of tongue sole (Cynoglossus semilaevis) at different frequencies and amplitudes of salinity fluctuation for 64d (mean ± standard error)
Notes: Different superscript letters in the same column mean statistically significant differences between treatments (Duncan’s multiple comparisons,<0.05). Lipid, protein, and ash contents are percentages of wet body weight.
The energy content of fish ranged from 16.33 to 17.32kJg−1and was significantly affected by the frequency and amplitude of salinity fluctuation (=124.208,0.01 and=255.592,0.01, respectively). There was significant interaction between the two parameters of salinity fluctuations (=2.307,0.05). Energy content of fish associated with salinity fluctuations at the amplitude of ±6 was significantly higher than that at ±8 at the frequency of 4d (0.05), and that associated with salinity fluctuations at the amplitude of ±8 was lowest among all treatments at the frequency of 8 d (0.05) (Table 3).
3.3 Energy Budget
The energy partitions for growth, feces, excretion, and metabolism were 24.67–65.01, 11.11–22.18, 12.61–19.61, and 97.50–134.57Jg−1d−1, respectively (Table 4).
The results of two-way ANOVA showed that most components of energy budget in juvenile fish were significantly affected by the frequency and amplitude of salinity fluctuation (0.05), and there was no significant interaction between two parameters (0.05) except for the energy for growth (=7.317,0.01). At the same frequency of salinity fluctuation, the energy for growth generally increased with increasing amplitude of salinity fluctuation from ±2 to ±6 but declined to the lowest level at ±8 (0.05); whereas the energy for excretion and metabolism was highest with salinity fluctuation at the amplitude of ±8 (0.05).
Energy allocation in juvenile tongue sole during the experimental period is shown in Table 5. The results of two-way ANOVA showed that the frequency and amplitude of salinity fluctuation significantly affected the pattern of energy allocation in juvenile fish. A large proportion of energy was allocated to growth and respiration. The proportion of energy allocated towards growth ranged from 13.02% to 32.96% and was significantly affected by the frequency and amplitude of salinity fluctuations (=104.140,0.05;=119.299,0.05, respectively). There was no significant interaction between the two parameters of salinity fluctuation (=1.983,=0.094).The proportion of food energy allocated to growth at lower frequency of salinity fluctuation was significantly higher than that at higher frequencyat the same amplitude of salinity fluctuation and the control (0.05). The proportion of food energy allocated to growth was significantly higher in D4S30±4, D4S30±6, D8S30±4, and D8S30±6 than in the other treatments and the control (0.05), while that at the amplitude of ±8 was significantly lower than that at other amplitudes at the same fluctuation frequency and the control (0.05).

Table 4 Energy parameters of tongue sole (Cynoglossus semilaevis) at different frequencies and amplitudes of salinity fluctuations for 64d (mean ± standard error, Jg−1d−1)
Notes:, energy consumedin food;, energy deposited for growth;, energy lost in feces;, energy in excretion; and, energy for metabolism. Different superscript letters in the same column mean statically significant differences between treatments (Duncan’s multiple comparisons,0.05).
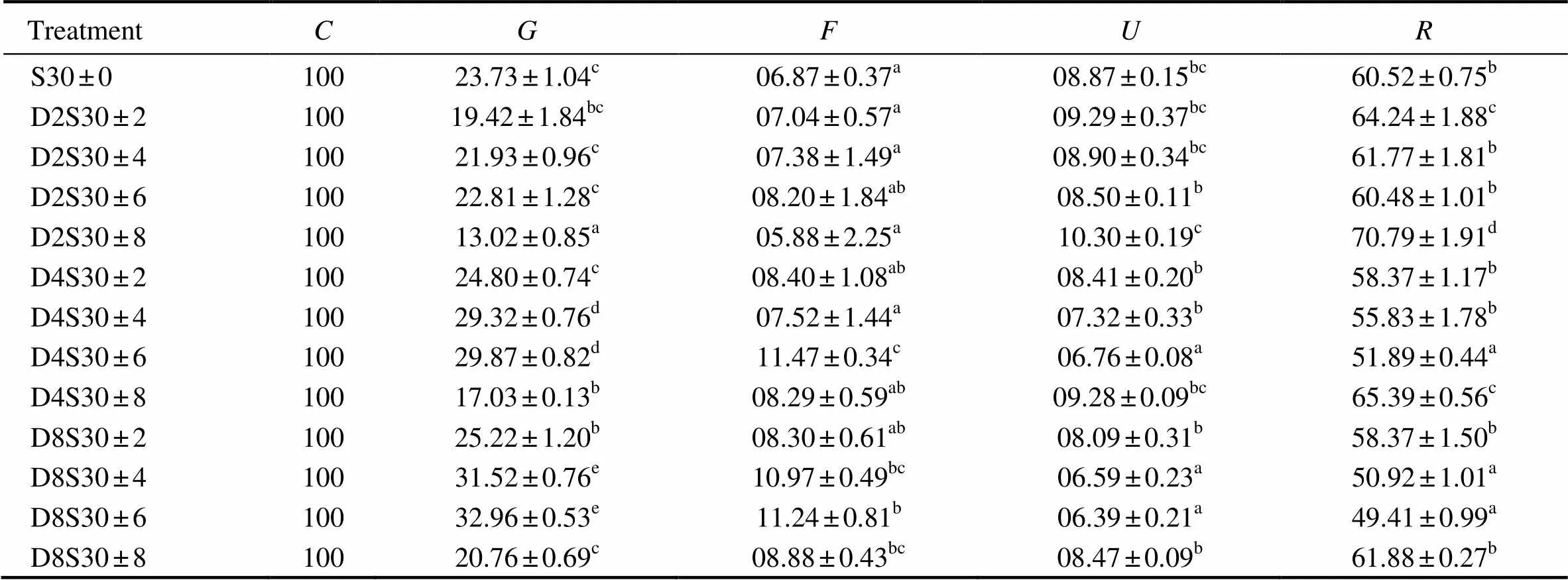
Table 5 Energy allocation of tongue sole (Cynoglossus semilaevis) at different frequencies and amplitudes of salinity fluctuations for 64d (mean±standard error, %C)
Notes: Mean values (± standard error,=4) were percentage of, energy consumedin food;, energy deposited for growth;, energy lost in feces;, energy in excretion; and, energy for metabolism.Different letters in the same column mean statistically significant differences between treatments (Duncan’s multiple comparisons,0.05).
The proportion of food energy allocated to feces was significantly affected by the frequency and amplitude of salinity fluctuation (=8.653,0.05 and=4.745,0.05, respectively) which had no significant interaction (=1.088,=0.388). The proportion of food energy allocated to feces was significantly higher in D4S30±6, D8S30±6, and D8S30±6 than in other treatments and the control (0.05).
The proportion of energy for excretion was significantly affected by the frequency and amplitude of salinity fluctuation (=85.036,0.01 and=65.234,0.01, respectively), and there was no significant interaction between these two parameters (=1.848,=0.117). The proportion of food energy allocated to excretion was significantly lower in D4S30±4, D4S30±6, D8S30±4, and D8S30±6 (0.05), but higher in D2S30±8 and D4S30 ±8 than in other treatments and the control (0.05).
The proportion of energy allocated for metabolism ranged from 49.41% to 70.79% and was significantly affected by the frequency and amplitude of salinity fluctuation (67.677,<0.01 and64.554,<0.01, respectively). There was no significant interaction between the two parameters of salinity fluctuation (1.545,0.05). The proportion of energy allocated for metabolism was significantly lower in D4S30±4, D4S30±6, D8S30±4, and D8S30±6 than in other treatments and the control (<0.05). Higher amplitude of salinity fluctuation (±4) significantly increased the proportion of energy allocated to metabolism compared with lower amplitude of salinity fluctuations at the same frequency (<0.05).
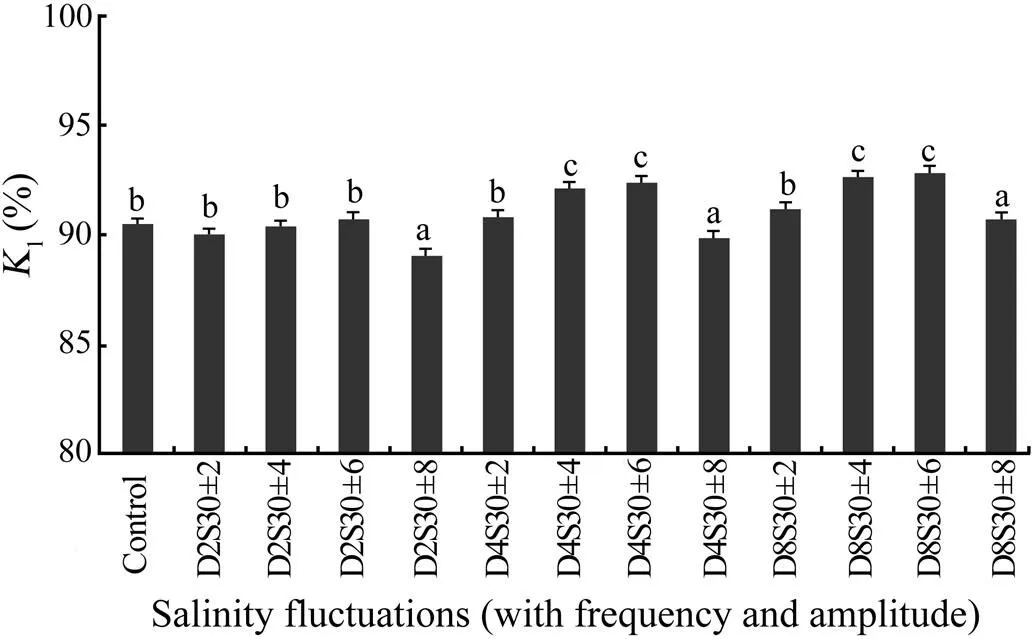
Fig.2 Assimilation efficiency (K1) of tongue sole reared at different fluctuating salinities for 64d. K1(%)=100×As/Abs=100×(G+R)/(G+R+U). Each bar represents the mean value (± standard error) of K1 for four replicates. Dissimilar letters above bars denote statistically significant differences in K1 between the treatments.
The assimilated efficiency showed sensitivity to salinity fluctuation (Fig.2). It was significantly higher in D4S30±4, D4S30±6, D8S30±4, and D8S30±6 (<0.05) but significantly lower in D2S30±8 and D4S30±8 than in other treatments and the control (<0.05). The percentage of assimilated energy channeled into growth (net growth efficiency) showed a similar tendency with the assimilated efficiency, whereas the net respiratory efficiency showed a reverse trend (Fig.3). Clearly, an increasing proportion of energy allocated to respiration decreased the energy assimilated into growth.
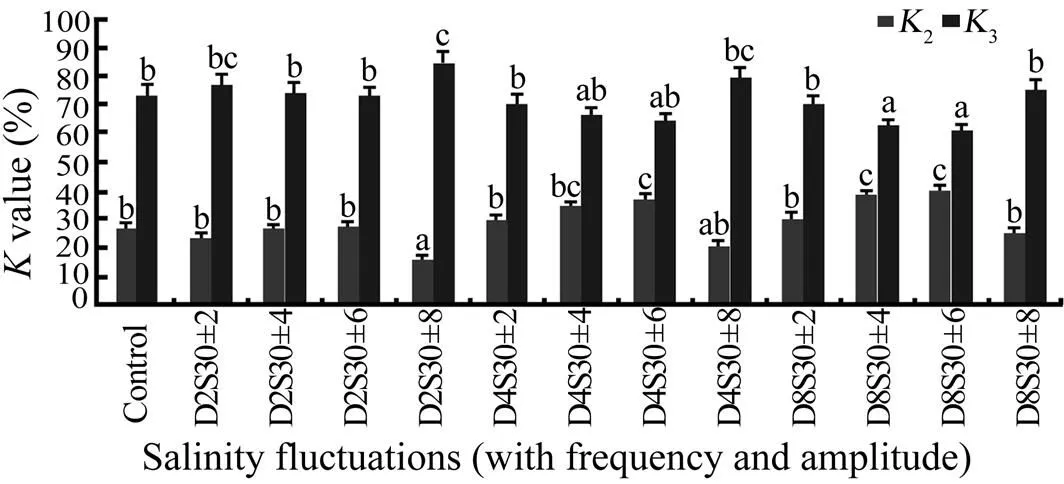
Fig.3 Net growth efficiency (K2) and net respiratory efficiency (K3) of tongue sole reared at different frequencies and amplitudes of salinity fluctuations for 64d. Data presented as the mean values (± standard error) of K for four replicates in each treatment. K2(%)=100×G/As=100×G/(G+R); and K3(%)=100×R/As=100×R/(G+R).
5 Discussion
Previously, researchers have mainly focused on the effect of constant salinity on aquatic organisms, and the idea that aquatic organisms grow better under constant salinity rather than fluctuating salinity is generally accepted. In fact, the positive influence of salinity fluctuation on the growth of aquatic organisms has been reported in Russian sturgeon () (Stroganov, 1962), common carp (), and white amur () (Konstantinov and Mar- tynova, 1993). Salinity fluctuation over the range of 0–2 considerably accelerated the growth of the above mentioned fish species in comparison with fresh water or water at the salinity of 2. Tongue sole is an euryhaline fish species which can adapt to a wide range of salinities (0–40) in a short period (Tian., 2011). To our knowledge, the present study represented the first work to investigate the effect of salinity fluctuation in terms of amplitude and frequency on the growth and bioenergetics of juvenile tongue sole.
Despite extensive studies on the energy budget of fish, their conclusions have not reached a consensus. Brett and Groves (1979) provided a general energy budget for carnivores and herbivores as follows: Carnivores, 100=29+20+7+44;and Herbivores, 100=20+41+2+37. Cui and Wootton (1988) hypothesized a constant pattern of energy allocation based on the study of a freshwater fish species, minnows (), and speculated that the pattern of energy allocation might be generally constant and independent of temperature at maximum rations. However, this hypothesis is not supported by the majority of studies on marine fish, such as red seabream (), black porgy (),(Sun, 1999, 2001, 2003), and juvenile cobia () (Sun., 2006). Additionally, Tang. (2003) established a uniform energy budget of seven marine fish species as 100=25.3+3.6+7.8+63.3. However, more than a constant equation, the energy budget of fish may be affected by a variety of biotic and abiotic factors. In the present study, the energy budget for juvenile tongue sole, a marine fish species, at constant salinity of 30 was determined to be 100=23.73+6.87+8.87+60.52, similar to the result of Tang. (2003). However, the pattern of energy allocation was significantly affected by the frequency and amplitude of salinity fluctuations, with no significant interaction between two parameters. For juvenile tongue sole reared at fluctuating salinities, the proportion of food energy assimilated to growth was 13.02%–32.96% and that spent in metabolism was 49.41%–70.79%. Likewise, the average energy budget which favored the growth of juvenile tongue sole at moderately fluctuating salinities was 100=30.92+10.30+6.77+52.01. Compared with those at constant salinity, the proportion of food energy assimilated to growth of fish increased significantly while the percentage of energy expenditure for respiration decreased significantly at fluctuating salinities. These observations mean that there exists an energetic advantage for juvenile tongue sole reared at suitable salinity fluctuations.
In the present study, both the frequency and amplitude of salinity fluctuation imposed a significant effect on the SGR of tongue sole, and significantly higher SGR of fish was observed at moderately fluctuating salinity (amplitude ±4–6; frequency 4–8d) than at constant salinity (30). However, higher fluctuation amplitude (8) or lower frequency (2d) of salinity fluctuation showed no positive influence on the growth of tongue sole. On the contrary, high salinity fluctuation reduced the scope for growth. The SGR of fish was significantly lower at the amplitude of 8 and the frequency of 2d than at the constant salinity of 30. Thus, the positive or negative effect of salinity fluctuations on the growth rate of tongue sole is dependent on not only the amplitude but also the frequency of salinity fluctuations. Several investigations have explored the factors which may enhance the growth of aquatic animals at fluctuating salinities(Marangos., 1989; Feng., 2008), but the results remain discrepant. A study showed that assimilation efficiencyofjuveniles was the highestat fluctuating salinities (amplitude ±5; frequency 4d) compared to that at constant salinity (20) or in other treatments; this result coincided with the growth pattern ofshrimp (Feng., 2008). When energy allocation of tongue sole at different salinity fluctuations was considered, excessive high or low water salinity fluctuations resulted in lower energy retention in tongue sole. This could be attributed to increased energy expenditure for compensating osmoregulation and ionic regulations. Contrarily, the fluctuating salinities favorable to the growth of juvenile tongue sole partitioned more energy for growth and less energy for metabolism than constant salinity and other fluctuating salinities.
In conclusion, moderate salinity fluctuation at the amplitudes of ±4–6and frequencies of 4–8d favored the growth of juvenile tongue sole. Energetic advantages at moderately fluctuating salinity, including increased energy intake, high assimilation efficiency, reduced metabolism expenditure, and more energy partitioned into growth, might account for the enhancement of fish growth. Commercial farmers engaged in tongue sole aquaculture are recommended to culture tongue sole indoor or in ponds adjacent to estuaries or creeks, where they can take the direct benefit of tidal salinity fluctuations for achieving the fastest growth of fish. This study provides reference data for farm site selection and salinity maintenance, thus having implications for maximizing commercial productivity in aquaculture of tongue sole.
Acknowledgements
This research was supported by the National Great Project of Scientific and Technical Supporting Programs of China (Grant No. 2011BAD13B03), the Program for Excellent Youth Foundation of Shandong province (Grant No. JQ201009), and the Public Science and Technology Research Funds Projects of Ocean, State Oceanic Administration of the People’s Republic of China (Grant No. 200905020).
Boeuf, G., and Payan, P., 2001. How should salinity influence fish growth?,130C: 411-423.
Brett, J. R., and Groves, T. D. D., 1979. Physiological energetics. In:. Hoar, W. S., and Randall, D. J., eds., Academic Press, New York, 279-352.
Carfoot, T. H., 1987.. Academic Press, New York, 407-515.
Chen, G., Wang, Z., Wu, Z., and Gu, B., 2009. Effects of salinity on growth and energy budget of juvenile cobia,., 40 (1): 374-382.
Cui, Y., 1989. Bioenergetics of fishes: Theory and methods., 13 (4): 369-383(in Chinese with English abstract).
Cui, Y., and Wootton, R. J., 1988. Pattern of energy allocation in the minnow,(L.) (Pisces: Cyprinidae)., 2 (1): 57-62.
Dou, S., 1993. Food habits and seasonal variation of stomach contents of tongue sole(Günther) in the Bohai Sea., 11 (1): 89-96.
Dou, S., 1995a. Life history cycles of flatfish species in the Bohai Sea, China., 34 (1-3): 195-210.
Dou, S., 1995b. Food utilization of adult flatfish co-occurring in the Bohai Sea of China., 34 (1-3): 183-193.
Du, W., Meng, Z. J., Xue, Z. Y., Jiang, Y. W., Zhuang, Z. M., and Wan, R. J., 2004. Embryonic development ofand its relationship with incubation temperature., 11 (1): 48-52 (in Chinese with English abstract).
Elliott, J. M., 1976. Energy losses in the waste products of brown trout (L.)., 45: 561-580.
Fang, J., Tian, X., and Dong,S. L., 2010. The influence of water temperature and ration on the growth, body composition and energy budget of tongue sole,., 299: 106-114.
Feng, C. M., Tian, X., Dong, S. L., Su, Y. P., Wang, F., and Ma, S., 2008. Effects of frequency and amplitude of salinity fluctuation on the growth and energy budget of juvenile., 39 (15): 1639- 1646.
Gaumet, F., Boeuf, G., Severe, A., Le Roux, A., and Mayer- Gostan, N., 1995. Effects of salinity on the ionic balance and growth of juvenile turbot., 47: 865- 876.
Jiang, Y. W., and Wan, R. J., 2005. Tongue sole () culture techniques. In:. Lei, J. L., ed., China Agricultural Press, Beijing, 663-665.
Konstantinov, A. S., and Martynova, V. V., 1993. Effect of salinity fluctuations on energetics of juvenile fish., 33: 1-8.
Kumlu, M., Eroldogan, O. T., and Aktas, M., 2000. Effect of temperature and salinity on larval growth, survival and development of.,188: 167- 173.
Liao, X., Shao, C. W., Tian, Y. S., and Chen, S. L., 2007. Poly- morphic dinucleotide microsatellites in tongue sole ().,7 (6): 1147-1149.
Liu, X. Z., Zhuang, Z. M., Ma, A. J., Chen, S. Q., Sun, Z. Z., Liang, Y., and Xu, Y. J., 2005. Reproductive biology and breeding technology ofGüther., 26 (5): 7-14 (in Chinese with English abstract).
Liu, Y. G., Sun, X. Q., Gao, H., and Liu, L. X., 2007. Micro- satellite markers from an expressed sequence tag library of half-smooth tongue sole () and their application in other related fish species., 7 (6): 1242-1244.
Liu, Y. G., Bao, B. L., Liu, L. X., Wang, L., and Lin, H., 2008. Isolation and characterization of polymorphic microsatellite loci from RAPD product in half-smooth tongue sole () and a test of cross-species amplification., 8 (1): 202-204.
Liu, Y. G., Gao, H., Liu, C. Y., Li, F. Z., and Chen, S. L., 2010. Genetics features of natural and cultured populations of half- smooth tongue sole () revealed by RAPD markers.,62 (2): 96-104.
Ma, A. J., Liu, X. Z., Xu, Y. J., Liang, Y., Zhuang, Z. M., Zhai, J. M., and Li, B., 2005. Study on feeding behavior and growth of tongue solein early development stage.,36 (2): 130-138 (in Chinese with English abstract).
Ma, A. J., Wang, X. A., Zhuang, Z. M., and Liu, X. Z., 2007. Study on relationship of the special sense organ and feeding behavior ofGüther., 38 (3): 240-245 (in Chinese with English abstract).
Marangos, C., Brogren, C.H., and Alliot, E.,1989. The influence ofwater salinity on the free amino acid concentrationin muscle and hepatopancreas of adult shrimps,.,17: 589-594.
Masuda, H., Amaoka, K., Araga, C., Uyeno, T., and Yoshino, T., 1984.. Tokai University Press, Tokyo, 1437pp.
Sun, L., Chen, H., and Huang,L.,2006. Effect of temperature on growth and energy budget of juvenile cobia ().,261: 872-878.
Sun, Y., Zhang, B., Guo, X., Wang, J., and Tang, Q., 1999. Effects of temperature on energy budget of., 20 (2): 54-59 (in Chinese with English abstract).
Sun, Y., Zhang, B., and Tang, Q., 2001. Effects of temperature on energy budget of., 21 (2): 186-190 (in Chinese with English abstract).
Sun, Y., Zheng, B., Zhang, B., and Tang, Q., 2003. Effect of temperature on energy budget of a seafish Species,., 25: 196-201 (in Chinese with English abstract).
Stroganov, N.S., 1962.. Moscow University Press, Moscow, 420pp.
Tang, Q., Sun, Y., and Zhang, B., 2003. Bioenergetics models for seven species of marine fish., 27 (5): 443-449 (in Chinese with English abstract).
Tian, X., Fang, J., and Dong, S., 2010. Effects of starvation and recovery on the growth, metabolism and energy budget of juvenile tongue sole,., 310: 122-129.
Tian, X., Wang, G., and Dong, S.,2011. Effect of salinity on plasma osmolality and gill Na+/K+-ATPase activity of the tongue sole ().,35: 27-31 (in Chinese with English abstract).
You, H. Z., Yang, Z. Q., Ge, S. G., and Jiang, Z. Q., 2009. Effect of different salinity on biochemical composition and energy budget forjuveniles., 185 (5): 16-19 (in Chinese with English abstract).
Zhuang, Z. M., Wu, D., Zhang, S. C., Pang, Q. X., Wang, C. L., and Wan, R. J., 2006. G-banding patterns of the chromosomes of tonguefish,Günther, and 1873., 22 (5): 437-440.
(Edited by Qiu Yantao)
DOI 10.1007/s11802-015-2400-z
ISSN 1672-5182, 2015 14 (1): 127-134
© Ocean University of China, Science Press and Spring-Verlag Berlin Heidelberg 2015
(May 14, 2013; revised August 1, 2013; accepted November 19, 2014)
* Corresponding author. Tel: 0086-532-82032117 E-mail: xianglitian@ouc.edu.cn
 Journal of Ocean University of China2015年1期
Journal of Ocean University of China2015年1期
- Journal of Ocean University of China的其它文章
- A Comparative Study of CART and PTM for Modelling Water Age
- Numerical Study of the Secondary Circulations in Rip Current Systems
- Effect of Anthracene on the Interaction Between Platymonashelgolandica var. tsingtaoensis and Heterosigma akashiwoin Laboratory Cultures
- Application of Numerical Simulation for Analysis of Sinking Characteristics of Purse Seine
- The Structure and Formation Mechanism of a Sea Fog Event over the Yellow Sea
- Molecular Cloning, Expression Pattern, and 3D Structural Prediction of the Cold Inducible RNA - Binding Protein (CIRP) in Japanese Flounder (Paralichthys olivaceus)