
日本囊对虾高位精养池浮游生物演替规律
2015-03-19 06:44李义军许文军王庚申陈兆明孙成波
安徽农业科学 2015年33期
李 婷,李义军,许文军,王庚申,陈兆明,孙成波*
(1.广东海洋大学水产学院,广东湛江524088;2.海南省昌江南疆生物技术有限公司,海南三亚572000;
3.浙江省海洋水产研究所,浙江舟山316021)
在养殖水生态环境中,浮游生物是水生态系统中能量供应和物质循环的重要环节,对构建稳健平衡的生态系统起着重要的作用。目前,国内外关于凡纳滨对虾、斑节对虾高位池、淡化养殖池的浮游生物群落结构的研究较多。张汉华[1]等对盐度为12.28‰~29.60‰的海水高位精养虾池浮游微藻种类的进行了调查。吴斌和廖思明[2]研究了广西北海凡纳滨对虾养殖池塘的微藻群落结构及多样性。杨兰秀[3]等报道了盐碱地封闭式对虾养殖池中浮游动物的群落结构及其演替规律。查广才等[4]分析了凡纳滨对虾淡化养殖池微型浮游生物群落及多样性。王丽卿等[5]调查了河口区斑节对虾淡化养殖塘浮游生物状况。但是,对日本囊对虾养殖池的浮游生物群落现状却未曾报道。笔者对日本囊对虾虾池中浮游生物种类、数量进行了整个养殖周期的监测,研究日本囊对虾养殖水体中的微型浮游生物群落组成及多样性的基本特征和动态,以期为后期水质调控及日本囊对虾高产养殖提供依据。
1 材料与方法
1.1 试验材料 试验在海南省昌江南疆生物技术有限公司板桥对虾养殖基地进行。试验池4口,面积各0.28 hm2,采用高位池养殖模式进行养殖日本囊对虾,池底铺地膜,其上覆盖30~50 cm细沙。所有养殖用水先经沙滤井过滤,然后采用有效氯为50 g/m3的漂白粉进行消毒。
日本囊对虾虾苗购自厦门,放苗时间为2010年8月16日,体长约4.47 mm,放苗密度为97.5万尾/hm2。放苗时水深1.2 m,30 d内逐渐加水到1.5 m,设4台功率为0.75 kW水车式增氧机增氧,养殖期间投喂恒兴牌人工配合饲料。
1.2 样品采集 试验中的月龄指的是从放苗起的养殖月龄。将虾池从塘壁到池中心均匀的分为3个区域,从放苗当天起进行第1次取样,此后间隔30 d取样1次,取样时间为早晨8:00,用2.5 L有机玻璃采水器分别在3个区域内任取3个位置各取中上层水样1份,混匀,取1 L,鲁哥氏液固定沉淀24 h以上,沉淀浓缩至100 ml甲醛保存,取5 L倒入浮游生物捕捉网过滤于50 ml小瓶内,然后用福尔马林和鲁哥氏液固定。
用2.5 L有机玻璃采水器分别在3个区域内选取3个方向分别取下层水样,用于水质理化因子检测。
1.3 样品的分析
1.3.1 浮游植物、原生动物和轮虫的定量。将定量样品摇匀,取0.1 ml样品置于计数框中,在显微镜下计数。浮游植物用鲁哥氏液固定,计数100个视野;原生动物、轮虫整片计数。
1.3.2 其他浮游动物的定量。将定量样品摇匀,取1 ml样品置于计数框中,在显微镜下整片计数。
1.3.3 浮游生物鉴定。浮游植物鉴定[6]通过活体观察及鲁哥氏液固定进行,浮游动物鉴定[7]通过活体观察进行。
1.3.4 理化因子测定。无机磷含量的测定采用磷钼蓝法;氨氮含量的采用奈氏比色法;硝酸氮含量的用锌镉还原法;亚硝酸氮含量的采用重氮-偶氮比色法;COD含量的采用酸性高锰酸钾法。试剂配制及测定步骤均参照雷衍之[8]的方法。
1.4 浮游生物的多样性分析 浮游生物的多样性指数和优势度分别采用Shannon-Weaner多样性指数H′和Simpson优势集中性指数D[9]来分析。
式中,Pi=Ni/N,Ni为物种i的个体数;N为群落样本个体总数;Pi为第i种个体数占总个体数的比例;S为群落中物种数。
2 结果与分析
2.1 日本囊对虾精养池营养盐的周期变化
2.1.1 虾池氮磷营养盐的周期变化。从图1可以看出,养殖过程中4个虾池的无机磷浓度变化趋势基本一致,随着养殖的进行,无机磷含量逐渐上升,到2月龄达到顶峰,此后随着浮游植物的大量繁殖无机磷含量下降,仅4号虾池在5月龄无机磷含量直线上升达到(0.139±0.037)mg/L,其他时期各虾塘无机磷的浓度范围为0.01~0.06 mg/L。
从图2~3可以看出,养殖过程中,4个虾池的亚硝酸氮、硝酸氮浓度变化趋势均基本一致,随着养殖的进行,饵料、对虾代谢物等逐渐沉积,亚硝酸氮、硝酸氮的浓度逐渐上升,到3月龄达到顶峰,此后浓度下降。
从图4可以看出,养殖过程中,4个虾池的氨氮浓度变化趋势基本一致,1月龄氨氮浓度较初放苗时明显降低,此后波动式的缓慢上升,3号和4号虾池的氨氮含量自4月龄后呈直线上升,在5月龄达到峰值(0.143±0.030)和(0.166±0.014)mg/L。
2.1.2 虾池氮磷比的周期变化。从图5可以看出,养殖过程中4个虾池2月龄前的变化趋势是相近的,均为1月龄氮磷比下降,2月龄上升,此后各月龄4个虾池的变化均有不同。1号虾池自1月龄起氮磷比呈上升趋势,5月龄达到最大值(4.732)。2号虾池自2月龄起氮磷比呈下降趋势,4月龄后有所上升。3号虾池氮磷比的变化曲线呈波浪变化,3月龄氮磷比突增至4.675,此后回落,4月龄后又上升。
2.2 日本囊对虾精养池浮游植物群落结构
2.2.1 日本囊对虾精养池浮游植物种类组成。由表1可知,4口虾池共观测到浮游植物7门26种,其中蓝藻门5种,绿藻门7种,硅藻门7种,裸藻门1种,隐藻门2种,甲藻门2种,黄藻门2种。种类最多的为绿藻门和硅藻门,各占种类数的26.9%,其次为蓝藻门,占19.2%,隐藻、黄藻和甲藻各占7.7%,裸藻占3.8%。养殖前中期,绿藻、硅藻多为优势种,蓝藻、隐藻及裸藻多为常见种和少见种;养殖后期,蓝藻、硅藻和绿藻多为优势种,隐藻、甲藻及裸藻的种类多为常见种和少见种。
2.2.2 养殖全过程日本囊对虾精养池浮游植物种类数量变化。从图6可以看出,4个虾池浮游植物种类数量变化趋势均是养殖初期种类相对较少,随着养殖时间的延长,种类增多,养殖后期浮游植物种类数量达到高峰。不同虾池由于底质、水质调控方式不同,藻类组成和变化不同。1号和3号虾池种类数量相对稳定,最为丰富,且3号虾池后期种类较丰富;2号虾池种类数量逐渐增加;4号虾池种类数量在养殖2月龄时出现低谷,此后又逐渐增加。4号虾池2月龄时出现的低谷现象,与该月在该池试用循环水机有关,循环水机中的滚筒过滤装置将部分浮游生物过滤出来,形成了虾池中浮游植物种类数量减少的现象。

表1 日本囊对虾精养池浮游植物的种类组成
2.2.3 高位池养殖日本囊对虾虾池浮游植物优势种演替动态。从图7可以看出,1号虾池浮游植物优势种为绿藻门的小球藻、圆形鼓藻、亚心形扁藻以及硅藻门的变异直链藻和蓝藻门的最小胶球藻,共计5个种。每次水样中,一般有2~3个优势种,优势种越少,其优势度越高。2号虾池浮游植物优势种为绿藻门的小球藻、圆形鼓藻、包氏卵囊藻及亚心形扁藻、硅藻门的三角褐指藻和变异直链藻以及蓝藻门的最小胶球藻、念珠藻和小颤藻,共计9种。早期优势种少,以小球藻为主,优势度高,后期优势种增多,以颤藻为主,颤藻主要出现在底质淤泥较深的老化虾池。3号虾池浮游植物优势种为绿藻门的小球藻、圆形鼓藻和包氏卵囊藻以及硅藻门的牟氏角毛藻和变异直链藻和黄藻门的拟气球藻以及蓝藻门的最小胶球藻和小颤藻,共计8种。前中期小球藻仍处于主导地位,优势度高,但后期则变化为小球藻与小颤藻、最小胶球藻等蓝藻共为优势种。
4号虾池浮游植物优势种为绿藻门的小球藻、圆形鼓藻以及硅藻门的变异直链藻和中华半管藻与隐藻门的尖尾蓝隐藻以及蓝藻门的最小胶球藻和小颤藻。小球藻的优势度随着养殖时间的延长逐渐降低,最终被蓝藻、隐藻等替代,颤藻从中后期开始成为优势种,优势度逐渐增大。
2.2.4 高位池养殖日本囊对虾虾池浮游植物多样性。由表2可知,4个日本囊对虾虾池浮游植物多样性指数的变化情况基本一致,表现为随着养殖的进行,饲料、虾代谢物等营养物质的增多,养殖1月龄的浮游植物多样性指数较初放苗水体高,2月龄由于虾类、浮游动物等的摄食作用多样性指数有所降低,此后随着养殖时间的延长,虾池浮游植物的多样性逐步增加,群落结构趋于复杂和稳定。浮游植物群落优势集中性指数的总体规律与多样性相反,早期优势度高,后期降低,最明显的是1号、2号和4号虾池,说明早期优势种比较突出。
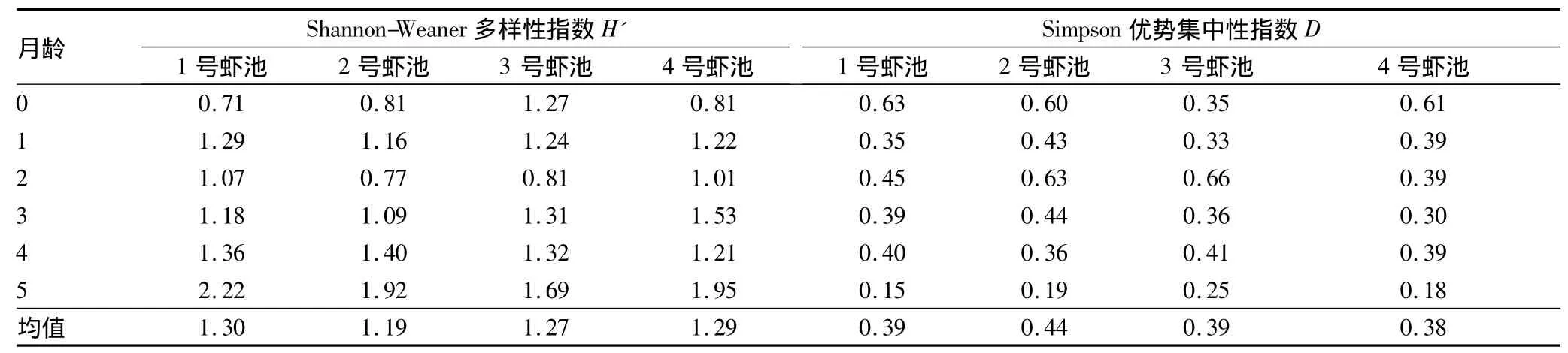
表2 日本囊对虾精养池浮游植物的群落多样性
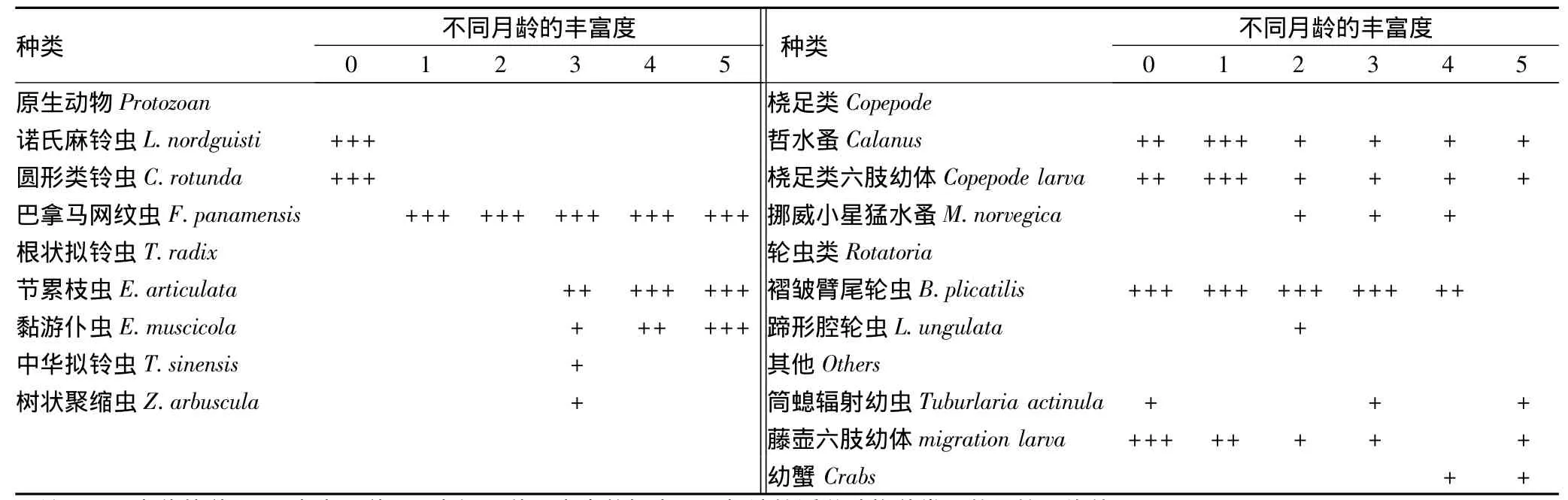
表3 日本囊对虾精养池浮游动物的种类组成
2.3 日本囊对虾精养池浮游动物群落结构
2.3.1 日本囊对虾精养池浮游动物种类组成。由表3可知,4口虾池观测到的浮游动物有:原生动物8种,分别为诺氏麻铃虫、类铃虫、巴拿马网纹虫、根状拟铃虫、节累枝虫、黏游仆虫、中华拟铃虫、树状聚缩虫;桡足类3种,分别为哲水蚤、桡足类六肢幼体、挪威小星猛水蚤;轮虫类2种,分别为蹄形腔轮虫、褶皱臂尾轮虫;其他浮游幼虫3种,分别为筒螅辐射幼虫、藤壶六肢幼体、幼蟹。
养殖前期原生动物、桡足类和轮虫类共为优势种,主要种类为诺氏麻铃虫、类铃虫、巴拿马网纹虫、哲水蚤、桡足类六肢幼体、褶皱臂尾轮虫;养殖中期原生动物和轮虫共为优势种,主要种类为巴拿马网纹虫、褶皱臂尾轮虫;养殖后期仅以原生动物为优势种,主要种类为巴拿马网纹虫、节累枝虫、黏游仆虫。
2.3.2 养殖全过程日本囊对虾精养池浮游动物种类数量变化。从图8可以看出,4个虾池浮游动物种类数量变化趋势均是养殖初期种类相对较少,随着养殖时间的延长,种类增多,养殖中期浮游动物种类数量达到高峰,养殖后期种类数量又有所下降。
2.3.3 日本囊对虾精养池浮游动物优势种演替动态。从图9可以看出,4个虾池的优势种的种类在养殖前期略有不同。由于各虾池底质状况不同,水质调控方法不同等原因导致放苗前水中的浮游动物种类有所差异,随着养殖的继续,养殖1月龄1号虾池以原生动物巴拿马网纹虫为优势种,2号虾池以原生动物巴拿马网纹虫和褶皱臂尾轮虫共为优势种,3号以哲水蚤和藤壶六肢幼体共为优势种,4号虾池则以哲水蚤和桡足类六枝幼体共为优势种。
养殖2月龄,1号、2号、3号虾池均以原生动物为优势种,主要种类为巴拿马网纹虫,4号虾池以原生动物巴拿马网纹虫和褶皱臂尾轮虫共为优势种。
养殖中期(养殖3月龄),4个虾池均出现轮虫,1号、2号虾池以其为优势种,3号、4号虾池以轮虫和原生动物共为优势种。
养殖后期(养殖4、5月龄),4个虾池均以原生动物为优势种,主要为节累枝虫和黏游仆虫等有害浮游生物。
2.3.4 日本囊对虾精养池浮游动物多样性。由表4可知,1号虾池多样性指数为0.57,优势集中性指数为0.70;2号虾池多样性指数为0.25,优势集中性指数为0.86;3号虾池多样性指数为0.65,优势集中性指数为0.62;4号虾池多样性指数为0.72,优势集中性指数为0.59。平均多样性指数最高值出现在4号虾池,2号虾池最低;各虾池优势集中性指数大小顺序为2号虾池>1号虾池>3号虾池>4号虾池。
总体而言,其多样性指数均较低,只有0.25~0.72,比自然海区低[10]。这主要与对虾养殖池在蓄水时的过滤(滤去大量浮游动物及幼体)和在整个养殖周期基本处于封闭状态有关。
2.4 日本囊对虾精养池浮游植物与浮游动物的相关关系
2.4.1 数量变化关系。从图10可以看出,浮游动物与浮游植物的栖息密度变化趋势较为一致,均表现为1月龄时浮游植物密度下降,浮游动物略有上升,而后二者的密度呈上升趋势,在3月龄达到峰值,此后随着养殖的进行二者的密度下降。4号虾池2~3月龄浮游植物密度出现的反常现象,与养殖2月龄在该池试用循环水机有关,循环水机中的滚筒过滤装置将部分浮游生物过滤出来,造成了虾池中浮游植物密度的减低,由于虾类、浮游动物的摄食作用,3月龄虾池中浮游植物的密度仍未恢复到正常状态;4月龄,随着浮游动物密度的慢慢降低,浮游植物的密度慢慢回升进入正常轨道。
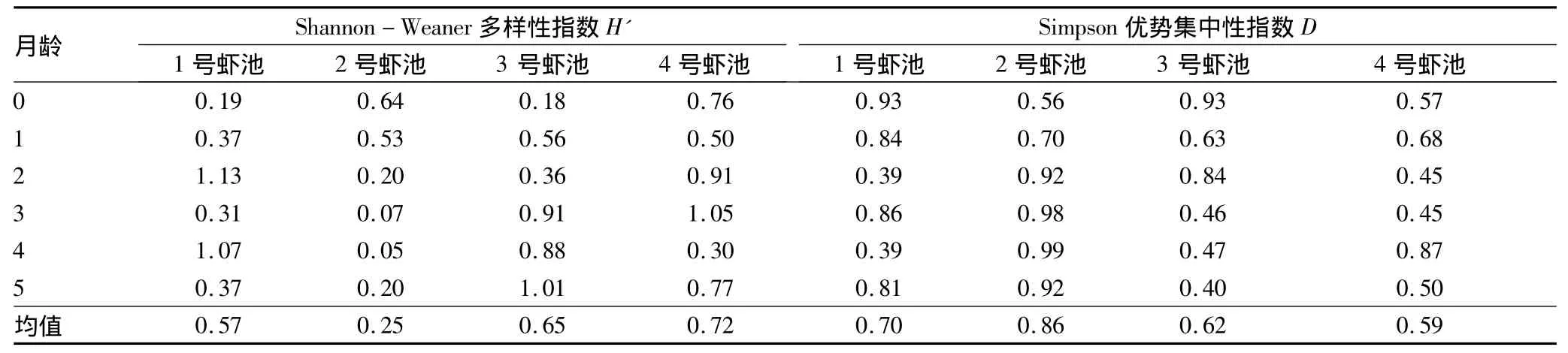
表4 日本囊对虾精养池浮游动物的群落多样性
2.4.2 数量的线性回归关系。将浮游动物个体数的平均值分别与浮游植物总细胞数的平均值进行线性回归分析。从图11可以看出,1号、2号、3号虾池浮游动物与浮游植物的关系在数量方面呈现较为密切的线性关系,二者之间的线性系数R2为0.718 1,其中线性关系最高为2号虾池,其次为1号虾池,最低为3号虾池。4号虾池在去除了2月龄与3月龄数据后,虾池浮游动物与浮游植物的关系在数量方面亦呈现极为密切的线性关系,二者之间的线性系数R2为0.992 5。
3 讨论
3.1 日本囊对虾精养池浮游生物种类组成及多样性特征
经过1个养殖周期日本囊对虾高位精养池的浮游生物监测发现,养殖前期,绿藻、硅藻多为优势种,原生动物、桡足类和轮虫类共为优势种;养殖中期,绿藻、硅藻多为优势种,原生动物和轮虫共为优势种;养殖后期,蓝藻、硅藻和绿藻多为优势种,原生动物为优势种;整个养殖周期中,浮游植物与浮游动物的多样性指数均较低,仅分别为1.19~1.30和0.25~0.72。
张汉华等[11]认为整个养虾周期养殖池塘浮游生物多样性指数为1~2,较自然海区的种类多样性指数(2~2.87)低。李永祺[12]等研究发现虾池内浮游植物群落的种类数相对较少,多样性指数相对较低,优势种亦与海域不同。黄翔鹄等[13]报道高位池浮游植物种类相对较少,优势种突出、单一,养殖中期、后期以绿藻、蓝藻为主。这些结论均与该试验监测结果相似。这与高位池整个养殖周期基本处于封闭状态,换水量少,使用的养殖用水均为经过砂滤等处理的过滤水,一开始便滤去了大量的浮游生物及幼体,加之养殖过程中,人为的饵料投喂、各种药物、营养盐的施用,水体营养化程度较正常海区增加的特殊环境有关[14-15]。这种水环境必然导致养殖前、中、后期浮游生物种类结构的差异。随着养殖的进行,水体富营养化程度逐渐增大,浮游生物的种类结构也逐渐向耐污型种类转变。
3.2 营养盐与浮游植物的关系 水体中的营养元素是浮游植物生长所必需的,因此营养盐浓度的高低与浮游植物的数量有着密切的关系。营养盐比例的变化尤其是氮磷比的变化,在决定浮游植物群落的组成上起着重要作用,它主要通过影响浮游植物之间的竞争和演替而对浮游植物群落产生影响[16]。研究表明,浮游植物的生长既与营养盐之间的比例有关,又与海水中营养盐的浓度有关[17]。也有学者认为氮、磷浓度越高,N/P距Redfield比值越远,浮游植物生物多样性指数越低[18],这与该试验结论是相符的,该试验中氮、磷浓度远高于海区,但N/P比仅为0.516~4.713,均小于16∶1,整个养殖周期的浮游植物生物多样性指数亦低于海区,仅为1.19 ~1.30。
对同一水体而言,其营养盐的峰值与浮游植物的繁殖峰期一般存在时间差,两者浓度或含量变化是动态的,具有一定的变化规律[19]。然而,虾塘是一个半封闭的人工生态系统,人为的、生物的、非生物的各种因素都会影响到虾池营养盐与浮游植物量的关系,二者之间的定量关系在虾塘是很难确定的。该试验监测了4个虾池各个养殖时期的N、P数据及其浮游植物数量变化,但分析显示4个虾池氮磷比与浮游植物总量无显著相关性。由此可见,只有通过连续的调查与监测,分析不同营养成分及其比例对浮游植物种类组成的影响,才能掌握营养盐和浮游植物质与量上的相关性[19]。
3.3 虾池浮游植物与浮游动物的关系 虾池浮游动物与浮游植物的数量存在显著相关性,二者的栖息密度变化趋势较为一致,均为1月龄浮游植物密度下降,浮游动物略有上升,而后二者的密度呈上升趋势,在3月龄达到峰值,此后随着养殖的进行二者的密度下降。
Harvery[20]摄食假说认为浮游植物与浮游动物二者在空间分布上呈相反关系,这解释了对虾养殖池前期浮游植物密度下降,浮游动物略有上升,可以认为前期浮游植物生物量较低是浮游动物摄食的结果。陈济丁[21]、韩希福[22]、赵文[23]等亦认为浮游动物是生物操纵的关键因子之一,主要依靠浮游植物的摄食来完成其能量的转换。
在虾塘生态系统中,浮游植物利用无机营养元素,通过光合作用合成有机物,浮游动物滤食浮游植物以及一定量的有机碎屑,早期对虾摄食一定量的浮游动物,这就形成了虾池中较为完整的食物链。然而,随着幼虾个体的成长,要求摄食的食物颗粒越来越大,其摄食浮游动物的机会越来越小,加上因为养殖过程中饲料投喂量增加而形成的残饵与有机碎屑量,均为浮游植物、浮游动物提供了营养盐与优质饵料,从而一定的时期内浮游生物的数量不断增加,在3月龄浮游生物数量达到了顶峰。养殖后期,由于养殖水体富营养化严重,水体中氨氮、亚硝酸氮等有害气体累积,可释放有毒有害物质的浮游植物大量滋生,并成为优势种,均制约了浮游动物的生长。
3.4 浮游生物对对虾养殖的影响 对虾养殖池是一个封闭、半封闭的生态系统,由对虾代谢物、残饵等外源性有机质分解形成的营养盐可被高密度的浮游植物利用,形成营养盐→浮游植物→轮虫→桡足类→幼虾的营养链,使虾池的生物、非生物环境达到相对平衡[24]。这种平衡环境下,对虾的自身免疫力较强,抵抗病害的能力也较强。
对虾的养殖实践证实微藻藻相的变动会影响水环境的稳定和对虾健康的生长,进而影响养殖效益。一般认为,在对虾养殖生产中通常以绿藻类和硅藻类为优势种的池塘为好,其水质稳定,病害少,对虾生长较好,其中裸藻类也能有效构建池塘良好水色,但其藻相较不稳定,易受环境影响[25]。以蓝藻、甲藻为优势的水体中,对虾一般生长缓慢且易引发病害。李卓佳等[26]认为绿藻类的许多微藻具有耐污、耐盐等宽生态位的性质,较易在养殖中期、后期逐渐富营养化的水体环境中形成相对稳定的藻相,对保持池水的“活、爽”作用功不可没,并且微藻种类数丰富,对于微藻多样性的增强也有益处。
但是,养殖水体中生物多样性较低,微藻种类单一,优势度过高,亦不利于养殖水体生态系统的稳定,不利于加强对虾抵抗胁迫因子的力度。若微藻种类过于单一,即使是绿藻类,当数量达到一定的阈值,也可形成水华,严重破坏水环境生态系统,导致对虾应激死亡。郭皓[27]和查广才等[28]研究发现过低的多样性指数容易导致对虾不同程度地出现病害和死亡。因此,合理的微藻种类、密度及物种多样性对调控虾池水质环境,提高对虾产量至关重要。
[1]张汉华,李卓佳,郭志勋,等.有益微生物对海水养殖池游生物生态特征的影响研究[J].南方水产,2005,1(2):7-14.
[2]吴斌,廖思明.广西北海凡纳滨对虾养殖池塘中微型藻类组成调查[J].广西科学,2008,15(4):452-455.
[3]杨秀兰,王爱敏,薄学锋,等.浮游生物在盐碱地封闭式对虾养殖中的生态作用[J].齐鲁渔业,2002,19(10):5-8.
[4]查广才,麦雄伟,周昌清,等.凡纳滨对虾低盐度养殖池浮游藻类群落研究[J].海洋水产研究,2006,27(1):1-7.
[5]王丽卿,王为东,臧维玲.河口区斑节对虾淡化养殖塘浮游生物状况[J].上海水产大学学报,2002,11(2):118-123.
[6]HUH J,LIRY,WEIH Z,et al.Freshwater algae of China[M].Shanghai:Shanghai Scientific & Technological Press,1980:478.
[7]SHEN Y F.Protozoology[M].Beijing:Science Press,1999:139-482.
[8]雷衍之,杨凤.化学实验[M].北京:中国农业出版社,2004:152-194.
[9]马克平,刘玉明.生物群落多样性的测度方法.I.α多样性的测度方法(下)[J].生物多样性,1994,2(4):231-239.
[10]黄长江,陈善文,何歆,等.2001-2002年粤东柘林湾浮游动物的生态学研究[J].海洋与湖沼,2003,34(2):117-124.
[11]张汉华,李卓佳,郭志勋,等.益生菌对海水虾池浮游生物的生态调控效果研究[J].海洋科学,2009,33(1):12-20.
[12]李永祺.海水养殖生态环境的保护与改善[M].济南:山东科学出版社,1999.
[13]黄翔鹄.对虾高位池优势浮游植物种群与成因研究[J].热带海洋学报,2002,21(4):36-44.
[14]孙耀,李锋,李健,等.虾塘水体中浮游植物群落特征与营养状况的关系[J].海洋水产研究,1998,19(2):47-50.
[15]姚泊,何建国,莫福,等.虾塘浮游生物种类调查[J].中山大学学报,2000,39(S1):224-228.
[16]RHEE GY.Phosphate uptake under nitrate limitation by Scenedesmus sp.and its ecological implications[J].Journal of phycology,1974,10:470-475.
[17]赵卫红,李金涛,杨登峰,等.现场培养实验研究营养盐对赤潮浮游植物生长的影响[C]//南中国海赤潮预防和管理国际研讨会.中国香港:南中国海会,2003:193-196.
[18]曲克明,陈碧鹃,袁有宪,等.氮磷营养盐影响海水浮游硅藻种群组成的初步研究[J].应用生态学报,2000,11(3):445-448.
[19]谢立民,林小涛,许忠能,等.虾池的理化因子及浮游植物群落的调查[J].生态科学,2003,22(1):34-37.
[20]郑重.浮游生物学概论[M].北京:科学出版社,1964.
[21]陈济丁,任久长,蔡晓明.利用大型浮游动物控制浮游植物过量生长的研究[J].北京大学学报(自然科学版),1995,31(3):373-381.
[22]韩希福,王荣.海洋浮游动物对浮游植物水华的摄食与调控作用[J].海洋科学,2001,25(10):31-33.
[23]赵文,李晓东,徐纪军.轮虫培育池不同粒级藻类对浮游植物生物量好生产量的贡献[J].水产学报,2004,28(2):167-174.
[24]杨秀兰,王爱敏,薄学锋,等.浮游生物在盐碱地封闭式对虾养殖中的生态作用[J].齐鲁渔业,2002,19(10):5-8.
[25]彭聪聪,李卓佳,曹煜成,等.虾池浮游微藻与养殖水环境调控的研究概况[J].南方水产,2010,6(5):74-80.
[26]曹煜成,李卓佳,杨莺莺,等.浮游微藻生态调控技术在对虾养殖应用中的研究进展[J].南方水产,2007,3(4):70-73.
[27]郭皓,于占国.虾池浮游植物群落特征及其与虾病的关系[J].海洋科学,1996(1):39-45.
[28]查广才,周昌清,黄建容,等.凡纳对虾淡化养殖虾池微浮游生物群落及多样性[J].生态学报,2004,24(8):1752-1759.
猜你喜欢
当代水产(2021年7期)2021-11-04
科学养鱼(2020年10期)2020-11-23
知识窗(2019年4期)2019-04-26
水利技术监督(2017年6期)2017-12-19
广东海洋大学学报(2015年3期)2015-12-22
应用海洋学学报(2015年3期)2015-11-22
现代盐化工(2015年3期)2015-02-24
湖泊科学(2014年2期)2014-02-27
云南农业科技(2011年5期)2011-12-10
