
科尔沁草甸生态系统水分利用效率及影响因素
2015-03-10 08:41:06李辉东关德新袁凤辉王安志金昌杰吴家兵井艳丽
生态学报 2015年2期
李辉东,关德新,袁凤辉,王安志,金昌杰, 吴家兵,李 峥,井艳丽
1 森林与土壤生态国家重点实验室, 中国科学院沈阳应用生态研究所, 沈阳 110016 2 中国科学院大学, 北京 100049
科尔沁草甸生态系统水分利用效率及影响因素
李辉东1,2,关德新1,*,袁凤辉1,王安志1,金昌杰1, 吴家兵1,李 峥1,2,井艳丽1,2
1 森林与土壤生态国家重点实验室, 中国科学院沈阳应用生态研究所, 沈阳 110016 2 中国科学院大学, 北京 100049
生态系统水分利用效率(WUE)是衡量碳水循环耦合程度的重要指标。利用科尔沁温带草甸草地碳水通量观测数据,对该生态系统总初级生产力水分利用效率(WUEGPP)的日季变化规律及对环境和生理因子的响应进行分析。结果表明:(1)WUEGPP日变化呈下降-稳定-上升的变化趋势,最大值出现在日出后1—2 h,阴天条件下WUEGPP高于晴天,生长中期WUEGPP高于生长初期和末期;(2)总初级生产力、总蒸散和WUEGPP季节变化均呈夏季高、春秋低的形式,生长季平均值分别为0.57 mg m-2s-1、0.08 g m-2s-1和5.97 mg/g,最大值分别为1.49 mg m-2s-1、0.16 g m-2s1和13.62 mg/g;(3)总初级生产力与饱和差、气温和叶面积指数均呈二次曲线关系,与冠层导度呈对数曲线关系;总蒸散与气温呈二次曲线关系,与饱和差、叶面积指数和冠层导度相关性均不显著;(4)WUEGPP与饱和差、气温和叶面积指数均呈二次曲线关系,与冠层导度呈对数曲线关系,饱和差、冠层导度和叶面积指数分别为2.0 kPa、0.0015 m/s和4.2是控制WUEGPP增加的阈值;(5)净生态系统生产力水分利用效率(WUENEP)和净初级生产力水分利用效率(WUENPP)季节变化规律与WUEGPP一致,均值分别为3.47和5.47 mg/g。
水分利用效率; 总初级生产力; 蒸散发; 影响因子
作为表征生态系统碳水耦合程度的重要指标之一,水分利用效率将生态系统的光合生产与蒸散耗水联系在一起,反映了生态系统碳水循环规律及其相互作用关系[1]。高的水分利用效率意味着可以利用有限的水资源获得更多的光合产物,生态系统的生产能力高,对人类的生产生活更具意义。如何有效的利用现有水资源,提高生态系统生产力和水分利用效率,一直是国内外学者探求的目标[2- 3]。全球变化背景下开展生态系统碳水通量的耦合机制及其对环境变化响应规律的研究,有助于掌握植物对气候变化的响应和水分利用策略,对预测陆地生态系统在未来气候情景下的结构和功能转变,应对全球变化都具有非常重要的科学意义[4]。
早期,国内外对水分利用效率的研究多集中在农作物,且研究方法较为简单,主要采用田间直接测定法[5]和气体交换法[6]。随着以稳定碳同位素(Stable Carbon Isotope)和涡度相关(Eddy Covariance)为代表的新技术的出现和应用,植物水分利用效率的测定方法得到了不断的补充和发展,且研究对象和尺度也在不断扩大[7- 8]。尤其是涡度相关方法以其对植物的破坏小和快速实时测定等优势[9]而得到青睐,近年来发展迅速,它可以直接测定生态系统碳水通量,获得植物群体的水分利用效率,在考察生态系统中短期内水分利用效率对环境变化的响应方面得到了广泛应用[10]。
草原约占全球陆地生态系统总面积的40%以上,是畜牧业生产和人类栖息生存的重要基地,同时,草地生态系统生态环境脆弱,对气候变化的响应较为敏感[11],是全球变化研究的热点。目前,在叶片、个体水平,无论研究对象属于哪类生态系统,其水分利用效率的国内外研究报道都较多[12- 15],在群体水平,森林生态系统、农田生态系统水分利用效率的相关报道远多于草地生态系统[16]。科尔沁温带草甸是内蒙古草地的重要组成部分,也是我国重要的畜牧业生产基地,但由于长期实行粗放经营的管理模式,草地不断退化,盐碱化和沙漠化趋势明显[17],同时该地区位于气候变化的敏感区域,对全球变化的响应十分显著[18]。开展科尔沁草甸生态系统水分利用研究,有助于科学制定水资源利用规划,合理配置水资源,提高生产力,同时增强应对全球变化的能力,为草地的脆弱性和适应性研究及草地生态系统的优化管理提供理论依据。
本文利用科尔沁温带草甸2012年4月到2012年9月涡度相关碳水通量观测数据,分析该生态系统的水分利用效率特征及影响因子,揭示了水分利用效率的日季变化规律及其对环境因子、叶面积指数和冠层导度的响应模式。
1 研究地区与研究方法
1.1 研究区概况
研究在科尔沁左翼后旗努古斯台苏木西哈伦嘎查(43°17′N, 122°16′E,海拔203 m)的草甸区进行。研究区属于温带大陆性季风气候,雨热同期,四季分明,积温充足。全年总辐射5035.1 MJ/m2,年均温6.2 ℃,年降水量378.8 mm,降水多集中在6—9月,约占全年降水量的70%,年蒸发量1755.7 mm。研究区观测东西长约3 km,南北宽约1.5 km,地势平坦开阔。其北部与农田和退化草地相邻,南部与稀疏的村庄和沙丘相接。土壤类型为草甸土,土层深度40—100 cm,土壤有机质含量可达8%—10%。植物群落分布较为均匀,优势种及伴生种以芦苇(Phragmitesaustralis)、羊草(Leymuschinensis)、野大豆(Glycinesoja)、车前(Plantagoasiatica)、小叶毛茛(Ranunculusmicrophyllus)为主。植被于4月末萌动发芽后进入生长期,7月末平均株高达0.5 m,最大高度平均可达0.8 m。9月初人工收割后留有5 cm左右的残茬。
1.2 观测方法
1.2.1 涡度相关和微气象观测
观测点建有高4 m的微气象观测塔,开路涡度相关系统(OPEC)安装在2.5 m高度,包括三维超声风速仪(CSAT3,Campbell,USA)和CO2/H2O红外气体分析仪(Li7500,LiCor Inc, USA)。脉动信号采样频率为10 Hz,通过数据采集器(CR3000,Campbell,USA)采集并按30 min计算通量平均值进行存储。塔上还安装了2层空气温湿度传感器(HMP45C,Vaisala,Finland),高度分别为1.5和3 m,净辐射传感器(NR-LITE,Kipp&Zonen,Netherlands)安装在1.5 m高度处。4个土壤温度传感器(105Tand107T, Campbell, USA)分别安装在0、5、10、20 cm土层,2个土壤含水量探头(Modell09,Campbell,USA)分别安装在5和20 cm深度,2个土壤热通量板(HFP01 and HFP01SC,HukseFlux,Netherlands) 安装在5 cm深度。
1.2.2 叶面积指数观测
在生长季5—8月份,每隔15—20d进行野外试验地采样。采样时随机选取5个1 m×1 m的样方,将样方内的植物用剪刀齐地面剪下,分出芦苇、羊草、具芒碎米莎草(Cyperusmicroiria)、野大豆和车前,每个物种选取代表性植株30株,摘下其叶片,记录各物种的叶片数。将叶片放在1 mm×1 mm方格的坐标纸上,用800万像素数码相机进行拍照后,在Photoshop软件中打开叶片照片,选择工具栏中的“缩放工具”,将图片放大到视野内目标叶片最大可视范围。然后粗选照片中的“连续的”,点击目标叶片将其外廓选中。然后将图片放大到100%,用“多边形套索工具”选择超过叶片范围以外的选定区域,将目标叶片以外的区域剪掉。最后查看“窗口-直方图”中叶片的像素值并记录。根据叶片像元与方格像素值之间的对应关系,求出单叶片的实际面积[19]。再根据下式计算总叶面积At:
(1)
式中,Si为第i个物种的平均单叶面积;Ni为第i个物种的叶片数量。
叶面积指数(LAI)可由下式计算得到:
(2)
式中,At为总叶面积(cm2);As为所采样叶的占地面积(cm2)。
1.3 计算方法
1.3.1 总初级生产力和水分利用效率的计算
对水分利用效率的定义和理解不同,或是所采用仪器及试验方法的不同,其结果表达也各不相同[1]。本文采用生态系统总初级生产力(GPP)与蒸散量(ET)的比值来定义生态系统水分利用效率WUEGPP,表达式为:
(3)
式中,ET通过涡度相关系统直接观测,GPP则通过下式进行计算:
GPP=NEP+Ri
(4)
式中,NEP为净生态系统生产力,可由涡度相关系统直接观测,“-”表示吸收;Re为生态系统呼吸量。
1.3.2 总生态系统呼吸量的计算
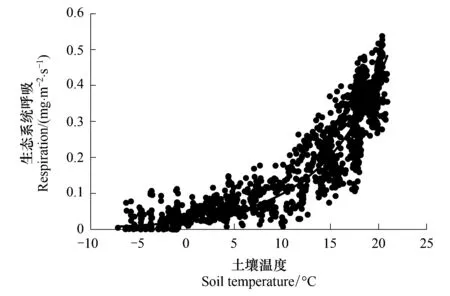
图1 科尔沁草甸夜间生态系统呼吸与土壤温度的关系Fig.1 Relationship between ecosystem respiration at night and soil temperature over Horqin Meadow
Re主要受土壤温度控制,利用土壤温度函数Q10模型来表达。夜间生态系统光合作用停止,生态系统碳通量净交换通常被认为是总生态系统呼吸量,可以利用夜间的涡度相关碳通量与土壤温度建立Re估算的经验关系式,由于Re主要受土壤温度单因子控制,因此该关系式也可以用于白天Re的估算。本文利用u*>0.2 m/s条件下夜间生态系统碳通量与土壤温度的关系(图1)建立回归方程如下:
Re=0.0235e0.1444TsR2=0.74
(5)
式中,Ts为5 cm深度土壤温度。
1.3.3 冠层导度的计算
冠层导度(gc)通过Pennman-Monteith方法反推计算[20]:
(6)
式中,Rn为净辐射(W/m2);γ为干湿常数(0.065 kPa/K);λ为蒸发潜热(2.25 MJ/kg);Δ为饱和差随温度变化的斜率(kPa/K);ra为大气边界层空气动力学阻抗(s/m);ρ为空气密度(kg/m3);cp是空气定压比热(1013 J kg-1K-1);VPD为饱和差(kPa)。其中,Rn和ET直接通过涡度相关观测获得,ra根据Monin-Obukhov相似理论中性条件假设计算:
(7)
式中,z为观测高度3 m;d为零平面位移;z0为空气动力学粗糙度;k为卡曼常数0.41;u*为摩擦速度。其中u*通过涡度相关观测直接获得,d和z0分别取值0.75h和0.1h[21],h为0.65 m。
1.4 数据处理
原始采样数据行进了WPL校正和3次坐标轴旋转修正,夜间风速较小易形成平流或泄流现象,以夜间风速u*> 0.2 m/s的通量值为有效数据,同时剔除受降雨、凝结水等气象因子影响造成的异常数据,并用能量平衡闭合度EBR来评价涡度相关系统的数据质量,研究期间该涡度相关系统能量平衡闭合度为0.80,处于国际中上水平[22]。计算日均值时,对时间段低于2 h的异常值根据前后数据进行线性插补,高于2 h的异常值不计其平均值,直接将该日排除。
在计算水分利用效率日变化时选取9:00—16:00之间的碳水通量值进行分析;在计算和分析季节变化时,选取9:00—16:00的平均值作为该天的代表值,用所得平均值计算该天的水分利用效率平均值。
2 结果分析
2.1 水分利用效率影响因子的季节变化
2.1.1 环境因子的季节变化
研究区生长季净辐射、气温和饱和差9:00—16:00均值的季节变化均呈单峰曲线形式(图2),净辐射和气温最大值均出现在7月中旬,分别为558.38 W/m2和30.2 ℃;饱和差最大值出现时间稍有滞后,出现在7月末,为2.61 kPa。生长季净辐射、气温和饱和差日均值分别为382.79 W/m2、21.07 ℃和1.34 kPa。
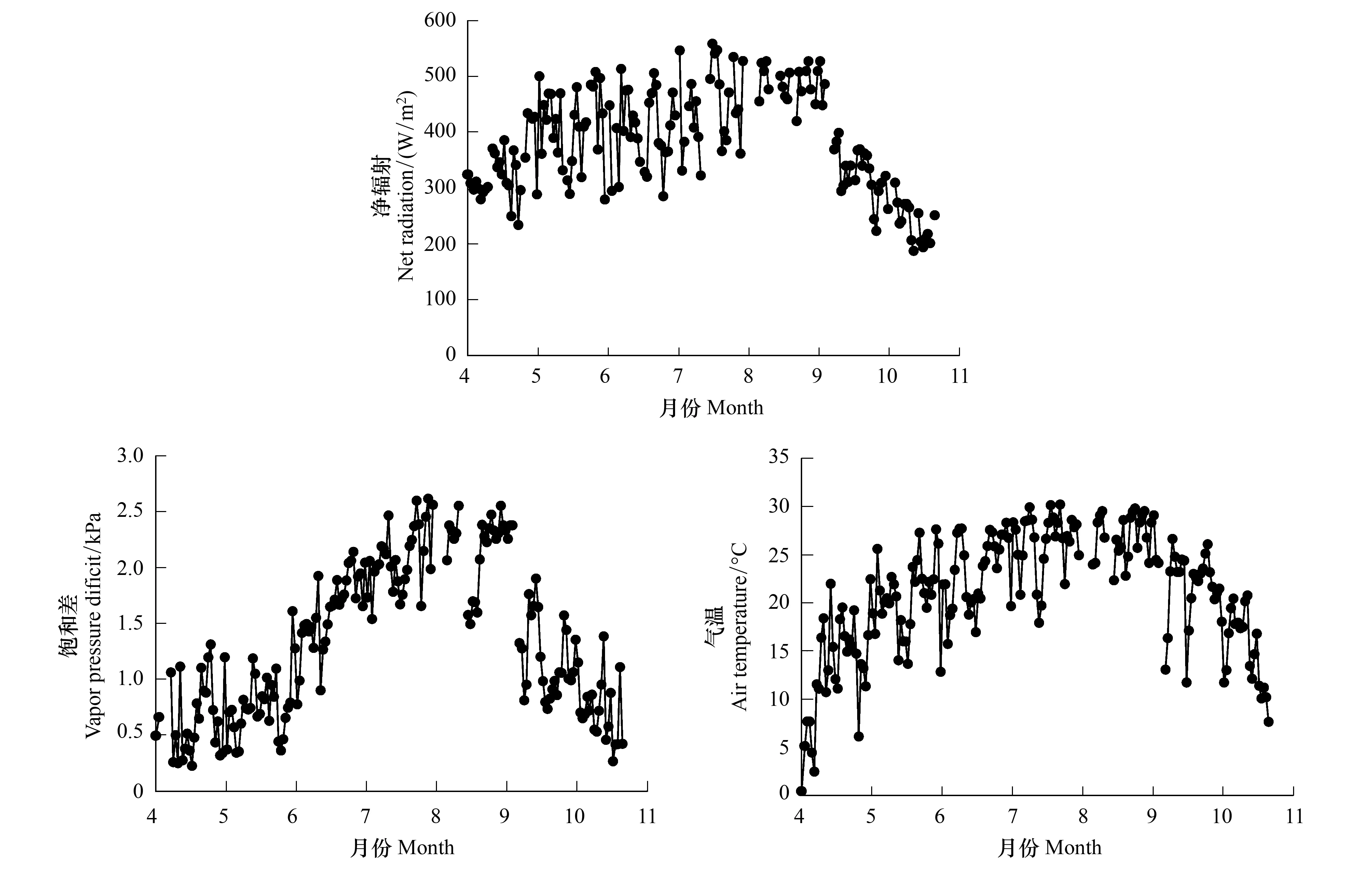
图2 科尔沁草甸净辐射、饱和差和气温的季节变化Fig.2 Seasonal variation of net radiation, vapor pressure deficit and air temperature over Horqin Meadow
2.1.2 植被叶面积指数和冠层导度季节变化
生长季LAI具有明显的季节变化,生长期内的5—6月中旬LAI迅速增加,随后增速下降,8月中旬达到最大值4.97,以后缓慢下降(图3)。生长季平均LAI为3.48,高于其他类型的草甸植被类型[23- 24]。
冠层导度gc在生长季内基本呈单峰曲线形式(图3)。4月末伴随着植被的萌发gc开始出现,之后随着植被的生长逐渐增大,7月中旬达最大,为0.0105 m/s,而后缓慢下降。生长季平均gc为0.0046 m/s。
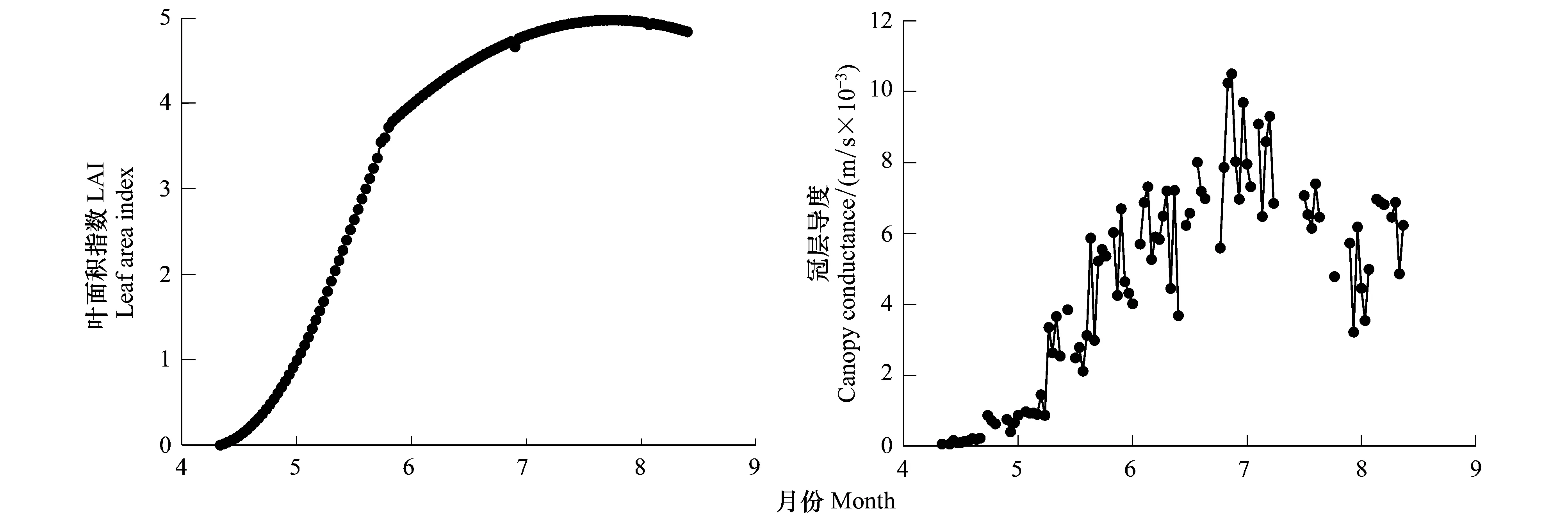
图3 科尔沁草甸植物叶面积指数和冠层导度的季节变化Fig.3 Seasonal variation of LAI and gc over Horqin Meadow
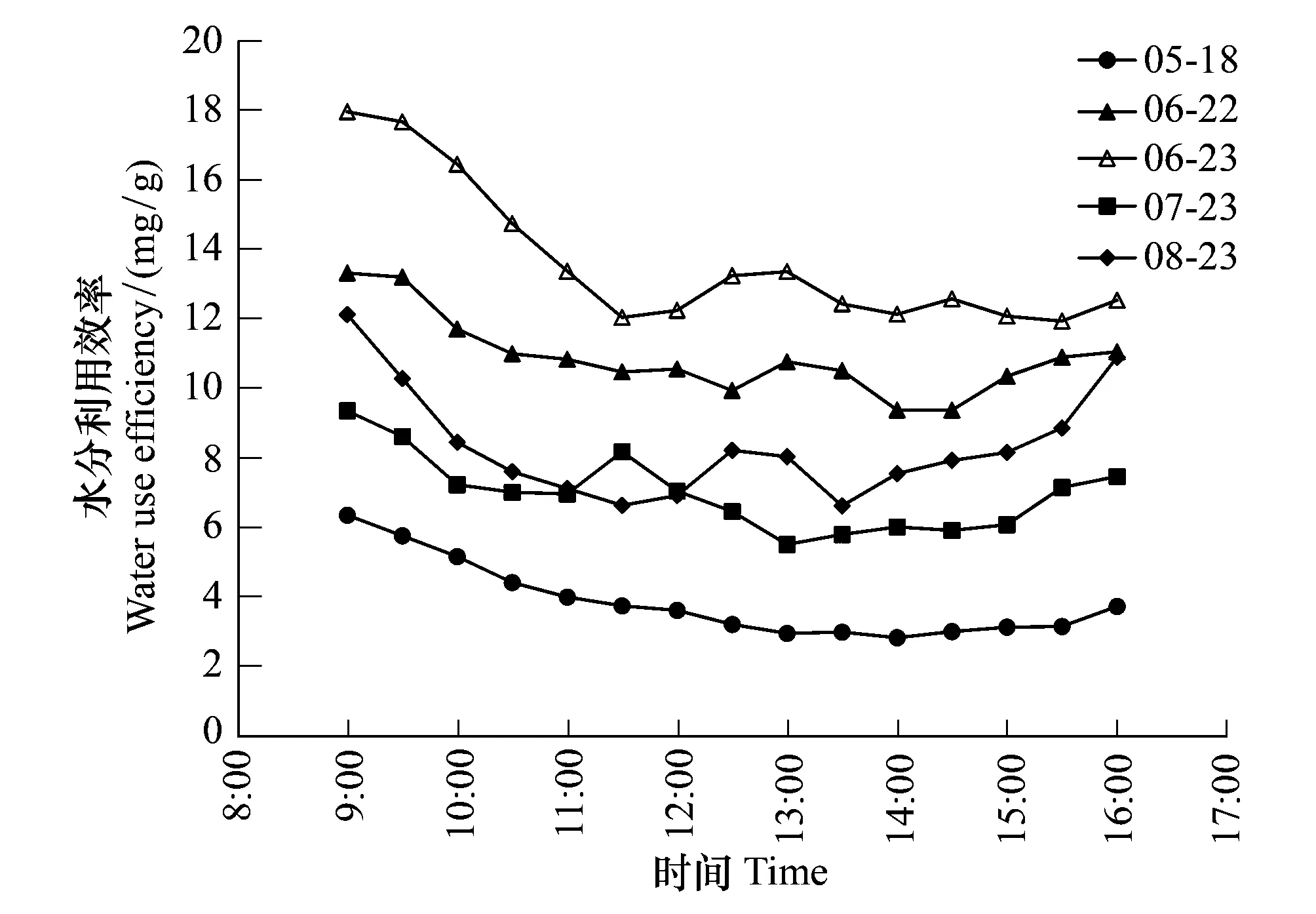
图4 科尔沁草甸总初级生产力水分利用效率的日变化Fig.4 Diurnal variation of WUEGPP over Horqin Meadow
2.2 水分利用效率日变化特征
该生态系统WUEGPP存在明显的日变化特征,尽管存在小范围波动,但在日变化过程中从9:00—16:00基本呈现出下降—稳定—上升的变化规律(图4)。WUEGPP日出后不久达到最大值,而后逐渐降低,到12:00前后趋于稳定,15:00以后又开始小幅上升。
天气变化对WUEGPP具有显著影响,以6月22日(晴天)和6月23日(阴天)为例,两天净辐射值分别为11.83和7.93 MJ/m2。虽然两天WUEGPP日变化规律基本相同,但因晴天和阴天得到的光照、辐射以及所造成的其他环境要素变化不同,导致大小不同,WUEGPP日均值分别为10.89和13.65 mg/g,阴天条件下WUEGPP高于晴天,与之前的研究结果[25- 26]一致。
从生长季4个月典型晴天5月18日、6月22日、7月23日和8月23日的比较来看,生长中期WUEGPP要高于生长初期和末期,日均值在6月22日最大,为10.89 mg/g,5月18日最小,为3.86 mg/g,7月23日和8月23日分别为6.98和8.55 mg/g。
2.3 总初级生产力、蒸散量和水分利用效率季节变化特征
GPP日均值季节变化呈夏季高、春秋低的形式(图5),与LAI增长速度趋势相同。4月末随着植被生长,GPP开始出现而后迅速增大,6月末出现最大值1.49 mg m-2s-1,高于其他类型草地生态系统[27- 28],之后逐渐降低,9月1日前后由于人工割草,GPP显著下降,之后维持较低数值。整个生长季GPP平均值为0.57 mg m-2s-1。GPP最大值并没有出现在净辐射最强烈的7月中旬,这主要是由于7月辐射较大,温度较高,午间气孔关闭出现“午休”现象,从而影响了光合作用,这是植被实现水分利用最优的一种适应性调节[29]。
ET受净辐射的影响,季节变化规律与净辐射相似(图5),7月中旬出现最大值0.16 g m-2s-1,生长季ET平均值为0.08 g m-2s-1,潜热通量占净辐射的61%。但生长旺季(6月到8月)ET始终保持较高数值,变化趋势不明显,这主要是由于该区降水丰富,且生长旺季潜热的能量来源(净辐射)和蒸散驱动力(饱和差)变化较小造成。
WUEGPP季节变化呈与GPP相似的形式(图5)。4月末随着植被生长开始迅速增大,到6月末出现最大值13.62 mg/g,与GPP类似,WUEGPP在7月同样出现了连续多天的低值情况,生长季平均WUEGPP为5.97 mg/g。
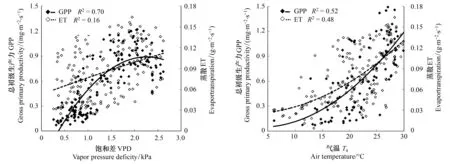
图6 科尔沁草甸总初级生产力和蒸散量与饱和差和气温的关系Fig.6 Relationship between GPP, ET and Ta, VPD over Horqin Meadow
2.4 水分利用效率的影响因子及其影响规律
2.4.1 环境因子对水分利用效率的影响
研究区生长季VPD与GPP呈显著的二次曲线关系(图6),生长初期GPP随饱和差的增大而增大,当VPD达到2.2 kPa左右时,GPP受到抑制不再上升,甚至出现下降趋势。其原因是当空气干燥时,气孔因叶片与大气间的水汽压差较高而关闭,抑制了光合作用[30]。ET随着VPD的增大而有略微增大,但两者间的相关性较差(图6)。综合VPD对GPP和ET的影响可以看出,VPD对WUEGPP的影响主要是通过影响GPP的大小而实现,因此WUEGPP与VPD间也呈二次曲线关系(图7),且随VPD的变化趋势与GPP相似,开始随着VPD的增大,WUEGPP迅速增大,到2.0 kPa时,不再增大,反而有略微下降。
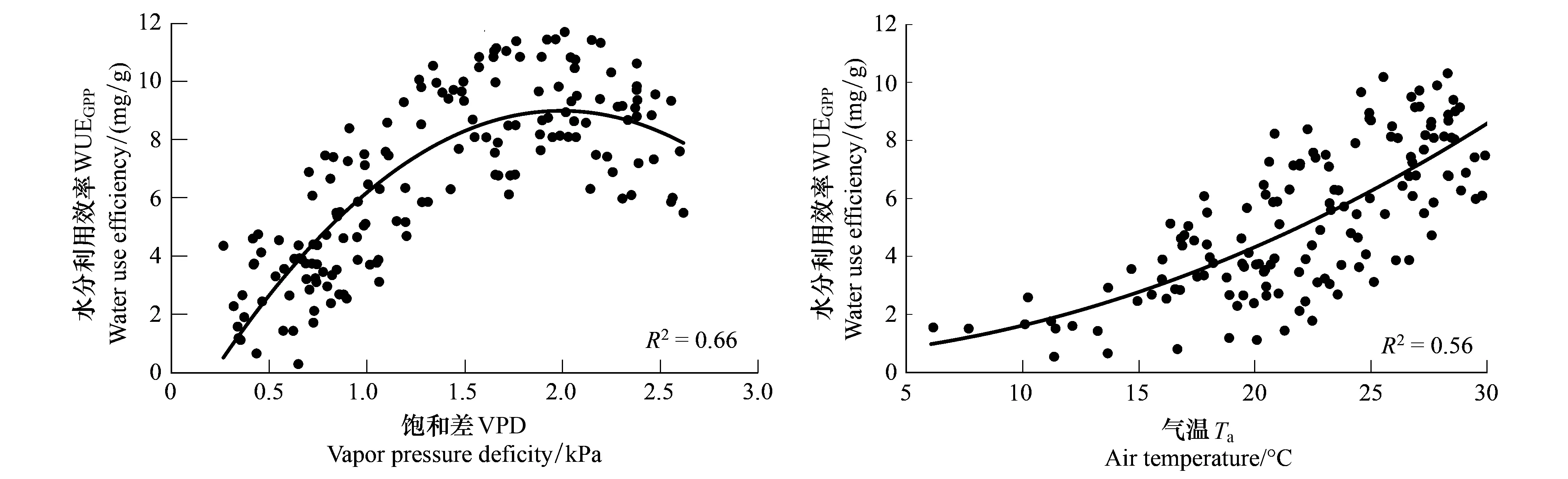
图7 科尔沁草甸水分利用效率与饱和差和温度的关系Fig.7 Relationship between WUEGPP and VPD, Ta over Horqin Meadow
研究表明一定范围内,生态系统光合和蒸散作用都会随着温度的升高而增大[31],但二者增加速率不同,由二者决定的生态生态系统WUEGPP的值也不同。科尔沁草甸草地GPP和ET都随气温的升高而增大,均与气温呈二次曲线关系(图6)。GPP和ET在高温时并没有显示下降的趋势,说明研究区生长旺季的土壤水分条件较好,未达到严重亏缺状态。但二者对气温升高的敏感程度不同,GPP随气温升高的速率大于蒸散,导致WUEGPP随着气温的升高而增大,也呈二次曲线形式(图7)。
由于VPD是温度的函数,通常气温的升高都要伴随VPD的增加,因此将两种因素的效应进行分离是非常困难的,同时VPD表征着空气水分状况,对土壤蒸发也具有调节作用。生态系统环境因子间相互作用,共同控制生态系统的碳水通量过程,其内在机制十分复杂,还需要深入研究。
2.4.2 叶面积指数和冠层导度对水分利用效率的影响
作为植被生长状况的特征参数之一,叶面积指数一方面直接影响生态系统总初级生产力[32]和地气间水汽交换的阻力大小,另一方面影响植被冠层内部的光分布状况[33],从而间接对光合和蒸散产生影响。胡中民[34]对中国典型草地生态系统的研究表明,LAI不仅是这些生态系统WUEGPP季节变化的控制因子,也是不同草地类型间WUEGPP差异的主要来源。
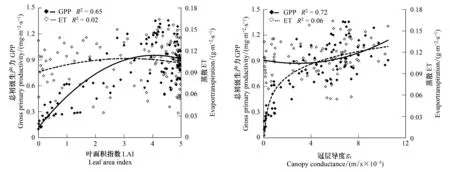
图8 科尔沁草甸总初级生产力和蒸散量与叶面积指数和冠层导度的关系Fig.8 Relationship between GPP, ET and LAI, gc over Horqin Meadow
研究区生长季GPP与LAI呈二次曲线关系(图8),当LAI<3.8时,随着LAI的增大GPP也增大,当LAI超过3.8时,GPP基本保持较高的数值,甚至有略微下降,可能是由于7月份LAI到达最大值时午间气孔关闭所致。ET与LAI的相关性较差(图8),随着LAI的变化ET变化不明显,主要是由于整个生长季降水充足,下垫面植被生长茂盛,蒸腾强烈造成。GPP和ET与LAI的上述关系决定了WUEGPP与LAI的相关性以及WUEGPP随LAI变化的趋势。由于ET与LAI的相关性较差,GPP与LAI相关关系决定了WUEGPP与LAI的相关关系,即WUEGPP与LAI呈二次曲线关系(图9)。与GPP随LAI的变化趋势相似,当LAI<4.2时,随着LAI的增加,WUEGPP增大,但当LAI超过阈值时,WUEGPP基本保持较高的数值,后略有下降。说明生长季随着植被的生长,固定相同质量的二氧化碳所需要的水分减少,主要是因为植被生长茂密,阻挡了太阳辐射到达地面,减少了无效的土壤蒸发,而随着植被的生长,部分叶片衰老,光合能力下降[35],消耗同样水分的同时固定的二氧化碳减少,因此当LAI超过一定值时WUEGPP表现下降。
作为植物生态系统气孔交换的整体指标,冠层导度是影响下垫面与大气间水汽传输阻力大小的重要因素之一,同时由于植被气孔对二氧化碳的阻抗是水汽阻抗的1.6倍,可以用水汽阻抗来计算植被气孔对二氧化碳的阻抗,因此冠层导度可以同时表征生态系统与大气间碳水交换的阻力大小从而对WUEGPP产生影响。
生长季GPP与gc呈对数曲线关系(图8),随着gc的增加GPP也增大,当gc<0.0015 m/s时尤为明显,gc超过0.0015 m/s后,GPP随gc增加的速度减慢,说明植被与大气间二氧化碳的交换在低gc条件下,对gc的变化反应敏感,而当gc超过一定值时,由于植被本身光合作用达到上限,光合作用对gc的变化反应能力下降,gc变成影响光合作用的次要因素。ET与gc的相关性同样较差(图8)。gc对植物光合作用和蒸腾作用的影响程度是不同的,比较GPP和ET对gc的响应模式可以看出,WUEGPP对gc的响应主要由GPP对gc的响应模式决定,WUEGPP随着gc的增加而增加,呈对数曲线关系(图9)。同样,当gc<0.0015 m/s时,WUEGPP随着gc的增加迅速增加,超过这一阈值随gc增加的速度减慢,gc变为影响WUEGPP的次要因素。
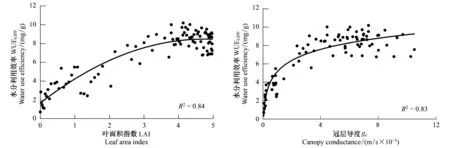
图9 科尔沁草甸水分利用效率与叶面积指数和冠层导度的关系Fig.9 Relationship between WUEGPP and LAI, gc over Horqin Meadow
3 结论与讨论
3.1 科尔沁草甸草地水分利用效率特征
本文利用科尔沁温带草甸碳水通量观测数据,对该生态系统WUEGPP的变化规律及其对环境和生理因子的响应规律进行了分析。结果表明:WUEGPP日变化呈下降-稳定-上升的变化趋势,最大值出现在日出后,阴天条件下WUEGPP明显高于晴天,生长中期WUEGPP高于生长初期和末期。GPP、ET和WUEGPP季节变化均呈夏季高、春秋低的形式,生长季平均值分别为0.57 mg m-2s-1、0.08 g m-2s-1和5.97 mg/g,最大值分别为1.49 mg m-2s-1、0.16 g m-2s-1和13.62 mg/g;GPP与VPD、Ta和LAI均呈二次曲线关系,与gc呈对数曲线关系;ET与Ta呈二次曲线关系,与VPD、LAI和gc相关性均不显著。WUEGPP与VPD、Ta和LAI均呈二次曲线关系,与gc呈对数曲线关系,VPD、gc和LAI分别为2.0 kPa、0.0015 m/s和4.2是控制WUEGPP增加的阈值,当超过阈值后,WUEGPP会保持不变或下降。
3.2 不同定义下的WUE的比较
近年来,涡度相关技术的不断发展为生态系统水分利用效率的研究提供了一种新的技术方法,但不同学者对生态系统水分利用效率的定义略有不同。一些学者将其定义为总初级生态系统生产力GPP与蒸散量的比(WUEGPP),而有些学者则建议用净生态系统生产力NEP与蒸散量的比(WUENEP)或用净初级生产力NPP与蒸散量的比(WUENPP)来定义,这样限制了生态系统间的对比分析[1]。为了综合反映研究区的碳水耦合特征,方便与其他研究区域进行比较,本文利用后两种方法对该研究区域的水分利用效率进行了同步计算,其中NEP直接通过涡度相关观测获得,NPP跟据涡度相关观测与土壤呼吸相加获得,其中土壤呼吸根据陈妮娜等[36]建立的与地表5 cm处土壤温度的回归方程计算。
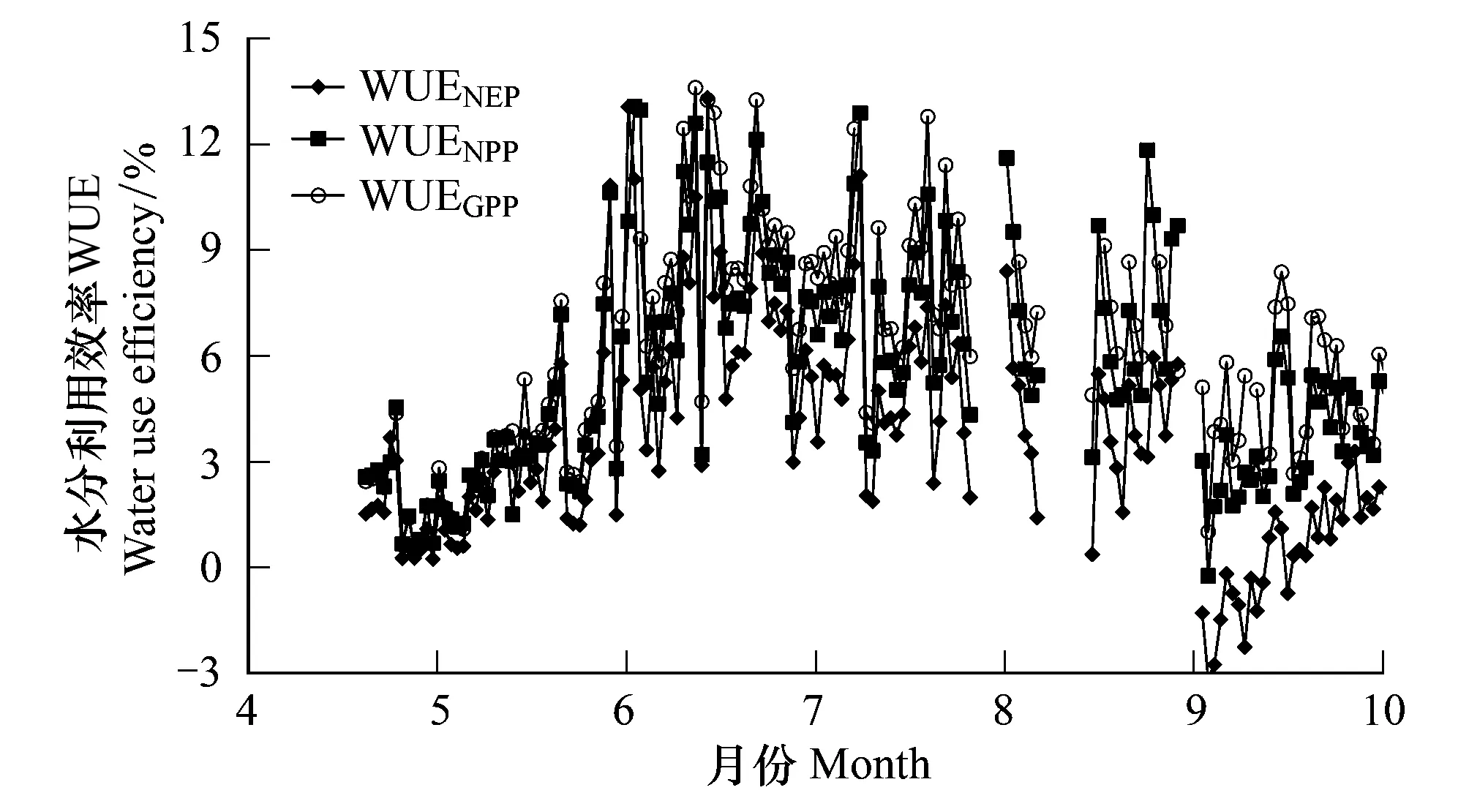
图10 不同定义下水分利用效率的季节变化 Fig.10 Seasonal variation of water use efficiency with different definitionsWUEGPP:总初级生产力水分利用效率(GPP/ET);WUENEP:净生态系统生产力水分利用效率(NEP/ET):WUENPP:净初级生产力水分利用效率(GPP/ET)
3种方法得到的水分利用效率季节变化趋势基本相同(图10),但由于生产力计算方法的差异,计算得到的水分利用效率大小不同,整个生长季WUENPP和WUENEP均值大小分别为5.21和3.49 mg/g,最大值分别为12.62和10.51 mg/g。
3.3 不同生态系统的WUE大小及变异特征
关于不同生态系统水分利用效率的比较已有很多研究。国际上Law等[37]基于全球通量观测网络(FLUXNET)涡度相关观测数据, 对不同生态系统月平均WUE进行总结发现,森林生态系统的WUE高于草地和冻原。国内胡中民等[1]对不同生态系统的WUE进行比较发现常绿针叶林的WUE高于落叶阔叶林,水热条件较好的草地生态系统WUE高于水热条件较差的草地生态系统。在上述研究基础上,结合草甸草地的研究结果表1对不同气候条件不同生态系统的WUE进行了进一步总结,发现森林生态系统的WUE高于草地,温带湿润草地和草甸草地高于温带荒漠草地和高寒草地,农田WUE介于森林和草地之间,接近于温带湿润草地和草甸草地。各生长季生态系统WUE的大致变异范围分别为: 常绿针叶林和针阔混交林在10 mg/g以上,人工混交林和常绿阔叶林在6—10 mg/g,草地在0.4—6 mg/g之间,农田在5.4 mg/g左右。科尔沁草甸草地生长季WUE平均为5.97 mg/g,与中国东北样带其他生态系统相比,低于森林生态系统(长白山针阔混交林),但高于荒漠草地生态系统(内蒙古锡林郭勒温带荒漠草地),与中国东北样带的生态系统多样性梯度研究结论一致[38]。
[1] 胡中民, 于贵瑞, 王秋凤, 赵风华. 生态系统水分利用效率研究进展. 生态学报, 2009, 29(3): 1498- 1507.
[2] Neumann H H, Den Hartog G, King K M, Chipanshi A C. Carbon dioxide fluxes over a raised open bog at the Kinosheo Lake tower site during the Northern Wetlands Study (NOWES). Journal of Geophysical Research, 1994, 99(D1): 1529- 1538.
[3] 崔骁勇, 杜占池, 王艳芬. 内蒙古半干旱草原区沙地植物群落光合特征的动态研究. 植物生态学报, 2000, 24(5): 541- 546.
[4] Xu L K, Hsiao T C. Predicted versus measured photosynthetic water-use efficiency of crop stands under dynamically changing field environments. Journal of Experimental Botany, 2004, 55(407): 2395- 2411.
[5] 刘文兆. 作物生产、水分消耗与水分利用效率间的动态联系. 自然资源学报, 1998, 13(1): 23- 27.
[6] 高聚林, 赵涛, 王志刚, 郭改玲, 范磊. 高丹草水分利用效率与叶片生理特性的关系. 作物学报, 2007, 33(3): 455- 460.
[7] Picotte J J, Rosenthal D M, Rhode J M, Cruzan M B. Plastic responses to temporal variation in moisture availability: consequences for water use efficiency and plant performance. Oecologia, 2007, 153(4): 821- 832.
[8] Ponton S, Flanagan L B, Alstad K P, Johnson B G, Morgenstern K, Kljun N, Black T A, Barr A G. Comparison of ecosystem water-use efficiency among Douglas-fir forest, aspen forest and grassland using eddy covariance and carbon isotope techniques. Global Change Biology, 2006, 12(2): 294- 310.
[9] Baldocchi D, Falge E, Gu L H, Olson R, Hollinger D, Running S, Anthoni P, Bernhofer C, Davis K, Evans R, Fuentes J, Goldstein A, Katul G, Law B, Lee X H, Malhi Y, Meyers T, Munger W, Oechel W, Paw K T, Pilegaard K, Schmid H P, Valentini R, Verma S, Vesala T, Wilson K, Wofsy S. FLUXNET: a new tool to study the temporal and spatial variability of ecosystem-scale carbon dioxide, water vapor, and energy flux densities. Bulletin of the American Meteorological Society, 2001, 82(11): 2415- 2434.
[10] 朱治林, 孙晓敏, 张仁华, 苏红波, 唐新斋. 作物群体 CO2通量和水分利用效率的快速测定. 应用生态学报, 2004, 15(9): 1684- 1686.
[11] Parton W J, Schimel D S, Cole C V, Ojima D S. Analysis of factors controlling soil organic matter levels in Great Plains grasslands. Soil Science Society of America Journal, 1987, 51(5): 1173- 1179.
[12] 陈拓, 冯虎元, 徐世建, 强维亚, 安黎哲. 荒漠植物叶片碳同位素组成及其水分利用效率. 中国沙漠, 2002, 22(3): 288- 291.
[13] 王静, 程积民, 万惠娥, 方锋. 黄土高原芨芨草土壤水分特征及水分利用效率研究. 干旱气象, 2004, 22(4): 51- 55.
[14] Martin B, Thorstenson Y R. Stable carbon isotope composition (δ13C), water use efficiency, and biomass productivity of Lycopersicon esculentum,Lycopersiconpennellii, and the F1hybrid. Plant Physiology, 1988, 88(1): 213- 217.
[15] Donovan L A, Dudley S A, Rosenthal D M, Ludwig F. Phenotypic selection on leaf water use efficiency and related ecophysiological traits for natural populations of desert sunflowers. Oecologia, 2007, 152(1): 13- 25.
[16] 于贵瑞, 伏玉玲, 孙晓敏, 温学发, 张雷明. 中国陆地生态系统通量观测研究网络 (ChinaFLUX) 的研究进展及其发展思路. 中国科学D辑, 2006, 36(S1): 1- 21.
[17] 王文, 王劲廷. 1951- 2009 年中国地表湿润状况变化趋势研究. 气象与环境学报, 2011, 27(5): 1- 6.
[18] 周广胜, 王玉辉, 白莉萍, 许振柱, 石瑞香, 周莉, 袁文平. 陆地生态系统与全球变化相互作的研究进展. 气象学报, 2004, 62(5): 692- 707.
[19] Jonckheere I, Fleck S, Nackaerts K, Muys B, Coppin P, Weiss M, Baret F. Review of methods for in situ leaf area index determination: Part I. Theories, sensors and hemispherical photography. Agricultural and Forest Meteorology, 2004, 121(1): 19- 35.
[20] 李峥, 牛丽华, 袁凤辉, 关德新, 王安志, 金昌杰, 吴家兵. 辽西农林复合系统中杨树冠层导度特征. 应用生态学报, 2012, 23(11): 2975- 2982.
[21] Thom A S. Momentum, mass and heat exchange of vegetation. Quarterly Journal of the Royal Meteorological Society, 2006, 98(415): 124- 134.
[22] Wilson K, Goldstein A, Falge E, Aubinet M, Baldocchi D, Berbigier P, Bernhofer C, Ceulemans R, Dolman H, Field C, Grelle A, Ibrom A, Law B E, Kowalski A, Meyers T, Moncrieff J, Monson R, Oechel W, Tenhunen J, Valentini R, Verma S. Energy balance closure at FLUXNET sites. Agricultural and Forest Meteorology, 2002, 113(1/4): 223- 243.
[23] 赵亮, 徐世晓, 李英年, 唐艳鸿, 赵新全, 古松, 杜明远, 于贵瑞. 青藏高原矮嵩草草甸和金露梅灌丛草甸CO2通量变化与环境因子的关系. 西北植物学报, 2006, 26(1): 133- 142.
[24] 孙建文, 李英年, 宋成刚, 王建雷, 张法伟, 王勤学. 高寒矮嵩草草甸地上生物量和叶面积指数的季节动态模拟. 中国农业气象, 2010, 31(2): 230- 234.
[25] Alarcón J J, Ortuo M F, Nicolás E, Navarro A, Torrecillas A. Improving water-use efficiency of young lemon trees by shading with aluminised-plastic nets. Agricultural Water Management, 2006, 82(3): 387- 398.
[26] 高照全, 李天红, 冯社章. 苹果叶片瞬时水分利用效率动态模拟. 干旱地区农业研究, 2011, 29(3): 101- 106.
[27] 闫巍, 张宪洲, 石培礼, 杨振林, 何永涛, 徐玲玲. 青藏高原高寒草甸生态系统 CO2通量及其水分利用效率特征. 自然资源学报, 2006, 21(5): 757- 767.
[28] 张法伟, 刘安花, 李英年, 赵亮, 王勤学, 杜明远. 青藏高原高寒湿地生态系统 CO2通量. 生态学报, 2008, 28(2): 453- 462.
[29] 郑有飞, 万长建, 颜景义, 廖志鸿. 小麦的水分利用效率及其最优化问题. 中国农业气象, 1997, 18(4): 13- 17.
[30] Maroco J P, Pereira J S, Chaves M M. Stomatal responses to leaf-to-air vapour pressure deficit in Sahelian species. Functional Plant Biology, 1997, 24(3): 381- 387.
[31] 蒋高明, 何维明. 毛乌素沙地若干植物光合作用、蒸腾作用和水分利用效率种间及生境间差异. 植物学报, 1999, 41(10): 1114- 1124.
[32] Gower S T, Krankina O, Olson R J, Apps M, Linder S, Wang C. Net primary production and carbon allocation patterns of boreal forest ecosystems. Ecological Applications, 2001, 11(5): 1395- 1411.
[33] Duncan W G. Leaf angles, leaf area, and canopy photosynthesis. Crop Science, 1971, 11(4): 482- 485.
[34] 胡中民. 中国草地样带典型生态系统水分利用效率的时空变异特征研究. 北京: 中国科学院地理科学与资源环境研究所, 2008.
[35] 杜占池, 杨宗贵. 羊草不同叶龄叶片光-光合特性的初步研究. 植物学报, 1988, 30(2): 196- 206.
[36] 陈妮娜, 关德新, 金昌杰, 袁凤辉, 杨弘. 科尔沁草甸草地土壤呼吸特征. 中国草地学报, 2011, 33(5): 82- 87.
[37] Law B E, Falge E, Gu L, Baldocchi D D, Bakwin P, Berbigier P, Davis K, Dolman A J, Falk M, Fuentes J D, Goldsein A, Granier A, Grelle A, Hollinger D, Janssens I, Jarvis P, Jensen N O, Katul G, Mahli Y, Matteucci G, Meyers T, Monson R, Munger W, Oechel W, Olson R, Pilegaard K, Pawu U K T, Thorgeirsson H, Valentini R, Verma S, Vesala T, Wilson K, Wofsy S. Environmental controls over carbon dioxide and water vapor exchange of terrestrial vegetation. Agricultural and Forest Meteorology, 2002, 113(1): 97- 120.
[38] 唐海萍, 张新时. 中国东北样带的生态系统多样性梯度研究. 第四纪研究, 1999, (5): 479- 479.
Water use efficiency and its influential factor over Horqin Meadow
LI Huidong1,2, GUAN Dexin1,*, YUAN Fenghui1, WANG Anzhi1, JIN Changjie1, WU Jiabing1, LI Zheng1,2, JING Yanli1,2
1StateKeyLaboratoryofForestandSoilEcology,InstituteofAppliedEcology,ChineseAcademyofSciences,Shenyang110016,China2UniversityofChineseAcademyofSciences,Beijing100049,China
At ecosystem level, water use efficiency (WUE, defined as the net carbon uptake per amount of water lost from the ecosystem) reflects the coupling of carbon and water cycles. Continuous observation of carbon and water fluxes was made over Horqin Meadow with eddy covariance system in growing season of 2012. This paper calculated the water use efficiency, WUEGPP, defined as the ratio of gross primary productivity, GPP, to evapotranspiration, ET. Diurnal and seasonal variation of WUEGPPand its response to environmental and physiological factors were analyzed. The results showed that the diurnal variation of WUEGPPpresented a trend of decreasing-stable-increasing, and the appearance of maximum was one or two hours later after sunrise. WUEGPPon cloudy day was greater than that on sunny day, and the WUEGPPat the mid of the growing season was greater than that in the beginning and at the end of the growing season. Seasonal variation of GPP and ET both presented a trend of higher in summer and lower in spring and autumn. Their maximum values appeared at the end of June and in mid-July, and were 1.49 mg m-2s-1and 0.16 g m-2s1, respectively. Because of harvest, GPP and ET decreased rapidly around September 1st. The seasonal averaged GPP and ET were 0.57 mg m-2s-1and 0.08 g m-2s1respectively. Seasonal variation of WUEGPPalso presented a trend of higher in summer and lower in spring and autumn, and the maximum value (13.62 mg/g) appeared at the end of June. The seasonal averaged WUEGPPwas 5.97 mg/g, which was higher than that of forest ecosystem and lower than that of desert grassland ecosystem. GPP presented a quadratic relationship with vapor pressure deficit, air temperature and leaf area index, respectively, and a logarithmic relationship with canopy conductance. ET presented a quadratic relationship with air temperature, while its correlation with vapor pressure deficit, leaf area index and canopy conductance was non-significant. WUEGPPpresented a quadratic relationship with vapor pressure deficit, air temperature and leaf area index, respectively, and a logarithmic relationship with canopy conductance. Vapor pressure deficit equal to 2.0 kPa, leaf area index equal to 4.2 and canopy conductance equal to 0.002 m/s were the critical points that limit the increase of WUEGPP. When vapor pressure deficit was less than 2.0 kPa, WUEGPPenlarged with the increase of vapor pressure deficit, and when it exceeded 2.0 kPa, the situation was opposite. With the increase of leaf area index, WUEGPPenlarged gradually under low leaf area index, but when it exceeded 4.2, WUEGPPdid not increase and tended towards stability with the continual increase of leaf area index. With the increase of canopy conductance, WUEGPPenlarged rapidly at begin, but slowly with the continual increase of canopy conductance when it exceeded the critical point. Water use efficiency expressed by other productive indexes was also estimated, including WUENEP(defined as the ratio of net ecosystem production, NEP, to ET) and WUENPP(defined as the ratio of net primary production, NPP, to ET). The seasonal variation of WUENEPand WUENPPpresented a similar trend with WUEGPP,and their averages were 3.47 and 5.47 mg/g respectively. Comparison of WUE in different ecosystems found that leaf area index was the dominant factor of WUE magnitude because that leaf area index was a comprehensive reflection of air temperature and precipitation, while air temperature or precipitation alone can not determine the WUE of an ecosystem.
water use efficiency (WUE); gross primary productivity (GPP); evapotranspiration(ET); influential factor
国家重点基础研究计划(973计划)(2013CB429902); 国家自然科学基金(40875069)
2013- 03- 31;
日期:2014- 03- 25
10.5846/stxb201303310566
*通讯作者Corresponding author.E-mail: dxguan@iae.ac.cn
李辉东,关德新,袁凤辉,王安志,金昌杰, 吴家兵,李峥,井艳丽.科尔沁草甸生态系统水分利用效率及影响因素.生态学报,2015,35(2):478- 488.
Li H D, Guan D X, Yuan F H, Wang A Z, Jin C J, Wu J B, Li Z, Jing Y L.Water use efficiency and its influential factor over Horqin Meadow.Acta Ecologica Sinica,2015,35(2):478- 488.
猜你喜欢
儿童故事画报·自然探秘(2024年5期)2024-05-22 22:26:58
疯狂英语·新读写(2023年4期)2023-05-10 10:44:22
青海草业(2022年2期)2022-07-23 09:34:58
草原歌声(2020年4期)2020-03-29 06:40:18
中国海洋大学学报(自然科学版)(2019年11期)2019-10-12 06:04:46
活力(2019年21期)2019-04-01 12:17:10
热带气象学报(2019年6期)2019-03-02 16:42:38
草原歌声(2018年3期)2018-12-03 08:14:46
草原歌声(2018年2期)2018-12-03 08:14:08
科技资讯(2018年30期)2018-03-11 09:30:58
