
丹江口水库水滨带植物群落空间分布及环境解释
2015-03-10 12:19刘瑞雪陈龙清史志华
生态学报 2015年4期
刘瑞雪,陈龙清,史志华
1 华中农业大学园艺林学学院园艺植物生物学教育部重点实验室,武汉 430070 2 深圳大学建筑与城市规划学院,深圳 518060 3 华中农业大学资源与环境学院,武汉 430070
丹江口水库水滨带植物群落空间分布及环境解释
刘瑞雪1,2,陈龙清1,*,史志华3
1 华中农业大学园艺林学学院园艺植物生物学教育部重点实验室,武汉 430070 2 深圳大学建筑与城市规划学院,深圳 518060 3 华中农业大学资源与环境学院,武汉 430070
探讨了环境因素对丹江口水库(南水北调中线水源地)水滨带植物群落空间分布的影响。通过对水滨带植物群落和环境因素的实地调查,用双向指示种分析(TWINSPAN)对201个水滨带植物群落进行分类;结合地形、土壤和水文因素用除趋势典范对应分析法(DCCA)分析环境因素对水滨带植物群落的影响;并对环境因素的解释能力进行定量分离。结果表明:(1)水滨带植物群落包括7种类型,分别是萹蓄群落、苘麻群落、细叶水芹+狗牙根群落、狗牙根群落、响叶杨-狗牙根群落、杜梨-白刺花-狗牙根群落和侧柏-牡荆-三穗苔草群落;(2)海拔和水淹影响对水滨带植物群落空间分布具有主导作用。海拔升高,水淹影响减弱,植物群落呈现由草本植物群落向木本植物群落变化的格局;(3)土壤因素的解释能力大于地形因素,水文因素的解释能力最小。各类环境因素之间存在交互作用,地形、水文和土壤因素三者间的交互作用最大,地形和土壤因素之间的交互作用最小。环境因素共解释水滨带植物群落空间分布的21.99%,未解释部分为78.01%。结果证明环境对植被的解释能力是由植被的复杂程度决定的,植被越复杂,环境的解释能力越低。
水滨带;植物群落;空间分布;环境解释;丹江口水库
植物群落的空间分布与环境之间的关系一直都是生态学研究的热点问题[1- 4]。植物群落的空间分布是自然界各种因素共同作用的结果[1,5- 6]。在大尺度上气候对植被分布起着决定性作用,在较小尺度上地形、土壤、生物之间的相互作用等因素也影响着植物群落的分布[7- 11]。水滨带是由于水位涨落而使水陆交界地带的土地周期性淹没或出露于水面的区域,包括最低水位线和最高水位线之间的区域,以及最高水位线以上可能受到水淹影响的区域[12]。虽然水滨带在地球生态系统中仅占很小比例,但其作为水生与陆地生态系统之间的过渡带,是两大生态系统相互作用的桥梁和纽带,发挥着重要的生态功能[13]。水滨带植被具有保护水域生态安全、保护水质、稳定堤岸、保持生境连通性和维持生物多样性等生态功能[14]。由于多种因素的影响[15- 18],目前世界范围的水滨带都出现了植被退化的现象,严重威胁水域的生态安全。
出于对水资源和能源的迫切需求,中国成为世界上最热衷于修建水利工程的国家之一,长江已成为世界上拥有已建或在建水利工程最多的河流[19]。水利工程的建设对社会经济发展和人民生活水平提高有重要作用,但也带来了严重的生态问题,其中之一就是造成水滨带植被的严重退化[16]。丹江口水库(32°36′—33°48′ N, 110°59′—111°49′ E)是我国南水北调中线工程的水源地,位于长江最大支流汉江和最长支流丹江的交汇处。水滨带植被是保护水质、预防水体污染的最后一道生态屏障,在水源地生态环境保护中起着重要作用。自1973年大坝建成,丹江口水库已运行了40a。经过40a的自然植被演替,丹江口水库的水滨带上已经形成明显不同于水生或陆生植被的水滨带植被。之前对丹江口水库植被的研究主要集中在浮游植物和滩涂湿地植物等方面[20- 23],针对水滨带植物群落的研究尚未见报道。研究该区域植物群落空间分布及其与环境因素的关系对丹江口水库乃至我国众多水利工程的水滨带植被保护与恢复有重要的现实意义。本文通过对丹江口水库水滨带的植物群落和环境条件进行实地调查,对植物群落的空间分布及其与环境之间的关系进行分析,并定量分离各类因素对水滨带植物群落空间分布的解释能力,目的在于为水滨带退化植被的保护与恢复重建提供理论依据和实践参考。
1 材料与方法
1.1 研究区概况
丹江口水库位于河南省和湖北省交界处,主要包括河南省淅川县的丹库和湖北省丹江口市的汉库。水库地处亚热带温暖半湿润季风区,年均气温13.7 ℃,年均蒸发量854 mm,年均降水量为873.3 mm,降雨主要集中在5—10月[23]。地带性植被主要为亚热带落叶阔叶林,以及部分常绿阔叶林和针阔混交林。植物种类主要有黑松(Pinusthunbergii)和栓皮栎(Quercusvariabilis)等乔木,牡荆(Vitexnegundovar.cannabifolia)、白刺花(Sophoradavidii)、马桑(Coriarianepalensis)和湖北算盘子(Glochidionwilsonii)等灌木,以及野艾蒿(Artemisialavandulaefolia)、狗牙根(Cynodondactylon)和白茅(Imperatacylindrica)等草本植物[24]。
1.2 样地设置与植物群落调查

图1 调查区域分布图 Fig.1 Distribution of 51 sampling sites in the riparian zone of Danjiangkou Reservoir
调查于2011年4月至5月进行。参考丹江口水库1∶1万地形图,结合高分辨率遥感影像,根据水滨带植被现状和生境特点以及可达性,在水滨带上设置51个重点调查的区域(图1),每个调查区域面积为1 km2(1 km×1 km)。在每个区域内从水面(约海拔140 m)以上有植物出现的地方起至海拔180 m的范围内,沿海拔梯度布设调查断面,每个调查断面与等高线垂直。在每条断面上,根据植被的物种组成变化划分植物群落。
记录样方中出现的所有维管植物。对乔木种测定株数、高度、盖度和胸径;对灌木种测定株数、高度和盖度。对草本植物测定高度和盖度,目测法测定多度,用Braun-Blanquet 5级制记录,非常多(个体数比例>75%)- 5,多(50%—75%)- 4,较多(25%—50%)- 3,较少(5%—25%)- 2,少(<5%)- 1。植物种类鉴定依据《中国植物志》和《湖北植物志》。
1.3 环境因素调查
本研究共选取3类环境因素,分别是地形、土壤和水文因素。地形因素包括海拔(ELEV)、坡度(SA)、坡向(ASP);土壤因素包括土层厚度(ST)、土壤质地(TEX)、土壤pH值(PH)、有机质含量(OM)、全氮含量(TN)、全磷含量(TP)、速效氮含量(AN)、速效磷含量(AP);水文因素包括年平均水淹时间(FD)和水淹频率(FF)。
在群落内用手持式GPS测定海拔,用罗盘仪测定坡度和坡向。坡向划分为5个等级并分别赋值,北坡(0°—22.5°和 337.5°—360°)为1,东北坡(22.5°—67.5°)和西北坡(247.5°—292.5°)为2,东坡(67.5°—112.5°)和西坡(292.5°—337.5°)为3,东南坡(112.5°—157.5°)和西南坡(202.5°—247.5°)为4,南坡(157.5°—202.5°)为5。数值越小表示生境条件越冷湿,光照条件越差,数值越大表示生境条件越干热,光照条件越好。
在群落内随机选点挖取土壤剖面,记录土层厚度。土层厚度分为4个等级并分别赋值,土层厚度小于20 cm为1,大于20 cm且小于40 cm为2,大于40 cm且小于60 cm为3,大于60 cm为4。根据土壤微粒的大小,将土壤质地分为6个等级并赋值,粘土1,粘壤土2,粉壤土3,壤土4,砂壤土5,砂土6。随机收集5个点0—20 cm土层中的土壤样品,充分混合为一份。样品带回实验室,根据《土壤农业化学分析方法》[25]进行土壤化学性质的测定。
根据丹江口水库的水位数据和植物群落所处的海拔,计算植物群落所处生境的水文因素(年平均水淹时间和水淹频率)。丹江口水库从1979年1月至2009年12月的水位数据由长江水资源保护科学研究所提供。
磁异常ΔT反映为中部较低、两端较高的磁场特征,并在隐爆角砾岩上方的低磁场背景中出现形态规整的弱磁异常,显示较隐伏岩体磁性较强的隐爆角砾岩的存在。F2、F3断裂分别对应两个不同磁场背景的接触带上,且分别倾向剖面中段的低磁场,显示该地段经历岩浆构造热液改造,在隐爆角砾岩和上述构造的虚脱部位是成矿最为有利的地段,据此圈定深部物探找矿靶区2个(图6),均表现为低磁和低阻异常特征,推测已知矿体为其外带的断裂(F3)控矿。
1.4 数据处理
以植物群落为单位,分乔木层、灌木层和草本层来计算物种的重要值。以物种重要值为基础,建立物种-样方矩阵。根据实地调查和室内实验分析,建立环境变量-样方矩阵。先对植物群落进行TWINSPAN划分植物群落类型,再进行DCCA排序分析植物群落与环境的关系。然后采用Borcard等[26]的方法分离地形、土壤和水文三类环境因素对植物群落的影响。TWINSPAN分析由WinTWINS 2.3 完成[27],DCCA分析由CANOCO for Windows 4.5 完成[28- 29],排序图由CanoDraw for Windows 4.5 绘制。
2 结果与分析
2.1 植物群落类型划分
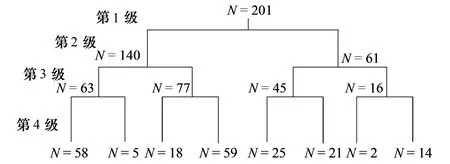
图2 201个水滨带植物群落TWINSPAN分类图 Fig.2 TWINSPAN dendrogram of 201 riparian plant communities
以物种-样方矩阵为基础,去除偶见种的影响,使用TWINSPAN对201个植物群落进行群落类型的划分。根据TWINSPAN的结果(图2),采取第4级水平的划分,并结合实际生态意义,将201个植物群落分为7种植物群落类型。依据植物群落每一层中平均重要值较大的优势物种对群落进行命名,7种植物群落类型的描述如下:
(1)萹蓄(Polygonumaviculare)群落
草本植物群落,主要包括萹蓄、狗牙根、苘麻、朝天委陵菜(Potentillasupine)和小藜(Chenopodiumserotinum),其中萹蓄是平均重要值最大的物种。伴生种有狗尾草(Setariaviridis)、小巢菜(Viciahirsute)、盾果草(Thyrocarpussampsonii)、红蓼(Polygonumorientale)、野老鹳草(Geraniumcarolinianum)等。分布在水滨带下部,地势平缓,水淹影响严重,环境极其湿润,包含58个植物群落共56个物种。
(2)苘麻(Abutilontheophrasti)群落
草本植物群落,苘麻为最具优势的物种。狗牙根和萹蓄在该类型所有群落中都出现,且在个别群落中有较大的重要值。伴生种有绵毛酸模叶蓼(Polygonumlapathifoliumvar.salicifolium)、稗(Echinochloacrusgalli)、打碗花(Calystegiahederacea)、朝天委陵菜(Potentillasupine)、细叶水芹、小巢菜和石龙芮(Ranunculussceleratus)等。主要分布在水滨带下部,地势平缓,水淹影响严重,环境湿润,包括5个植物群落,物种共计10个。
(3)细叶水芹(Oenanthedielsiivar.stenophylla)+ 狗牙根(Cynodondactylon)群落
草本植物群落,主要包括细叶水芹、狗牙根、菵草(Beckmanniasyzigachne)、小藜和苘麻,其中细叶水芹和狗牙根是最具优势地位的物种,平均重要值大于10。伴生种众多,如泽漆(Euphorbiahelioscopia)、鹅观草(Roegneriakamoji)、四叶葎(Galiumbungei)、水蓼(Polygonumhydropiper)、短叶水蜈蚣(Kyllingabrevifolia)等。主要分布在水滨带中下部,地势较平坦,受水淹影响较大,环境潮湿。包括18个植物群落,物种共计60个。
(4)狗牙根(Cynodondactylon)群落
草本植物群落,狗牙根是最具优势的物种,平均重要值大于30。平均重要值较大的物种还有细叶水芹、小藜、朝天委陵菜、打碗花、苘麻和野老鹳草。平均重要值较小的伴生种众多,如附地菜(Trigonotiapeduncularis)、猪殃殃(Galiumaparinevar.tenerum)、水苦荬(Veronicaundulata)、蚤缀(Arenariaserpyllifolia)和虉草(Phalarisarundinacea)等。主要分布在水滨带中部,地形稍陡,环境较潮湿,包括59个植物群落,物种共计72个。
(5)响叶杨(Populusadenopoda)-狗牙根(Cynodondactylon)群落
具有乔灌草复合结构,乔木层主要包括响叶杨、柘树(Cudraniatricuspidata),其中响叶杨是群落中最优势的物种;灌木层包括小果蔷薇(Rosacymosa)、野山楂(Crataeguscuneata)和算盘子(Glochidionpuberum)等,但平均重要值都较低;狗牙根是草本层最主要的物种,伴生种还有细叶水芹、朝天委陵菜、鼠麴草(Gnaphaliumaffine)和小苜蓿(Medicagominima)等。分布在水滨带上部,地形陡峭,受水淹影响小,环境较干燥,包括24个植物群落,物种共计94个。
(6)杜梨(Pyrusbetulifolia)-白刺花(Sophoradavidii)-狗牙根(Cynodondactylon)群落
具有乔灌草复合结构,乔木层包括黑松(Pinusthunbergii)、杜梨和构树(Broussonetiapapyrifera),其中杜梨是最优势的物种;灌木层主要包括白刺花(Sophoradavidii)、牡荆(Vitexnegundovar.cannabifolia)、算盘子等。狗牙根、白茅(Imperatacylindrica)和野艾蒿(Artemisialavandulaefolia)是草本层的主要物种,平均重要值大于10。大量的伴生种,如刺儿菜(Cirsiumsetosum)、菅草(Themedavillosa)、翻白草(Potentilladiscolor)、苍耳(Xanthiumsibiricum)和风轮菜(Clinopodiumchinense)等。分布在水滨带上部,地形陡峭,水淹影响较小,环境干燥,包括21个植物群落,物种共计95个。
(7)侧柏(Platycladusorientalis)-牡荆(Vitexnegundovar.cannabifolia)-三穗苔草(Carextristachya)群落
具有乔灌草复合结构,包括16个植物群落,物种112个,是7种群落类型中包含物种数目最多的。乔木层包括侧柏、刺槐(Robiniapseudoacacia)、柏木(Cupressusfunebris)和乌桕(Sapiumsebiferum),其中侧柏是该植物群落类型中最具优势的物种;灌木层主要由牡荆、小果蔷薇、竹叶椒(Zanthoxylumplanispinum)等组成,牡荆是灌木层平均重要值最大的物种;三穗苔草是草本层最重要的物种。伴生种有构树、千里光(Senecioscandens)、野菊(Dendranthemaindicum)等。主要分布在水滨带上部,地形陡峭,基本不受水淹影响,环境十分干燥。
2.2 植物群落与环境的关系
丹江口水库水滨带植物群落与环境因素的DCCA排序结果见表1。前4个典范轴解释了植被变化的12.7%。Monte Carlo随机置换检验表明,所有典范特征轴与植物群落变化都是极显著相关的(P=0.001)。因DCCA排序前两轴的特征值之和占全部排序轴特征值总和的79.06%,包含了排序的绝大部分信息,其中轴1的特征值占全部特征值总和的60.20%,因此采用前两轴的数据来分析植物群落与环境因素的关系(图3)。根据13个环境变量与DCCA排序轴的相关性分析可知(表2),海拔、年平均水淹时间和水淹频率与DCCA第1轴相关性最强,表明在水滨带上海拔和水淹影响是决定植物群落空间分布的主导因子。土壤pH、有机质含量和全氮含量与DCCA第2轴显著相关,表明土壤养分在决定水滨带植物群落的空间分布上也有一定作用。
根据DCCA排序图可知(图3),各植物群落在排序空间中的位置反映了群落的生态学特征。沿第1轴从右到左,海拔逐渐升高,水淹时间和水淹频率降低,植物群落类型从草本植物群落过渡到木本植物群落。其中草本植物群落分布在较低的海拔上,受到长时间和高频率的水淹影响;木本植物群落分布在较高的海拔上,较少或几乎不受水淹的影响。群落类型沿海拔和水淹影响,从右至左依次分布萹蓄群落、苘麻群落、细叶水芹+狗牙根群落、狗牙根群落、响叶杨-狗牙根群落、杜梨-白刺花-狗牙根群落和侧柏-牡荆-三穗苔草群落,各种类型之间存在着不同程度的重叠。其中萹蓄群落生长在水滨带最下部,受水淹影响最严重的区域;侧柏-牡荆-三穗苔草群落则主要生长在水滨带上部边缘,几乎不受水淹影响。沿第二轴从下到上,土壤pH值、有机质含量和全氮含量增加。植物群落在第2轴上变化没有第1轴明显,这一结果与表1所反映的结果一致,说明海拔和水淹影响对水滨带植物群落空间分布所起的作用远远大于土壤养分的作用,第1轴能很好的解释水滨带植物群落与环境的相互关系。结果表明尽管植物群落的空间分布是多种因素综合作用的结果,但在丹江口水库水滨带上海拔和水淹影响在植物群落空间格局的形成中起着主导作用。

表1 丹江口水库水滨带植物群落DCCA排序Table 1 DCCA ordination of plant communities in the riparian zone of Danjiangkou Reservoir
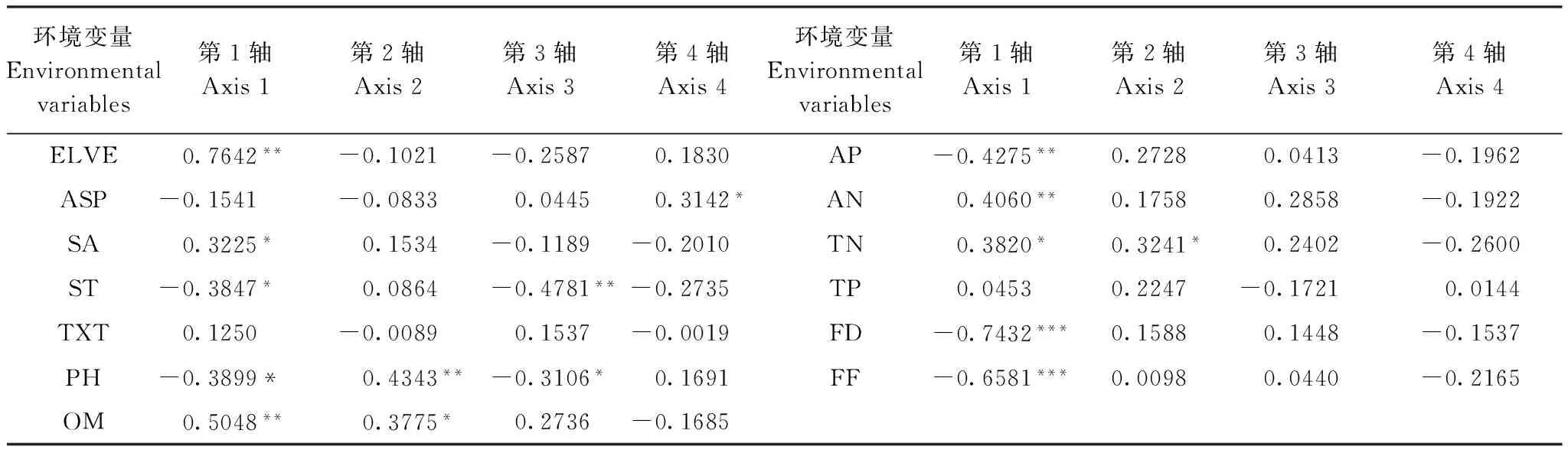
表2 13个环境变量与DCCA排序轴的相关性Table 2 Correlation coefficients between 13 environmental variables and DCCA ordination axes
*、**和***分别表示在0.05、0.01和0.001水平上的显著性; ELEV:海拔Elevation;SA: 坡度Slope;ASP: 坡向Aspect;ST: 土层厚度Soil thickness;TEX: 土壤质地Soil texture;pH: 土壤pH Soil pH;OM: 有机质含量Organic matter;TN: 全氮含量Total nitrogen;TP: 全磷含量Total phosphorus;AN: 速效氮含量Available nitrogen;AP: 速效磷含量Available phosphorus;FD: 年平均水淹时间 Flood duration; FF: 水淹频率Flood frequency
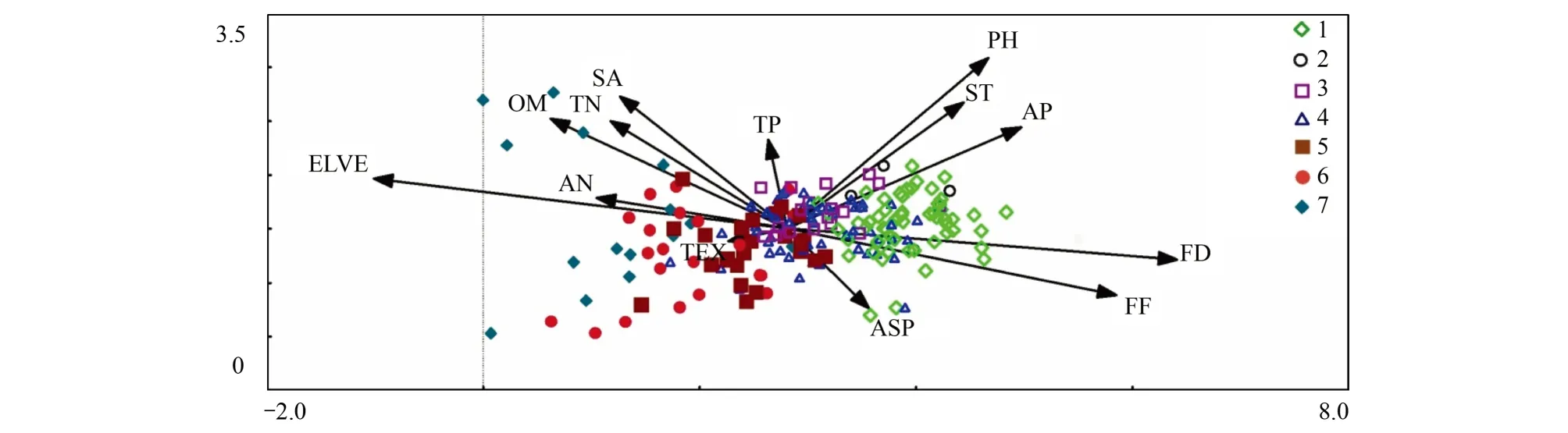
图3 水滨带植物群落的DCCA排序图Fig.3 DCCA ordination of 201 riparian plant communities1:萹蓄群落,2:苘麻群落,3:细叶水芹+狗牙根群落,4:狗牙根群落,5:响叶杨-狗牙根群落,6:杜梨-白刺花-狗牙根群落,7:侧柏-牡荆-三穗苔草群落; ELEV:海拔,SA: 坡度,ASP: 坡向,ST: 土层厚度,TEX: 土壤质地,PH: 土壤pH ,OM: 有机质含量,TN: 全氮含量,TP: 全磷含量,AN: 速效氮含量,AP: 速效磷含量,FD; 年平均水淹时间,FF: 和水淹频率
2.3 植被群落空间分布的环境因素分离
通过多次排序,将环境因素对水滨带植物群落空间分布的解释能力进行定量分离,结果如表3。由表3可知,所收集的3类环境因素共解释水滨带植物群落空间分布格局变化的21.99%,剩下的78.01%属于未解释部分。地形因素的解释能力为7.96%,其中3.15%为纯地形因素引起,余下的4.81%是地形因素与土壤和水文因素的交互作用引起的;由土壤因素引起的植物群落空间格局变化占总变化的14.58%,其中10.24%为纯土壤因素引起的,4.34%是土壤因素与地形和水文因素的交互作用引起的;水文因素的解释能力为7.86%,其中3.04%为纯水文因素引起的变化,余下的4.82%是水文因素与土壤和地形因素的交互作用引起的。水滨带上环境因素对植物群落的影响,既有各类因素的单独作用,又有相互之间的耦合作用。地形和土壤因素的交互作用为0.73%,土壤和水文因素的交互作用为0.75%,地形和水文因素的交互作用为1.22%,3类环境因素的共同作用为2.86%。结果表明水滨带植物群落的空间分布是长期生态过程中植物与环境相互作用的结果。

表3 约束排序和环境因素分解Table 3 Results of DCCA and partitioning of environmental variables
3 讨论
3.1 丹江口水库水滨带植物群落的空间分布
水滨带是典型的生态过渡带,是生态系统中能量流动和物种循环的活跃地带。作为生态过渡带,水滨带有着高度异质的环境条件、复杂的生态过程和多样的生物群落。水滨带的植物群落在物种组成、群落结构、空间分布等方面明显不同于相邻的陆地或者水生植物群落[30- 31]。以草本植物群落为主,木本植物群落较少,反映出水滨带受到持续不断的环境干扰,这些干扰造成水滨带群落结构趋于简单化[32]。尽管植物群落的空间分布格局是多种环境因素综合作用的结果,但水滨带上海拔和水淹影响是决定植物群落分布的主导因素。在丹江口水库水滨带上 7种植物群落类型主要沿海拔和水淹影响梯度分布,萹蓄群落、苘麻群落、细叶水芹+狗牙根群落、狗牙根群落分布在水滨带的中下部,物种组成和群落结构较简单。物种组成上以草本植物为主,几乎没有木本植物;群落结构上都是单一草本层的结构。响叶杨-狗牙根群落、杜梨-白刺花-狗牙根群落和侧柏-牡荆-三穗苔草群落分布在水滨带的上部,物种组成上包含多种生活型的物种,群落结构上以乔灌草复层结构为主。造成水滨带植物群落这种空间分布格局的主要原因是植物对环境影响的反映不同。水滨带低海拔生境受到较长时间和较高频率的水淹影响,而且土壤养分较低,严酷的环境条件阻碍植物生长,特别是高大木本植物的生长[12,33- 34]。而低矮的草本植物具有较强的环境适应性和较宽的生态幅,成为水滨带低海拔上的优势物种。水滨带高海拔区域受到水淹影响较小,土壤养分含量也较多,这样的生境条件适宜大多数植物的生长。适宜的生境条件增强了物种对资源的竞争,如对光和土壤养分的竞争。木本植物具有较大的生物量,竞争能力明显强于低矮的草本植物[35],因而在水滨带高海拔区域木本植物占据优势地位。
3.2 环境因素的相对重要性
使用约束排序对3类环境因素进行植物群落空间分布解释能力的定量分离。据表3可知,3类环境因素对植物群落空间分布的独立解释能力低于混合解释能力,且它们的解释能力存在重叠效应。这一结论证明了Brocard等[26]的观点:不同类别的环境因素之间存在复杂的相互作用。根据本研究的结果,无论是独立解释能力还是混合解释能力,土壤因素的解释能力总是最强,地形因素的解释能力次之,水文因素的解释能力最低。出现这一结果的原因是各类环境因素中所包含的因子数目不同,土壤因素包括8个因子,因此解释能力远高于地形因素和水文因素的解释能力。3类环境因子共解释水滨带植物群落空间分布的21.99%,未解释部分为78.01%。综合前人的研究成果,沈泽昊和张新时[8]对三峡大老岭常绿阔叶林的研究发现,环境因素解释了森林群落空间分布的9.5%;在对岷江上游干旱河谷植物群落的研究中,环境因素解释了灌丛群落变化的21.47%[6];对喀斯特峰丛洼地热带混交林的研究发现,环境因素解释了森林群落变化的39.16%[36];在松嫩平原草地植物群落中环境因素解释了植被变化的69.10%[37];张庆等[4]对内蒙古荒漠草原短花针茅群落的研究发现,环境因素解释了荒漠草原植物群落变化的70.7%。可见,植被类型从常绿阔叶林到荒漠草原,复杂性降低,环境的解释能力增大。丹江口水库水滨带植被的复杂程度与干旱河谷灌丛植物群落类似。根据沈泽昊和张新时[8]的观点,环境因素对于植被变化的可解释程度主要是由植被本身的复杂性决定的,植被越简单,说明环境对植被的可控性越大,环境因素的解释能力越高;植被越复杂,环境的可解释程度越低。水滨带是典型的生态过渡带,环境条件复杂,植被类型变化多样,这些原因共同造成了环境因素的未解释部分较高。未解释的部分可能包括气候因素、生物相互作用因素、干扰因素和随机因素等。
致谢:华中农业大学艾蕾、庞宏东、王万平、张辛阳、詹娟、朱良艳、吴韩、黄旭东、段诚和袁岸琼在植物群落和环境因素调查中给予帮助。感谢华中农业大学园艺林学学院园艺植物生物学教育部重点实验室刘秀群老师对写作的帮助。
[1] Legendre P, Fortin M J. Spatial pattern and ecological analysis. Vegetation, 1989, 80(2): 107- 138.
[2] Brooker R W. Plant-plant interactions and environmental change. New Phytologist, 2006, 171(2): 271- 284.
[3] Clark C M, Cleland E E, Collins S L, Fargione J E, Gough L, Gross K L, Pennings S C, Suding K N, Grace J B. Environmental and plant community determinants of species loss following nitrogen enrichment. Ecology Letters, 2007, 10(7): 596- 607.
[4] Zhang Q, Niu J M, Buyantuyev A, Ding Y, Kang S R L, Wang F L, Zhang Y N, Yang Y, Han Y J. Ecological analysis and classification of Stipa breviflora communities in the Inner Mongolia region: the role of environmental factors. Acta Prataculturae Sinica, 2012, 21(1): 83- 92.
[5] Burke A. Classification and ordination of plant communities of the Naukluft mountains, Namibia. Journal of Vegetation Science, 2001, 12(1): 53- 60.
[6] 张文辉, 卢涛, 马克明, 周建云, 刘世梁. 岷江上游干旱河谷植物群落分布的环境与空间因素分析. 生态学报, 2004, 24(3): 552- 559.
[7] 牛建明. 内蒙古主要植被类型与气候因子关系的研究. 应用生态学报, 2000, 11(1): 47- 52.
[8] 沈泽昊, 张新时. 三峡大老岭地区森林植被的空间格局分析及其地形解释. 植物学报, 2000, 42(10): 1089- 1095.
[9] Abd El-Ghani M M, Amer W M. Soilvegetation relationships in a coastal desert plain of southern Sinai, Egypt. Journal of Arid Environments, 55(4): 607- 628.
[10] 贺强, 崔保山, 赵欣胜, 付华龄, 廖晓琳. 黄河河口盐沼植被分布、多样性与土壤化学因子的相关关系. 生态学报, 2009, 29(2): 676- 687.
[11] 陈宝瑞, 李海山, 朱玉霞, 李刚, 辛晓平, 张宏斌, 周磊. 呼伦贝尔草原植物群落空间格局及其环境解释. 生态学报, 2010, 30(5): 1265- 1271.
[12] Naiman R J, Decamps H. The ecology of interfaces: Riparian zones. Annual Review of Ecology and Systematics, 1997, 28(1): 621- 658.
[13] Gregory S V, Swanson F J, McKee W A, Cummins K W. An ecosystem perspective of riparian zones: Focus on lines between land and water. Bioscience, 1991, 41(8): 540- 551.
[14] Gunderson P, Laurén A, Finér L, Ring E, Koivusalo H, Stersdal M, Weslien J O, Sigurdsson B D, Högbom L, Laine J, Hansen K. Environmental services provided from riparian forests in the Nordic countries. Ambio, 2010, 39(8): 555- 566.
[15] Nilsson C, Jansson R, Zinko U. Long-term responses of river-margin vegetation to water-level regulation. Science, 1997, 276(5313): 798- 800.
[16] New T, Xie Z Q. Impacts of large dams on riparian vegetation: applying global experience to the case of China′s Three Gorges Dam. Biodiversity and Conservation, 2008, 17(3): 3149- 3163.
[17] Pennington D N, Hansel J R, Gorchov D L. Urbanization and riparian forest woody communities: Diversity, composition, and structure within a metropolitan landscape. Biological Conservation, 2010, 143(1): 182- 194.
[18] Yang J, Dilts T E, Condon L A, Turner P L, Weisberg P J. Longitudinal-and transverse-scale environmental influences on riparian vegetation across multiple levels of ecological organization. Landscape Ecology, 2011, 26(3): 381- 395.
[19] 王勇, 刘义飞, 刘松柏, 黄宏文. 三峡库区消涨带植被重建. 植物学通报, 2005, 22(5): 513- 522.
[20] 邬红娟, 彭建华, 韩德举, 简东, 邹清. 丹江口水库浮游植物及其演变. 湖泊科学, 1996, 8(1): 43- 50.
[21] 刘文治, 张全发, 刘贵华, 李天煜, 李伟, 吴文颖. 丹江口库区湿地水生维管束植物的区系研究. 武汉植物学研究, 2005, 23(5): 449- 454.
[22] 刘文治, 张全发, 李天煜, 李伟, 吴文颖, 刘贵华. 丹江口库区湿地植被的数量分类和排序. 武汉植物学研究, 2006, 24(3): 220- 224.
[23] 刘文治, 卜红梅, 刘贵华, 张全发. 丹江口库区湿地植被对南水北调中线工程响应研究. 水土保持通报, 2009, 29(4): 149- 152.
[24] 刘瑞雪, 詹娟, 史志华, 陈龙清. 丹江口水库消落带土壤种子库与地上植被和环境的关系. 应用生态学报, 2013, 24(3): 801- 808.
[25] 鲁如坤. 土壤农业化学分析方法. 南京: 河海大学出版社, 2000.
[26] Borcard D, Legendre P, Drapeau P. Partialling out the spatial component of ecological variation. Ecology, 1992, 73(3): 1045- 1055.
[27] Hill M O,milauer P. Twinspan for Windows Version 2. 3. Center for Ecology and Hydrology & University of South Bohemia, Huntingdon & Ceske Budejovice, Czekh, 2005.
[28] Leps J, Smilauer P. Multivariate Analysis of Ecological Date Using CANOCO. Cambridge: Cambridge University Press, 2003.
[29] ter Braak C J F,milauer P. CANOCO Reference Manual and CanoDraw for Windows User′s Guide: Software for Canonical Community Ordination (Version 4. 5). New York, Ithaca: Micro Computer Power, 2002.
[30] 尚宗波, 高琼. 流域生态学-生态学研究的一个新领域. 生态学报, 2001, 21(3): 468- 473.
[31] 代力民, 王青春, 邓红兵, 陈高, 王庆礼. 二道白河河岸带植物群落最小面积与物种丰富度. 应用生态学报, 2002, 13(6): 641- 645.
[32] LaPaix R, Freedman B. Vegetation structure and composition within Urban Parks of Halifax Regional Municipality, Nova Scotia, Canada. Landscape and Urban Planning, 2010, 98(2): 124- 135.
[33] Ali M M. Plant functional types in Lake Nubia in relation to physicogeographic factors. Limnologica-Ecology and Management of Inland Waters, 2003, 33(4): 305- 315.
[34] Capon S J. Flood variability and spatial variation in plant community composition and structure on a large arid floodplain. Journal of Arid Environment, 2005, 60(2): 283- 302.
[35] Roem W J, Berendse F. Soil acidity and nutrient supply ratio as possible factors determining changes in plant species diversity in grassland and heathland communities. Biological Conservation, 2000, 92(2): 151- 161.
[36] 宋同清, 彭晚霞, 曾馥平, 王克林, 覃文更, 谭卫宁, 刘璐, 杜虎, 鹿士杨. 木论喀斯特峰丛洼地森林群落空间格局及环境解释. 植物生态学报, 2010, 34(3): 298- 308.
[37] 辛晓平, 高琼, 李镇清, 杨正宇. 松嫩平原碱化草地植物群落分布的空间和环境因素分析. 植物学报, 1999, 41(7): 775- 781.
Spatial distribution of plant communities and environmental interpretation in the riparian zone of Danjiangkou Reservoir
LIU Ruixue1,2, CHEN Longqing1,*, SHI Zhihua3
1KeyLaboratoryofHorticulturalPlantBiology(MinistryofEducation),CollegeofHorticultureandForestryScience,HuazhongAgricultureUniversity,Wuhan430070,China2CollegeofArchitectureandUrbanPlanning,ShenzhenUniversity,Shenzhen518060,China3CollegeofResourcesandEnvironment,HuazhongAgriculturalUniversity,Wuhan430070,China
Riparian vegetation performs important ecological functions in protecting water quality, stabilizing the bank, maintaining habitats connectivity and enhancing local biodiversity. Yet, riparian vegetation has been degraded severely throughout the world mainly due to anthropogenic disturbances. This study focused on the riparian vegetation communities and the impact of environmental factors on spatial distribution of plant communities in the riparian zone of Danjiangkou Reservoir, the water source for the Middle-Route of the South-to-North Water Transfer Project. Field survey was carried out from April to May, 2011. The 201 plant communities were investigated. And three topographic factors (elevation, slope and aspect), eight soil factors (soil thickness, soil texture, soil pH, organic matter, total nitrogen, total phosphorus, available nitrogen, and available phosphorus), and two hydrological factors (flooding duration and flooding frequency) were measured. On the basis of the investigation of riparian plant communities and environmental factors, the 201 plant communities were classified into different community types by means of Two Way Indicator Species Analysis (TWINSPAN); the impact of environmental factors on riparian plant communities were analyzed by using Detrended Canonical Correspondence Analysis (DCCA) combined with topographic, hydrologic and soil factors. And the explanatory power of environmental factors on the riparian plant community was quantitatively partitioned. The results showed that (1) the riparian plant communities were classified into seven types ofPolygonumavicularecommunity,Abutilontheophrasticommunity,Oenanthedielsiivar.stenophylla+Cynodondactyloncommunity,Cynodondactyloncommunity,Populusadenopoda-Cynodondactyloncommunity,Pyrusbetulifolia-Sophoradavidii-Cynodondactyloncommunity andPlatycladusorientalis-Vitexnegundovar.cannabifolia-Carextristachyacommunity; (2) the results of DCCA indicated that riparian vegetation and environmental factors had a significant correlation. The first axis had a significant correlation with elevation, flood duration and flood frequency, it indicated that elevation and flooding played a dominant role in spatial distribution of the riparian plant communities. Soil organic matter content, pH and total nitrogen content presented a significant correlation with the second axis, which indicated that soil nutrient also played a role in spatial distribution of the riparian plant communities. (3) the effects of soil factors, hydrology factors, topographic factors and their interaction on the total variation of riparian vegetation were quantitatively partitioned and showed that the contribution rate of the soil factors was 10.24% separately, and the contribution rate of the topographic factors was 3.15% and the hydrological factors was 3.04% separately. 0.73% for soil factors coupled with topographic factors, 1.22% for topographic factors and hydrological factors, 0.75% for hydrology factors with soil factors, and the interactive effect of topographic, soil and hydrological factors was 2.86%. The collected environmental factors accounted for 21.99% of spatial distribution of the riparian plant communities, the unexplained portion for 78.01%. Our results demonstrated that the explanatory power of environmental factors was determined by the complexity of vegetation, and the more complexity of vegetation, the less explanatory power by environmental factors. The riparian zone is a typical ecotone, and environment and vegetation are complicated and varied. These caused that the explanatory power of environmental factors was low. The unexplained portion may be caused by climate, biotic interaction, interference and random factors.
riparian zone; plant community; spatial distribution; environmental interpretation; Danjiangkou Reservoir
国家“十二五”科技支撑计划项目(2012BAC06B03)
2013- 04- 23;
日期:2014- 04- 11
10.5846/stxb201304230786
*通讯作者Corresponding author.E-mail: chenlq@mail.hzau.edu.cn
刘瑞雪,陈龙清,史志华.丹江口水库水滨带植物群落空间分布及环境解释.生态学报,2015,35(4):1208- 1216.
Liu R X, Chen L Q, Shi Z H.Spatial distribution of plant communities and environmental interpretation in the riparian zone of Danjiangkou Reservoir.Acta Ecologica Sinica,2015,35(4):1208- 1216.
猜你喜欢
汽车实用技术(2022年16期)2022-09-03
小哥白尼(军事科学)(2018年3期)2018-06-15
科学与财富(2018年8期)2018-05-09
现代园艺(2017年21期)2018-01-03
绿色科技(2017年10期)2017-07-05
科技资讯(2017年7期)2017-05-06
制造技术与机床(2016年7期)2016-08-31
河南科技(2015年3期)2015-02-27
园艺与种苗(2015年10期)2015-02-27
当代畜禽养殖业(2014年5期)2014-08-31
