
黑龙江省西部草地蝗虫群落时间动态的研究
2015-03-17 10:15王皙玮张李香范锦胜王世喜周艳丽陈中华王贵强
黑龙江大学工程学报 2015年1期
王皙玮,韩 笑,张李香,范锦胜,孙 元,王世喜,周艳丽,陈中华,张 坤,王贵强,*
(1.黑龙江大学 农作物研究院,哈尔滨 150080;2.黑龙江大学 农业资源与环境学院,哈尔滨 150080;3.大庆市农业技术推广中心,黑龙江 大庆 150080)
黑龙江省西部草地蝗虫群落时间动态的研究
王皙玮1,韩 笑2,张李香2,范锦胜2,孙 元2,王世喜3,周艳丽2,陈中华2,张 坤2,王贵强2,*
(1.黑龙江大学 农作物研究院,哈尔滨 150080;2.黑龙江大学 农业资源与环境学院,哈尔滨 150080;3.大庆市农业技术推广中心,黑龙江 大庆 150080)
2011~2012年黑龙江省西部草地蝗虫群落时间动态的研究结果表明: 西部草地蝗虫6月初出现的早发型种类有中华稻蝗、毛足棒角蝗、宽须蚁蝗等,6月中下旬出现的晚发型的种类有大垫尖翅蝗、笨蝗、亚洲小车蝗、红翅皱膝蝗、轮纹异痂蝗、白边雏蝗等。蝗虫发生的种类及数量随着时间变化渐增或减少。优势种的中华稻蝗在群落中所起的主导作用明显,并通常选择植被覆盖率较低、地表裸露向阳、土壤pH值为碱性且有机质和含盐量较低的栖息地存活,其发生数量呈现上升趋势,应防止其在农牧交错地带暴发成灾。
草地蝗虫;群落结构;时间动态;生态系统
0 引 言
黑龙江省是我国拥有草原的10个省份之一,草地面积为433×104hm2,多分布于西部松嫩平原地区的大庆、杜蒙、肇源及肇州等市县,也是本省的主要畜牧产区。该产区的草地由于其特殊的自然地理及气候条件,蝗虫发生的种类较多[1]。近十几年来,由于全球性气候变暖对西部草地生物多样性的影响,以及持续性的干旱,温度、湿度以及营养条件的改变,草地植被退化严重,荒漠化趋势严峻,草地蝗虫发生危害加重,对当地的生态环境质量和经济发展已造成深刻影响[2-6]。
自2000年以来,亚洲飞蝗在松嫩平原的黑龙江与吉林两省曾多次大规模爆发。例如:2001年在吉林省长春市农安县巴吉垒镇波罗湖地区亚洲飞蝗的大面积发生危害,受害的苇塘及草地面积为12 000 hm2,农田面积约6 700 hm2,2009年该地区亚洲飞蝗再次大爆发,受灾面积为3 000 hm2。在2003年至2011年吉林省大安县仅间隔几年就接连大规模发生亚洲飞蝗,受灾面积为1 000 hm2,且大爆发时其蝗蝻虫口密度为1 000~10 000头/m2,成虫将植被的叶片、叶鞘全部吃尽仅留茎秆,给农牧业生产造成重大的经济损失[7]。
黑龙江省每年均有一定面积的草地遭受到虫害的威胁,其中草地蝗虫就是危害较重的害虫之一。2009年是黑龙江省70 a以来的首次记录亚洲飞蝗(Locusta migratoria migratoria )在本省肇州县与肇源县交界的大白水苇塘,以及龙江县哈拉海湿地大规模的爆发,其发生面积分别为6 700 hm2和14 400 hm2。近十年来,黑龙江省西部地区畜牧业发展迅速、生态环境建设等问题日益受到人们的关注。虽然国家及地方政府对草地蝗虫防治也提供了必要的人力、物力及财力,然而每年草地蝗虫的防治面积只占发生面积的10%左右,尚难以有效遏制虫害加重趋势[8-12]。加强黑龙江省西部草地的生态环境研究,建立生态友好、资源可持续利用的蝗虫治理体系已成当务之急。
国外在蝗虫群落结构变迁与环境因子之间的关系已有较多的研究报道。不同的植物群落类型为草地蝗虫提供了各异的生存和繁殖环境,因而也造就了物种的多样性。Solomon等[13]研究南非等地的坡度和植被覆盖度等地形异质性与蝗虫群落多样性之间的关系结果表明,尽管小山区的蝗虫物种丰富度和密度均较大山区的相对要大,地形异质性对蝗虫物种丰富度的影响较小。然而也有研究认为,地形异质性对蝗虫群落结构的变化存在重要的影响[14-18],如人类的火烧和刈割等活动会严重影响蝗虫的物种数及丰富度[19]。
有研究表明美国迈阿密地区植物群落的组成与昆虫群落空间分布之间的关系,呈现昆虫栖地的牧草覆盖度对昆虫的物种丰富度会产生重要影响[20]。Gesarr等的报道也证实:人类的畜牧业生产活动,不仅改变了草地的土壤紧实度,还能影响植物群落的结构和多样性,使生态系统的结构和功能发生改变,蝗虫的物种丰富度与牲畜对草地的啃食与践踏关联显著[21]。
植物为蝗虫生存提供了必需的食物、适宜的微气候等环境条件,而蝗虫对食物的选择性也能够影响植物群落结构的改变[22]。许多关于蝗虫群落结构变化与植物群落之间关系的研究证实两者群落多样性之间存在密切关系[23-30],优势植物的生物量及物种丰富度对蝗虫群落结构产生重要影响[31],但持反对意见的则认为,植物群落的多样性与蝗虫物种丰富度之间的关系并不显著[32-33]。
近年来,国内有关蝗虫发生的时间动态研究虽有报道[7,34],但关于黑龙江省西部草原蝗虫群落结构组成和发生的时间动态研究却鲜有报道。草地蝗虫在草地生态系统的物质循环和能量流动中扮演重要作用,它们不仅是消费者,而且又是次级生产者,在食物链与食物网中是不可或缺的重要链节。不同生态环境中蝗虫种类组成和发生的时间动态具有较大差异性,因此,研究蝗虫种类组成和群落的时间动态规律性对于揭示草地生态系统的物质循环和能量流动,监控蝗虫种群密度变化动态,为蝗虫治理及蝗区生态环境的改造既可提供理论支撑,还可达到有效控制蝗虫危害与草地资源可持续利用的目的。
本文选取黑龙江省大庆市让胡路区喇嘛甸镇兴华村、杜尔伯特蒙古族自治县一心乡前锋村、肇源县头台镇团结村及肇州县肇州镇中华村的草甸草原4个不同植被的草地样区,对黑龙江省西部草地蝗虫及优势蝗种发生的时间动态及影响因子间相互关系进行研究,旨在为蝗虫的预测预报和科学防治提供依据。
1 材料与方法
1.1 昆虫标本的采集与鉴定
选取黑龙江省西部地区的大庆市让胡路区喇嘛甸镇兴华村、杜尔伯特蒙古族自治县一心乡前锋村、肇源县头台镇团结村、肇州县肇州镇中华村等4处生境类型不同的草地进行蝗虫扫网采集抽样调查。取样时间为2011年和2012年的6~9月,期间每隔10~15 d调查一次。每次调查的时间均固定在上午9:30~12:30,若遇雷雨或大风天气而无法工作则在随后之日进行取样。调查时,在样区内采用五点式取样法,每点用标准捕虫网贴近草面随机扫取100网,将采集到的昆虫放入毒瓶处死,再装入写有标签的塑料袋内带回实验室进行种类鉴定,统计个体数量及数据整理。
1.2 土壤样品的采集和自然特征的统计
采集土样时,各样区均采用棋盘法。首先,在每个试验样区选取5~10个1 m×1 m大小的样点。然后,每个样点用军锹沿草地表层垂直向下挖深20 cm做一剖面,再用土铲自上而下垂直向下切取约1 cm厚的土壤,将采集的土样放入注明标签的塑料袋中,带回实验室中待测。对土样的理化性质在进行测定之前,先将同一样区各样点采集到的土壤放在一起混合均匀,去除土样内的侵入体,最后将土壤密封干燥待测定。
在对各样区的土壤样品进行采集的同时,还要调查各样区的植被种类及其覆盖度情况以及当地2011年及2012年的气象资料,分析温度、降水量、相对湿度等环境因子与黑龙江省西部草地蝗虫发生之间的关系。
1.3 土壤理化性质的检测方法
采用电位法测定土壤的pH值。将pH玻璃电极和甘汞电极插入土壤悬液中,测定电动势值后换算成pH值的方法。测定时,水土比例对pH值有较大影响,尤其对石灰性土壤稀释效应的影响更为严重,因此本研究测定时采用1∶1的小水土比例。
采用重铬酸钾容量法测定土壤的有机质含量,将定量5 mL的重铬酸钾—硫酸溶液在电加热条件下,使土壤中的有机质氧化。然后将剩余的重铬酸钾用硫酸亚铁标准溶液滴定,以二氧化硅为添加剂作为空白标定。根据氧化前后氧化剂质量的差值,计算出有机碳量,再乘以系数1.724,即为土壤有机质的含量。
以残渣烘干法测定土壤中盐总量时,首先吸取一定量的土壤浸出液放在瓷蒸发皿中,水浴蒸干后用过氧化氢氧化有机质,在105~110 ℃烘箱中烘干,称重,即得烘干残渣质量。
2 结果与讨论
2.1 蝗虫发生期类型的划分
自2011和2012年的调查数据结果显示:黑龙江省西部草地大多数种类的蝗虫发生时期集中出现于7~8月。若以蝗蝻出现时间的早晚划分发生期类型,可分为早发型与晚发型两种类型(表1)。早发型的蝗虫一般于6月初即可见到蝗蝻,包括的种类有中华稻蝗(Oxyachinensis)、毛足棒角蝗(Dasyhippusbarbipes)、宽翅曲背蝗(Paracrypteramicropterameridionalis)、宽须蚁蝗(Myrmeleotettixpalpalis)等。而晚发型的一般约在6月下旬始见蝗蝻,7月中下旬至8月上旬发生危害,这些种类包括大垫尖翅蝗(Epacromiuscoerulipes)、笨蝗(Haplotropisbrunneriana)、亚洲小车蝗(Oedaleusasiaticus)、红翅皱膝蝗(Angaracrisrhodopa)、轮纹异痂蝗(Bryodemellatuberculatumdilutum)、白边雏蝗(Chorthippusalbomarginatus)以及黄胫小车蝗(Oedaleusinfernalis)等。不同蝗虫种类其发生时期上的差异,除了种类遗传特性差异之外还有环境因素如温度、光照、植被等多种生态因子综合作用的结果,也体现出不同的蝗虫种类通过适应发生时期的早晚来与其生态环境形成特有的共存性。
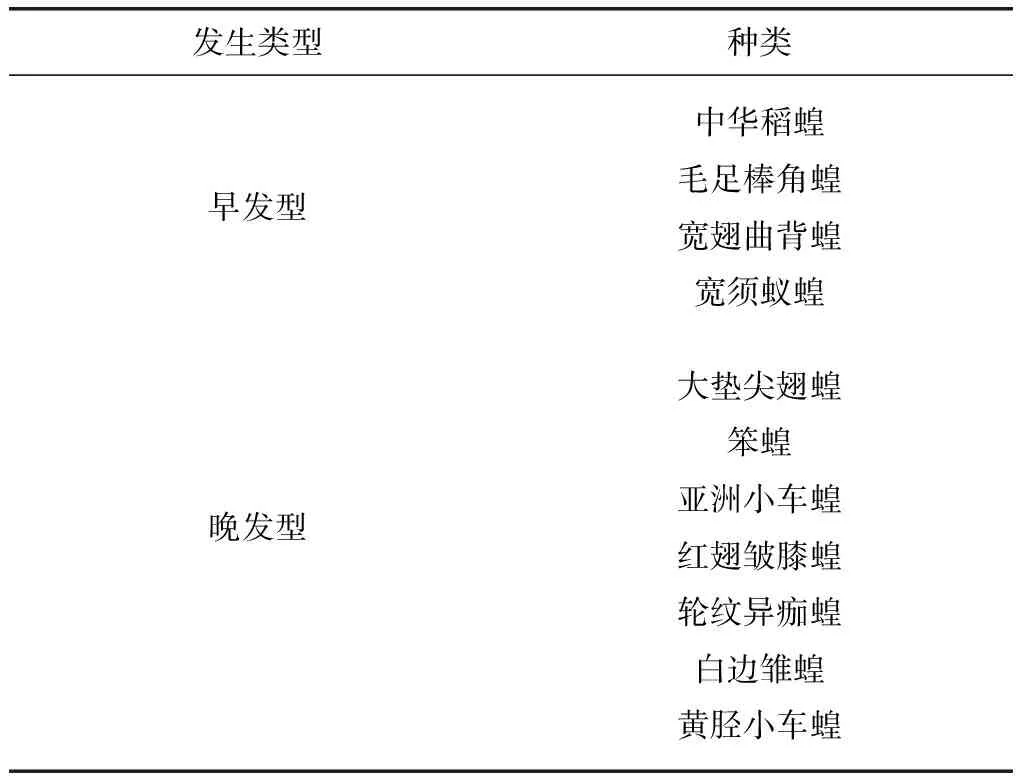
表1 黑龙江省西部草地蝗虫群落组成的时间动态
2.2 蝗虫科、属、种组成发生的时间动态
调查期间黑龙江省西部草地蝗虫7科14属16种的群落结构组成发生的时间动态,既受环境条件中季节性周期变化的制约,又与不同蝗虫种类的遗传特性、物候周期及其取食植物的群落密切相关。
该省西部地区物候期特点,一般是5月上旬植物返青发芽,7月中旬开花结实,9月末植物地上部分枯黄。与物候变化相适应的是,栖息于植物群落中的蝗虫群落结构也随之发生系列变化。从2011和2012年4样区调查蝗虫科、属、种组成的时间动态结果显示,6月初只有中华稻蝗、毛足棒角蝗、宽须蚁蝗等少数几种早发型蝗虫出现,但随着时间的推移,蝗虫的科、属、种类数以及种群密度逐渐增加,至7月蝗虫发生的种类及数量达到一年中的高峰(图1~图4)。自8月中下旬蝗虫发生的种类数逐渐减少,至9月初只有笨蝗、大垫尖翅蝗等少数蝗种存在。草地蝗虫的这种发生规律与黑龙江省的季节变化相吻合,6月初黑龙江省西部地区的平均气温虽达20 ℃但昼夜温差较大,除早发型蝗虫外,有效积温还不能满足多数蝗虫种类卵发育的需要,延至6月下旬卵才孵化开始出现蝗蝻。随着气温逐渐升高,草地植被生长日见繁盛并在7~8月达到生长的高峰期,此时蝗虫的科、属、种数达到一年中的最高值。当9月气温转凉,植被结实枯黄,蝗虫种类数随之减少,草地蝗虫一年的活动时间动态表明其对生态环境的高度适应性。
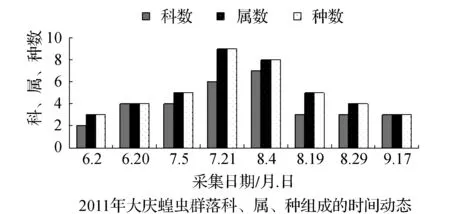
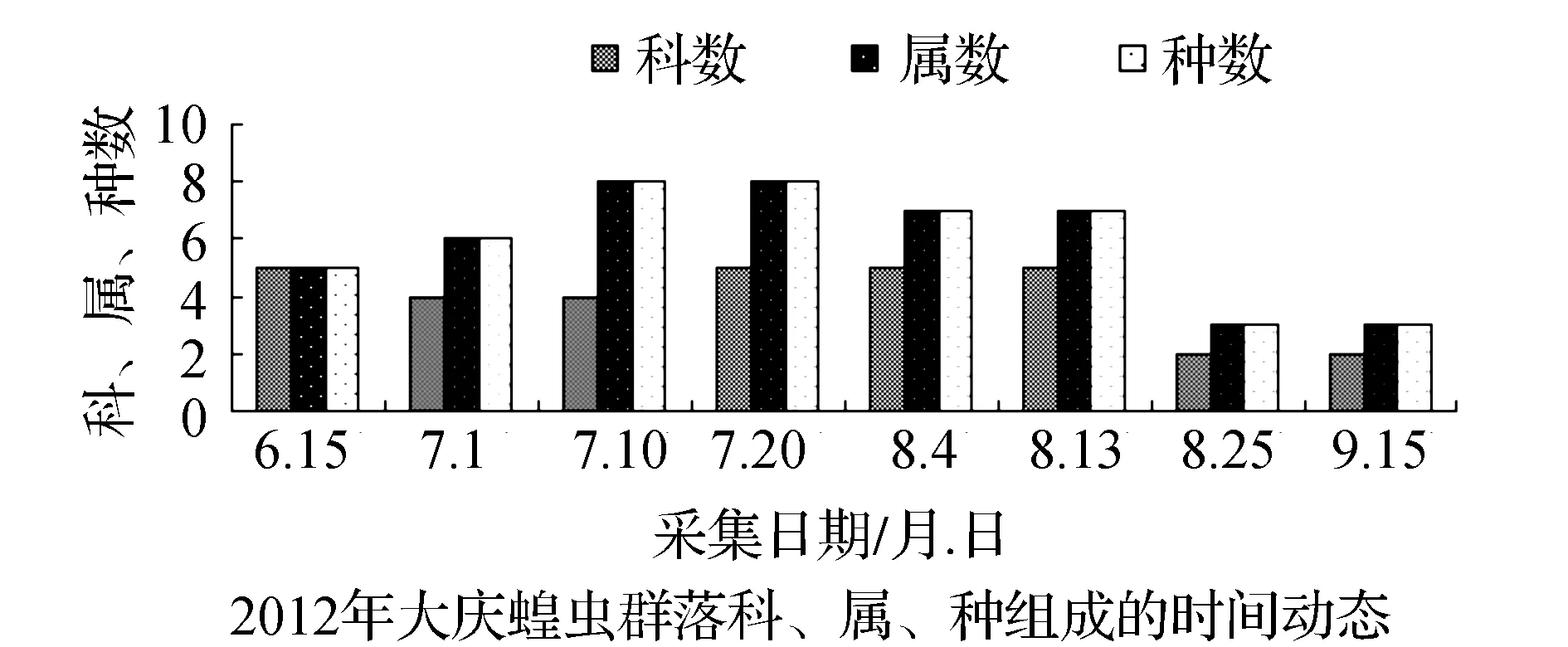
图1 2011~2012年大庆蝗虫群落科、属、种组成的时间动态Fig.1 Time dynamics of grasshopper community make up of region, genus and species in Daqing in 2011 and 2012
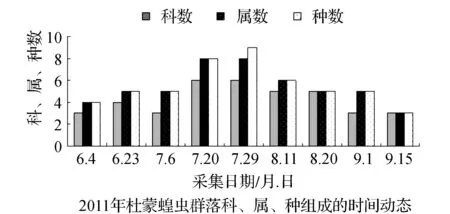
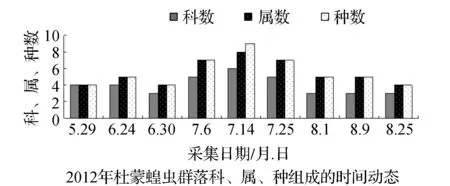
图2 2011~2012年杜蒙蝗虫群落科、属、种组成的时间动态Fig.2 Time dynamics of grasshopper community make up of region, genus and species in Dumeng in 2011 and 2012
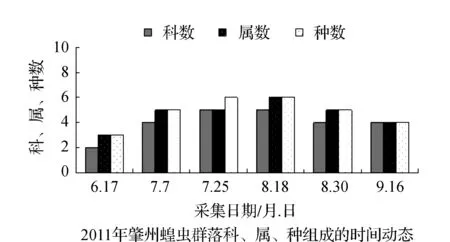
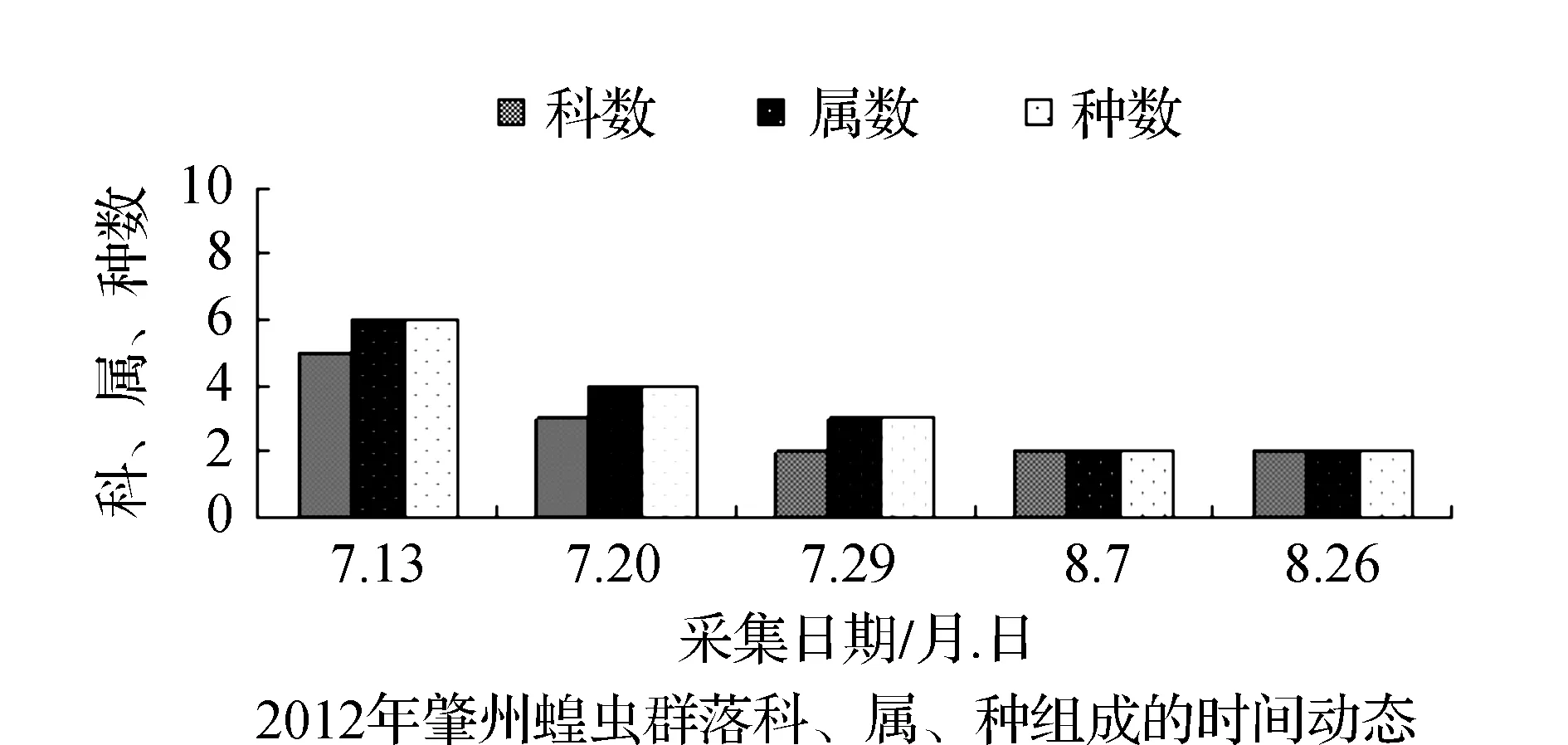
图3 2011~2012年肇州蝗虫群落科、属、种组成的时间动态Fig.3 Time dynamics of grasshopper community make up of region, genus and species in Zhaozhou in 2011 and 2012
2.3 西部草地蝗虫优势种群的时间动态
2011及2012年4样区蝗虫种群优势度时间动态调查结果显示,中华稻蝗、大垫尖翅蝗、笨蝗、中华剑角蝗,在群落结构中起优势种的主导作用,其发生时间的早晚和数量高峰期的先后差异明显(表2~表5)。其中,中华稻蝗在一年中的6~8月
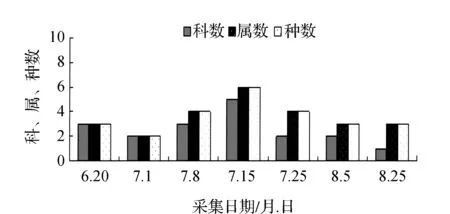
图4 2012年肇源蝗虫群落科、属、种组成的时间动态Fig.4 Time dynamics of grasshopper community make up of region, genus and species in Zhaoyuan in 2012
均占据主导作用。6月只有中华稻蝗发生数量最多,其他种类的蝗虫数量发生的很少,7月中华稻蝗的数量达到高峰期。早发型蝗虫种类的遗传特性决定了其卵的发育起点温度要求低,所需积累的热量值较小,满足阶段发育有效积温要求的历期较短,中华稻蝗发生的时间就早,而此时晚发型蝗虫种类还未出现,因此它在种群数量上占据了绝对优势。8月中华稻蝗数量开始呈现下降趋势,而大垫尖翅蝗和中华剑角蝗等数量达到了高峰,笨蝗的数量也逐渐呈现上升的趋势,直至9月笨蝗的数量到达最高,而中华稻蝗的数量明显降到最低(图5~图8)。由此可见,优势蝗种的种群发生时间、盛发期数量不尽相同,各蝗虫种类的生活习性、生理特点、活动规律在种属间存在差异,呈现出各蝗种与生态环境相互适应协同演化的关系。
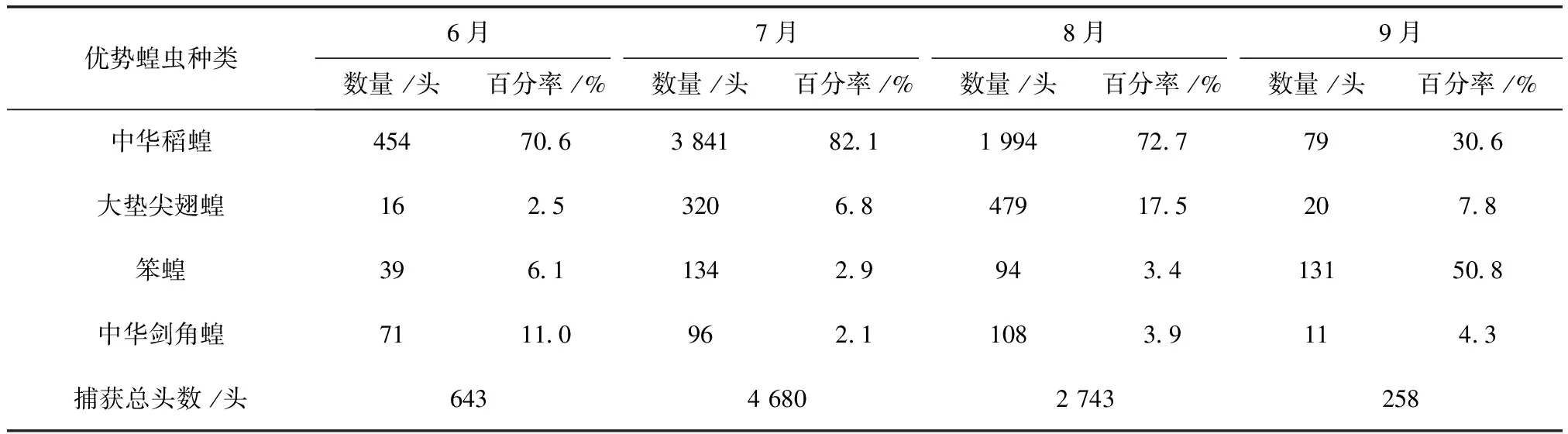
表2 2011~2012年大庆各月份优势蝗虫数量及百分率
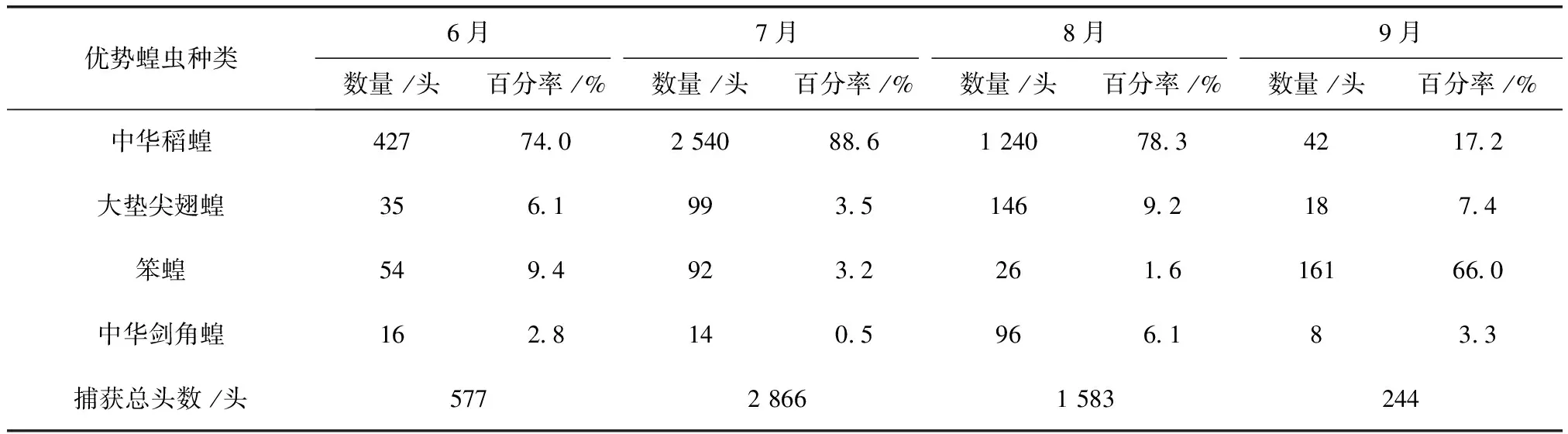
表3 2011~2012年杜蒙各月份优势蝗虫数量及百分率
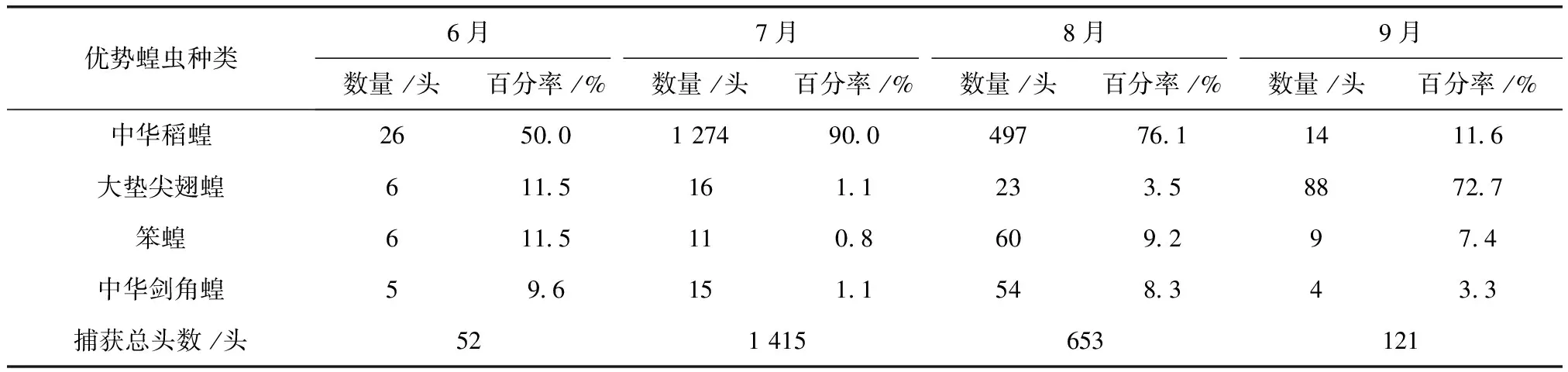
表4 2011~2012年肇州各月份优势蝗虫数量及百分率
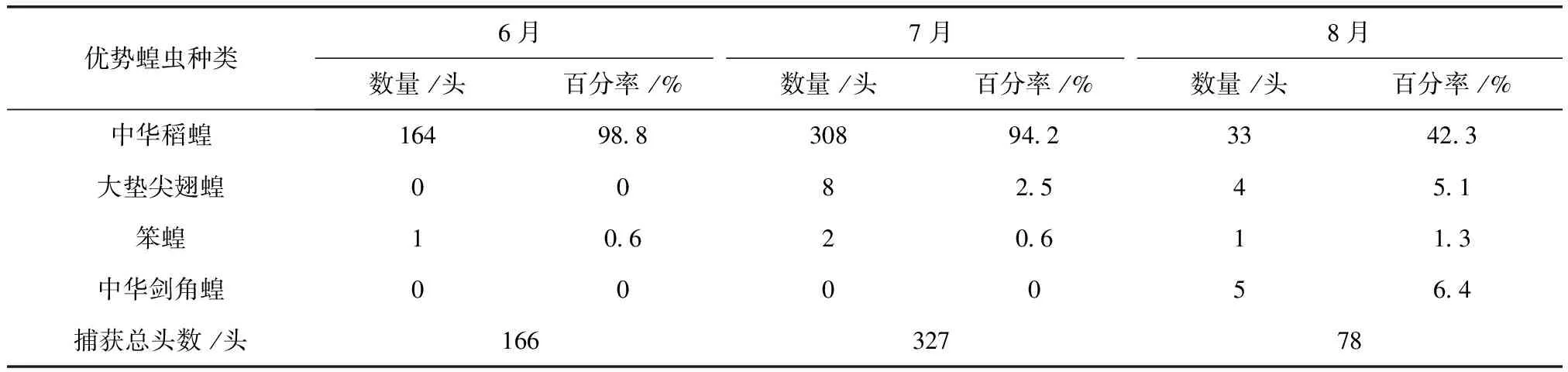
表5 2012年肇源各月份优势蝗虫数量及百分率
2.4 中华稻蝗的发生与环境因子之间的关系
有研究报道中华稻蝗在黑龙江省西部地区一年发生一代,以卵在土壤中越冬。次年春季随着气温的升高,卵于5月上中旬开始陆续孵化,6月上旬至7月中旬为蝗蝻期,7月下旬至8月上旬羽化为成虫,8月下旬交尾产卵于土壤中,10月中旬成虫死亡。本调查结果表明中华稻蝗在不同群落中起到明显的主导作用,因而对中华稻蝗的发生与环境因子之间关系有必要进一步研究。
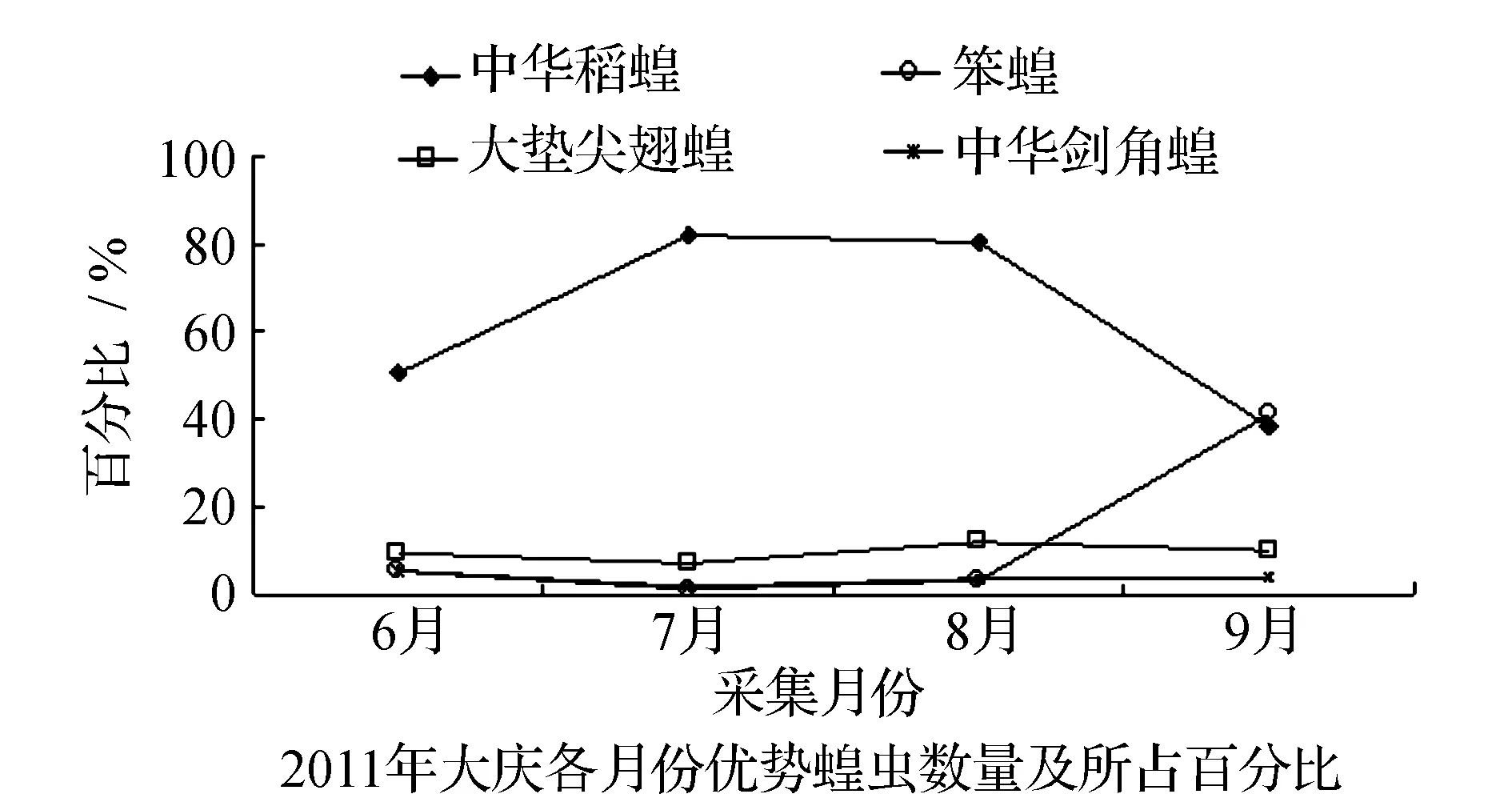
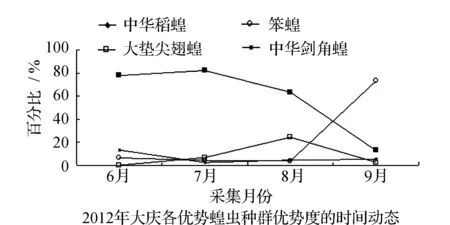
图5 2011~2012年大庆各优势蝗虫种群优势度的时间动态Fig.5 Temporal dynamics of dominant grasshopper populations in Daqing in 2011 and 2012
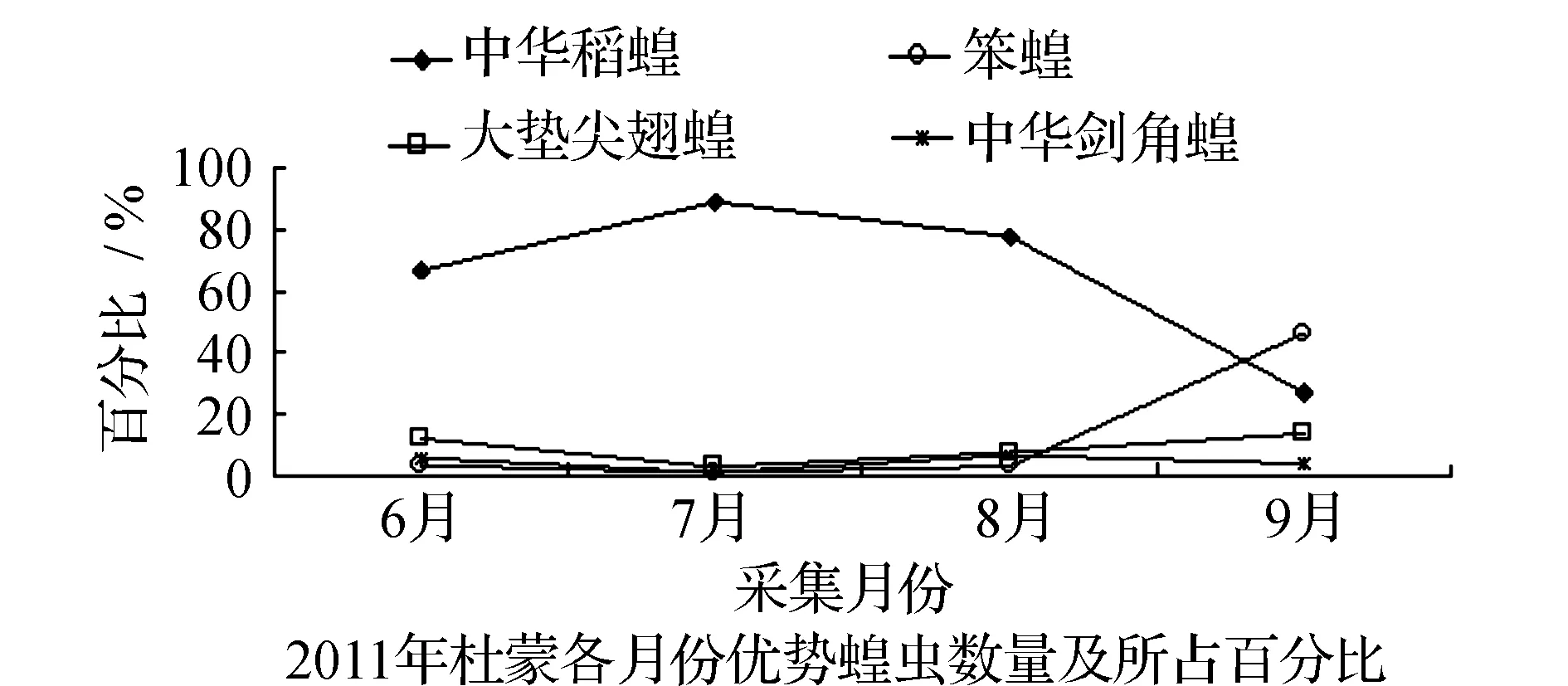
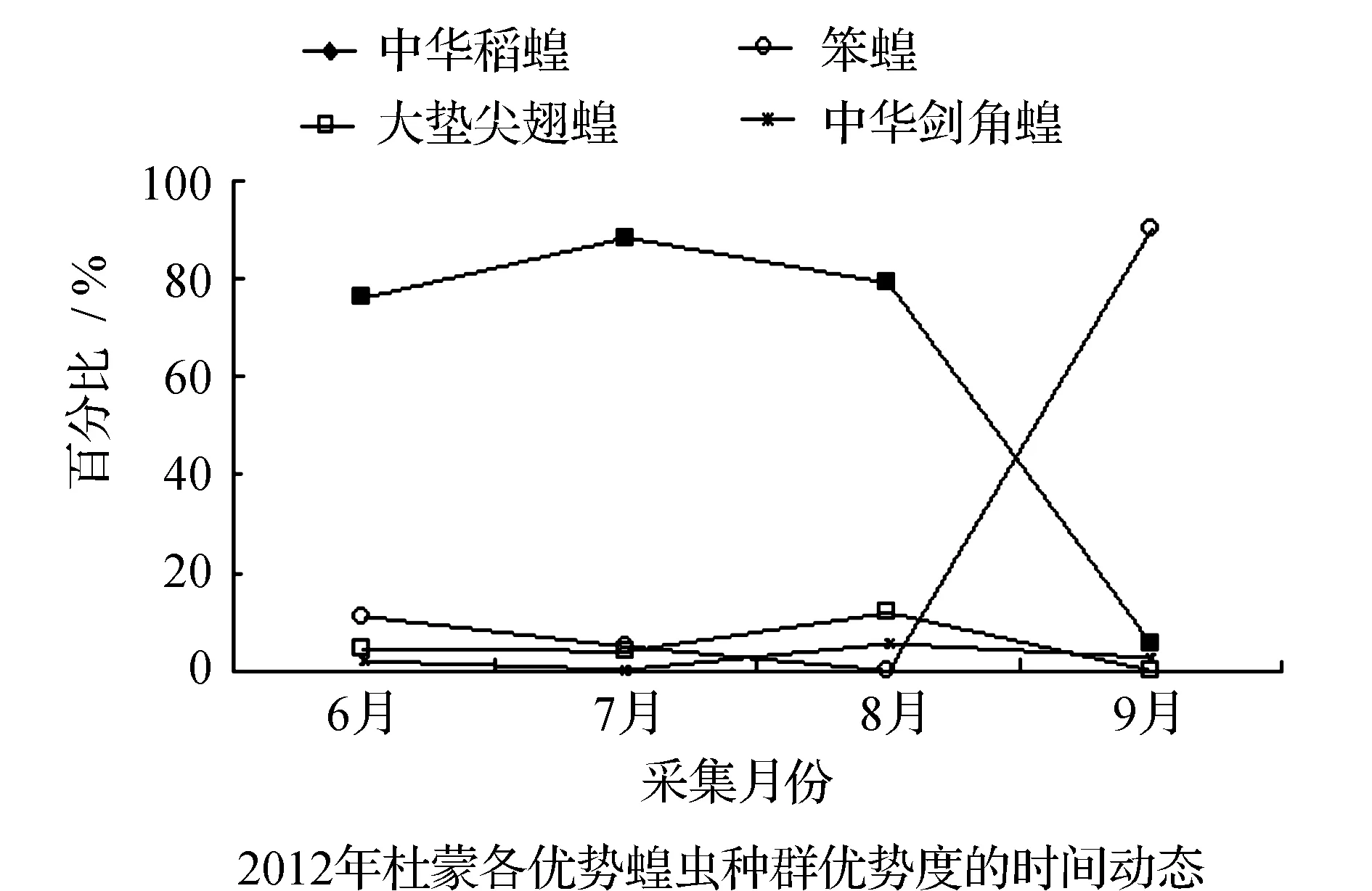
图6 2011-2012年杜蒙各优势蝗虫种群优势度的时间动态Fig.6 Temporal dynamics of dominant grasshopper populations in Dumeng in 2011 and 2012
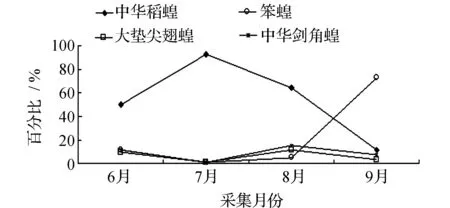
图7 2011年肇州各优势蝗虫种群优势度的时间动态Fig.7 Temporal dynamics of dominant grasshopper populations in Zhaozhou in 2011
2.4.1 中华稻蝗发生区土壤的理化性质
土壤是影响蝗虫发生的重要环境因素之一,其理化性质直接影响蝗虫产卵地的选择及卵在土壤中的发育和孵化。不同土壤类型其生长的植被种类差异也可间接影响蝗虫的空间分布。经对大庆、杜蒙、肇源、肇州4样区的土壤理化性质测定并计算各区的蝗虫发生数量与pH、有机质含量、总盐量之间的相关性结果表明:西部草地蝗虫发生区的土壤pH为7~10,土壤含盐量为0.9~1.5 g/kg,土壤有机质含量13~34 g/kg,变化较大。相关系数(r值)分别为0.593、-0.633、-0.784,其绝对值均大于0.5,表明蝗虫的发生量与各地区土壤的理化性质相关关系明显(表6)。样区间土壤理化性质的差异主要与土壤表层植被的生长情况有关。有研究报道,土壤含盐量对蝗虫的产卵具有明显影响。土壤盐分过高不利于蝗虫产卵,蝗虫产卵地的土壤含盐量一般小于1%,而土壤含盐量大于1%时蝗虫产卵量显著下降。尽管黑龙江西部草地蝗虫发生区的土壤条件适宜中华稻蝗产卵,但是对于活动区域内土壤的理化性质仍有一定的选择性。通常选择在植被覆盖率较低、地表裸露向阳、土壤的pH值为碱性、有机质和含盐量较低的区域更有利于其存活。调查结果中大庆样区的土壤pH 值偏碱性,有机质和含盐量相对较低,中华稻蝗的发生数量最大也是极好的例证。
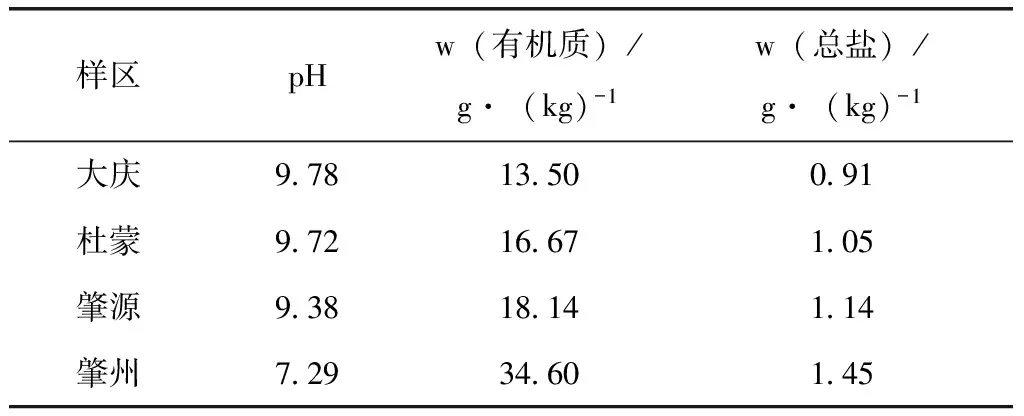
表6 2012年各样区的土壤理化性质
2.4.2 中华稻蝗发生区的气候特点
对2011~2012年黑龙江省西部草地蝗虫6~9月发生期的气温、相对湿度以及降水量的调查结果见表7。统计计算蝗虫发生数量与其对相应的平均气温、平均相对湿度以及降水量之间的相似性系数以及发生量与环境因子之间的关系结果表明:蝗虫的发生量与平均气温、平均相对湿度以及降水量间的相关系数分别为0.071、-0.21、0.029,r值未达显著标准,各变量之间的线性相关程度非常低,也证实了西部草地蝗虫发生期间6~9月的气温、相对湿度和降水量,对中华稻蝗蝗卵孵化与蝗蝻生长发育没有显著的影响。

表7 2011~2012年各样区6~9月的气象资料统计
3 结 语
1) 黑龙江省西部草地蝗虫种类每年大多于6~9月出现。6月初早发型种类有中华稻蝗、毛足棒角蝗、宽翅曲背蝗、宽须蚁蝗等;6月中下旬晚发型的种类有大垫尖翅蝗、笨蝗、亚洲小车蝗、红翅皱膝蝗、轮纹异痂蝗、白边雏蝗等。蝗虫群落科、属、种组成的时间动态,是随着季节转换、温度变化其科、属、种类数量以及个体数量逐渐增多或减少。
2) 黑龙江省西部草地蝗虫优势种中华稻蝗、大垫尖翅蝗、笨蝗、中华剑角蝗的发生和高峰期虽各不相同,但它们在群落中所起的主导作用非常明显,各优势种的生活习性、生理特点、活动规律之间的差异,亦是与生态环境相互适应,协同演进的结果。
3) 中华稻蝗的发生对于栖息地土壤的理化性质具有一定选择性,通常选在植被覆盖率较低、地表裸露向阳、土壤pH值为碱性且有机质和含盐量较低,有利于蝗卵存活和孵化的场所。
4) 黑龙江省西部草地蝗虫群落结构的演替中,中华稻蝗呈现上升趋势,应注意防止其在农牧交错地带形成新的蝗灾发生区,并持续加强对西部草地蝗虫群落结构多样性动态的深入探究。
[1]任炳忠,王皙玮,王贵强.东北草地的蝗虫 [M]. 哈尔滨: 黑龙江大学出版社,2012.
[2]倪绍祥.环青海湖地区草地蝗虫遥感监测与预测[M].上海:上海科学技术出版社,2002.
[3]吴瑞芬,霍治国,卢志光,等,蝗虫发生的气象环境成因研究概述[J].自然灾害学报,2005,14(3),66-73.
[4]赵 军,党国锋. 祁连山草地资源利用面临的问题及治理对策[J]. 草业科学,2003,20(7):1-4.
[5]刘 玲,郭安红.2004年内蒙古草地蝗虫大发生的气象生态条件分析[J].气象,2004,30(11):55-57.
[6]韩秀珍,马建文,马超飞,等.利用TM及地面样方与光谱数据综合分析东亚飞蝗灾害[J].自然灾害学报,2001,10(4):173-178.
[7]任炳忠,赵 卓. 松嫩平原蝗虫群落动态变化初步研究[J]. 吉林农业大学学报,2000,22:123-128.
[8]李宏昌,陈永林. 内蒙古典型草原亚带锡林河流域蝗虫区系的研究[J]. 草原生态系统研究,1988:26-40.
[9]童雪松,潜祖琪. 浙西南山区蝗虫的生态地理研究[J]. 昆虫知识,1987,2:4-10.
[10]杨大荣. 滇西北昆虫区系特点[J]. 动物学研究,1992,13(4):333-341.
[11]廉振民,梁 沛. 秦巴山区蝗虫群落多样性研究[J]. 生物多样性,1999,7(2): 119-122.
[12]钟玉林,郑 坚,郑哲民. 湖北大别山蝗虫区系研究[J]. 华中师大学报:自然科学版,2001,35(4):459-463.
[13]Solomon G, Michael J S. Topographic heterogeneity plays a crucial role for grasshopper diversity in a southern African megabiodiversity hotspot [J]. Biodiversity and Conservation, 2006, 15:231-244.
[14] Stabaev I V. Periodic changes in the ecological distribution of grasshoppers in the temperate and extreme continental Steppe regions, and their importance for the local ecosystems.In Hemming C.F. and Taylor T.H.C.(eds), Proceedings of the International Study Conferenceon Current and Future Problems of Acridology [J]. Londonpp, 1972, 207-213.
[15]Narisu, Lockwood,Schell . Rangeland grasshopper movement as a function of wind and topography: implications for pest management [J]. J.Orthoptera Res, 2000, 9:111-120.
[16]Wachter,O′Neill,Kemp. Grasshopper (Orthoptera:Acrididae) communities on an elevational gradient in southwestern Montana.J.Kansas Entomol [J]. Soc. 1998, 71:35-43.
[17]Sam ways M J. Land forms and winter habitat refugia in the conservation of montane grasshoppers in South Africa[J]. Conserv,Biol, 1990, 4: 375-382.
[18]Claridge, Singhrao. Diversity and altitudinal distribution of grasshoppers (Acridoidea) on a Mediterranean mountain[J].J. Biogeogrific, 1978, 5:239-250.
[19]Chambers, Samways. Grasshopper response to 40-year experimental buring and mowing regim, with recommendations for inverbrate conservation management[J]. Biodiversity and Conservation 1998,7: 985-1 012.
[20]Crist, P D,Summerville. Spatial variation in insect community and species responses to habitat loss and plant community composition[J]. Oecologia, 2006, 147: 510-521.
[21]Samways,Knut kreuzing. Vegetation, ungulate and grasshopper interactions inside vs.outside an African savanna game park[J]. Biodiversity and conservation, 2001,10:1 963-1 981.
[22]Joern. Resource utilization and community structure in assemblages of arid rangeland grasshoppers[J]. Transactions of American Entomological Society, 1979,105: 253-300.
[23]Sandra, Maria. Grasshopper (Orthoptera: Acridoidea) and plant community relationships in the Argentine pampas[J]. Journal of Biogeography, 2009, 29: 221-229.
[24]Stern V M. The integration of the mi cal and biological control of the spotted aphid [J]. Hildegard, 1995, 29 (2):81- 101.
[25]Hunter M D. Interaction within herbivore communities mediated by the host plant: The keystone herbivore concept[C]//Effects of Resource Distribution on Animal-Plant Interactions.Academic Press,1992:287-325.
[26]Tilman D. Resource Competition and Community Structure[M]. Princeton: Princeton University Press.1982.
[27]Bach,C E. Long-term effects of insect herbivory and sand accretion on plant succession on sand dunes [J]. Ecology,2001,82:1 401-1 416.
[28]Fagan,Bishop. Trophic interactions during primary succession: herbivores low aplant reinvasion at Mount St.Helens [J]. America Naturalist,2000,155:238-251.
[29]May R M. How many species Philosophical Transactions of the Royal Society of London[J]. Biological Sciences,1990,327:171-182.
[30]Carson W P, Root. Top-down effect of insect herbivores during early succession: Influence on biomass and plant dominance[J]. Oecology, 1999,121:260-272.
[31]Mark, Ritchie, Tilman. Interspecific competition among grasshoppers and their effect on plant abundance in experimental field environments[J]. Oecologia,1992, 89:524-532.
[32]Joern A. Grasshopper-communities respond to fire,bison grazing and weather in North American tall grass prairie:a long term study[J]. Oecologia, 2007,153:699-711.
[33]Symstad, Evan S, John H. An experimental test of the effect of plant functional group diversity on arthropod diversity[J]. Oikos, 2000,89:243-253.
[34]吴 亚,金翠霞. 草甸昆虫群落及其空间与时间结构[J]. 昆虫学报,1980,23(2):156-165.
Research on community dynamics of rangeland grasshopper in western Heilongjiang Province
WANG Xi-Wei1, HAN Xiao2, ZHANG Li-Xiang2, FAN Jin-Sheng2, SUN Yuan2, WANG Shi-Xi3, ZHOU Yan-Li2,CHEN Zhong-Hua2, ZHANG Kun2, WANG Gui-Qiang2,*
(1. College of Agricultural Resource and Environment, Heilongjiang University, Harbin 150080, China; 2. College of Biology, Heilongjiang University, Harbin 150080, China;3.Daqing Extension Center of Agricultural Technology, Daqing 163002,Heilongjiang,China)
Research on community dynamics of rangeland grasshopper was undertaken in the western Heilongjiang Province from 2011 to 2012, and the results showed that there were two appearing types of grasshoppers, one was the early June appearing species ofOxyachinensis,DasyhippusbarbipesandMyrmeleotettixpalpalis, and the other was the late June appearing species ofEpacromiuscoerulipes,Haplotropisbrunneriana,Oedaleusasiaticus,Angaracrisrhodopa,BryodemellatuberculatumdilutumandChorthippusalbomarginatus. The species and numbers of increasing or decreasing were over time. Dominant species of O. chinensis obviously played a leading role in the community, and often select lower vegetation cover, surface exposed Sunny, soil organic matter and a pH of alkaline and lower-salinity habitat to survive, its occurrence number was on the rise, which should be taken to prevent its outbreak in farming-pastoral transitional zone in the future.
rangeland grasshopper; community construction; time dynamics;ecosystem
10.13524/j.2095-008x.2015.01.013
2014-06-20
http://www.cnki.net/kcms/detail/23.1566.T.20150119.1618.006.html
国家公益性行业(农业)科研专项资助项目(201003079)
王皙玮(1982-),女,黑龙江哈尔滨人,博士研究生,研究方向:森林生态与害虫防治;*通讯作者:王贵强(1953-),男,黑龙江哈尔滨人,博士,教授,博士研究生导师,研究方向:昆虫生态学,E-mail:GQWang53@126.com。
S154.5
A
2095-008X(2015)01-0060-09
猜你喜欢
小哥白尼(野生动物)(2022年5期)2022-08-15
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
科学大众(2020年10期)2020-07-24
河南科学(2020年3期)2020-06-02
故事作文·低年级(2019年6期)2019-06-20
收藏界(2018年1期)2018-10-10
创新作文(小学版)(2018年31期)2018-05-16
幼儿教育·父母孩子版(2017年10期)2017-12-13
摄影之友(影像视觉)(2017年1期)2017-07-18
