
撑×绿杂交竹叶部细菌多样性1)
2015-03-10 06:55彭艳毛毳朱天辉刘洋
东北林业大学学报 2015年6期
彭艳 毛毳 朱天辉 刘洋
(四川农业大学,雅安,625014)
撑×绿杂交竹叶部细菌多样性1)
彭艳 毛毳 朱天辉 刘洋
(四川农业大学,雅安,625014)
通过固定标准地分季节采样,采用稀释平板分离,借助分子生物学方法研究了撑×绿杂交竹叶片附生细菌和内生细菌数量及种类的季节性动态变化、群落组成及叶部细菌和微环境的关系。在此基础上,将以上4个指标从病叶和健康叶上着重进行了对比分析,发现杂交竹病叶附生和内生细菌数量的年平均值都大于健康叶;夏、秋两季细菌的类群最多,其中以Bacilluscereus和Lysinibacillusfusiformis的分离频率较高;无论是健康叶还是病叶,其附生和内生细菌的物种丰富度随季节变化较大,而且季节变化对内生细菌物种的丰富度影响显著;叶片生理指标表明健康叶附生细菌的数量随pH值的升高而增加,健康叶内生细菌和病叶附生细菌的数量与蛋白质质量分数相关性显著。
撑×绿杂交竹;健康叶;病叶;细菌多样性;生物防治
Based on sampling on permanent sample plots in different seasons, we studied the seasonal dynamic changes of quantity and species of epiphytic bacteria and endophitic bacteria, bacteria community composition and microenvironment of leaves ofBambusapervariabilis×Dendrocalamopsisdaiiby plate dilution method and molecular biological method. We also comparatively analyzed four indexes between healthy leaves and diseased leaves. The quantity of epiphytic bacteria and endophytic bacteria of diseased leaves were more than healthy leaves in all the seasons. Bacteria species were most in summer and autumn, and isolation rates ofBacilluscereusandLysinibacillusfusiformiswere higher than that of the others’. For healthy leaves and diseased leaves, the species richness of epiphyticbacteria and endophytic bacteria changed greatly with seasons, and seasonal change had significant influence on species richness of endophytic bacteria. Physiological indexes showed that quantity of epiphytic bacteria of healthy leaves increased with pH, and the correlations between quantity of endophytic bacteria of healthy leaves and epiphytic bacteria of diseased leaves and content of protein were significant.
一直以来,人们对植物根际细菌做过深入的研究[1-2],而作为叶部微生物的重要组成成分,植物叶部的附生细菌和内生细菌的研究却被人们忽视。自从Last[3]和Ruinen[4]分别对禾谷类作物叶面和热带森林叶表微生物生态进行系统研究以后,才极大地促进了人们对叶部微生物的研究。有研究表明叶部附生细菌在其生长发育过程中会产生具有拮抗性或竞争性的一种或几种代谢产物,从而达到抑制植物病原菌的效果[5],此外,其在促进植物生长、降解环境污染、生物传感监测等方面也有重要作用[6-7]。植物内生细菌同样具有多方面的生物学作用,如促生、防病、内生固氮[8-10],同时研究发现,植物内生细菌还是良好的外源基因载体,为防病、杀虫及固氮等植物外源基因导入植物体内提供了另一种方法和途径[11]。
撑×绿杂交竹是具有良好发展前途的优良经济竹种。近年,来雅安地区的撑×绿杂交竹屡屡遭受病害的侵袭,产量不断减少,给当地的林业经济造成了严重的损害。为了更好地防治撑×绿杂交竹叶部病害,本文通过对撑×绿杂交竹病、健康叶上细菌多样性的对比分析,从微生态的角度研究了细菌数量及种群的变化,以期为杂交竹枝叶病害生物防治和生态调控提供理论和实践依据。
1 材料与方法
分别于春季(3月)、夏季(6月)、秋季(9月)、冬季(12月)4个季节在四川省雅安市姚桥镇(东经103°0′55″~103°4′55″,北纬29°59′46″~30°2′48″,海拔622.8 m),采集撑×绿杂交竹健康叶、病叶,杂交竹栽培区面积为6.67 hm2,将样地分为3个区,每个区任选10株杂交竹(重复10次),每株分上、中、下3层,每层的东、西、南、北各方位取两片叶子,共计24片。样品带回实验室用牛肉膏蛋白胨培养基进行分离培养。
附生细菌的分离:每株杂交竹每片叶上随机打孔5个,共得120个直径1 cm的叶圆片,混匀后分成3份。分别放入盛有100 mL灭菌水的三角瓶中,在振荡器上恒温25 ℃振荡1 h之后采用平板稀释分离法[12]进行分离。25~28 ℃下培养2~3 d,用平板计数法进行计数[12]。
内生细菌的分离:分离内生细菌之前,对样品进行表面消毒,之后剪成1.0~1.5 cm的小段,混匀后分为3份,每份加入10 mL无菌水,研碎成浆,分别转入已灭菌的300 mL三角瓶中。每份5 g叶片共加入无菌水50 mL,之后采用平板稀释法进行分离,25~28 ℃下培养2~3 d,用平板计数法进行计数[12]。
叶表附生微生物:微生物密度=(平均每皿菌落数×稀释倍数)∕叶圆片总面积(正反),单位为个/cm2。
叶片内生微生物:微生物密度=(平均每皿菌落数(为同一稀释度的3个平板上菌落平均数)×稀释倍数)∕含菌样品质量,单位为个/g。
相对分离频率=(某种菌分离菌落数/分离总菌落数)×100%。
细菌的鉴定采用16SrDNA分析法[13]鉴定到种,引物选用细菌通用引物PrimerA:5′-AGA GTT TGA TCC TGG CTC AG-3′,PrimerB:5′-GGT TAC CTT GTT ACG ACT T-3′。叶片生理指标的测定分别采用pH计、考马斯亮蓝G-250法和蒽酮法来测定叶片pH值、蛋白质质量分数和可溶性糖的质量分数[14]。
2 结果与分析
2.1 撑×绿杂交竹健、病叶细菌数量的季节性变化
从附生细菌数量的季节性动态变化(表1)来看,健、病叶的细菌数量都呈现出在夏、秋两季达到1 a中的最大值的规律。原因是夏、秋两季是病害的高发期而且也是各种细菌大量繁殖的时期。健康叶附生细菌数量从春季到夏季急剧增加,到秋季达到最大值32.5个/cm2,病叶与之呈现相似的趋势,从春季到夏季急剧增加并在夏季达到1 a中的最大值1 174.8个/cm2。从秋季开始数量又逐渐下降,但速率较缓。可见季节变化对叶面附生细菌数量有明显的影响。另外,健、病叶叶面附生细菌数量差异明显,夏季差异最大,分别为5.9和1174.8个/cm2。可见叶片是否感病对附生细菌也有影响。
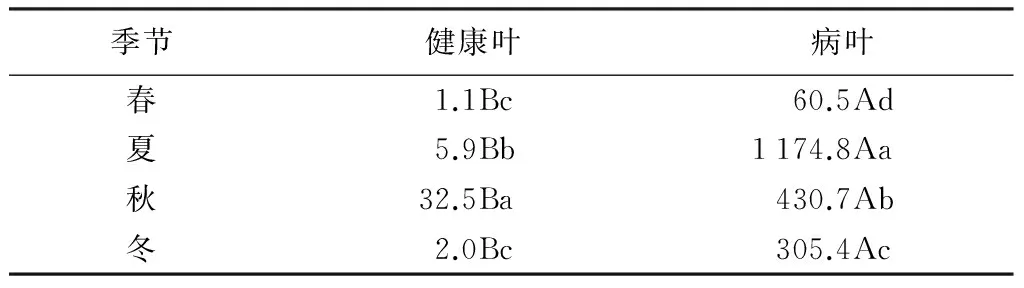
表1 撑×绿杂交竹健、病叶附生细菌数量季节变化 个·cm-2
注:同行不同大写字母表示差异显著(P<0.05);同列不同小写字母表示差异显著(P<0.05)。
健、病叶的内生细菌数量变化趋势相似(表2),春季之后数量急剧升高,到夏季达到最大值,分别为3 406.6和4 733.2个/cm2,夏季之后数量缓慢下降,过了秋季数量急剧下降,到冬季达到最小值,分别为0.7和793.2个/cm2。总体上看,内生细菌数量随季节变化有明显差异,各个季节病叶内生细菌都大于健康叶,且秋季二者之间差异最大。
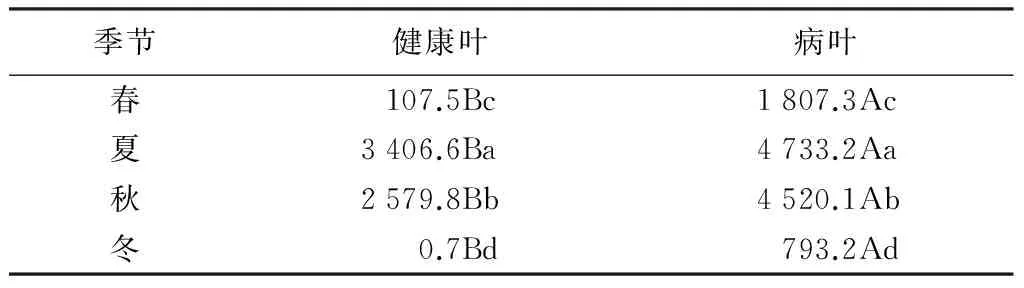
表2 撑×绿杂交竹健、病叶内生细菌数量季节变化 个·cm-2
注:同行不同大写字母表示差异显著(P<0.05);同列不同小写字母表示差异显著(P<0.05)。
2.2 撑×绿杂交竹健、病叶细菌类群特征
2.2.1 撑×绿杂交竹健、病叶附生细菌类群多样性
健康叶叶面附生细菌群落组成及季节演替情况见表3。健康叶叶面共分离鉴定细菌23种。3个季节在健康叶叶面上出现的细菌有2种,占8.70%;在2个季节出现的有15种,占65.22%;仅在1个季节出现的有6种,占26.09%。春季从健康叶叶面分离到细菌12种,相对分离频率达到10%的细菌有1种,是B.subtilis(69.62%),为该季节的优势菌;夏季共分离到4种,优势种是Sphingobacteriumsiyangensis(30.95%)和B.subtilis(42.85%);秋季分离到12种,由B.cereus(32.67%)、L.fusiformis(15.03%)2个种组成优势类群;冬季分离到4种,优势种为B.fusiformis(14.63%)。其中,B.cereus在秋季处于活性高峰,从秋季到冬季在细菌中所占的比例逐渐下降,B.subtilis出现在春、夏两季,且在这两季处于优势地位;B.fusiformis在冬季处于活性高峰,其余2个优势种也仅在1个季节相对分离频率达到优势种标准。从总体来看,B.cereus在健康叶叶面附生细菌的数量组成中占有极其重要的地位,其次S.siyangensisa也是较为稳定的组成成分。
病叶附生细菌有29种。在3个季节出现的有3种,占10.34%;在2个季节出现的有1种,占3.44%;仅在1个季节出现的有25种,占86.21%。春季分离到细菌9种,优势种由B.sp.2(23.41%)和Enterobacteriaceaebacterium(50.77%)组成;夏季共分离到10种,有2个优势种,分别为B.subtilis(40.47%)和Xanthomonadaceaebacterium(35.71%);秋季共分离到10种,其中有3株组成优势类群,分别为L.fusiformis(10.15%)、B.thuringiensis(17.76%)和Erwiniaamylovora(12.18%);在冬季共分离到7种,Pseudomonasoryzihabitans(17.97%)和B.ceres(15.66%)为优势种。在病叶叶面,没有在3个季节均处于优势地位的种。从总体变化趋势看,B.subtilis在细菌的年动态变化中处于重要地位。
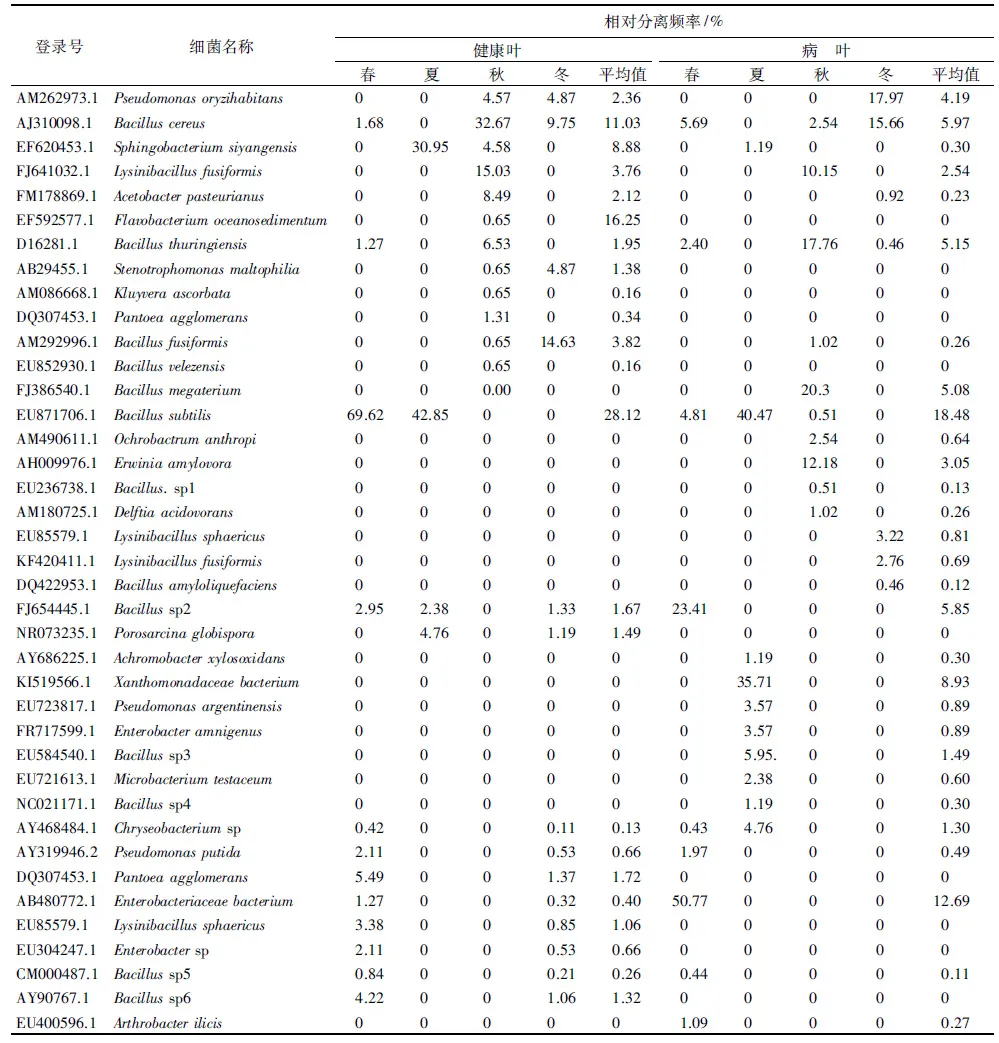
表3 叶面附生细菌的群落组成及季节演替
2.2.2 撑×绿杂交竹健、病叶内生细菌类群
共鉴定健康叶内生细菌14种,其群落组成及季节演替见表4。健康叶内生细菌中,未发现在4个季节或3个季节均出现的菌种;在2个季节出现的有4种,占分离总物种数的28.57%;其余10种都只在一个季节出现,占分离总物种数的71.43%。春季共分离到内生细菌5种,达到优势菌的有2株,分别是B.cereus和L.fusiformis,相对分离频率分别为13.54%和78.56%;在夏季共分离到7种,其中B.fusiformis、Stenotrophomonasmaltophilia和P.plecoglossicida为优势种。相对分离频率分别为16.05%、17.42%和52.84%;秋季共分离到5种,其中以S.siyangensis和Microbacteriumtestaceum的数量优势最明显,相对分离频率为28.97%和66.41%;在冬季,共分离到1种:B.subtilis,占绝对优势。总体来看,在健康叶内生优势类群中,来自芽孢杆菌属(Bacillus)细菌占了一部分,是健康叶内生菌的重要组成部分。
病叶内生细菌共19种。所分离的病叶内生细菌中,没有在4个季节和3个季节都出现的菌种;在2个季节出现的有2种,占分离总株数的10.53%,其余17个种都只在一个季节出现,占分离总株数的89.47%。在春季,分离到5种内生细菌,其中4株为芽孢杆菌;在夏季共分离到7种,5种为优势菌,分别是B.subtilis和E.bacterium(10.08%)、P.argentinensis(47.06%)、P. sp(27.73%)和P.putida(22.69%);秋季共分离4种,相对分离频率最高的为S.siyangensis和M.testaceum分别达到42.25%和43.66%;在冬季也分离到5种,S.maltophilia和P.plecoglossicida占优势分离率都分别为31.87%和65.4%。病叶内生菌优势类群也以假单孢属细菌(Pseudomonas)居多。

表4 叶片内生细菌的群落组成及季节演替
2.3 撑×绿杂交竹健、病叶细菌类群物种丰富度
2.3.1 撑×绿杂交竹健、病叶细菌类群物种丰富度的季节性动态变化
健、病叶面附生细菌丰富度季节变化如表5所示,不同季节健康叶叶面附生细菌物种丰富度的变化趋势有大到小为冬季、春季=秋季、夏季;而病叶上的变化趋势由大到小为夏季=秋季、春季、冬季。健康叶叶面附生细菌物种丰富度在春秋冬三季就均高于病叶,且波动幅度很大。

表5 叶面附生细菌物种丰富度变化
注:同行不同大写字母表示差异显著(P<0.05);同列不同小写字母表示差异显著(P<0.05)。
健、病叶内生细菌丰富度季节变化如表6所示,不同季节健康叶内生细菌物种丰富度的变化趋势由大到小夏季、春季=秋季、冬季;病叶叶面附生细菌物种丰度的变化趋势由大到小同为夏季、春季=冬季、秋季,二者在春、夏两季的物种丰富度基本相同,冬季差异较大。可见,内生细菌物种丰富度随季节变化波动较为明显,而在健、病叶间的差异较小。

表6 叶片内生细菌物种丰富度变化
注:同行不同大写字母表示差异显著(P<0.05);同列不同小写字母表示差异显著(P<0.05)。
2.3.2 撑×绿杂交竹叶部细菌物种丰富度
如表7所示,叶片类型和季节变化对附生细菌的影响不显著。在内生细菌方面,季节变化对细菌物种丰富度的影响显著,而叶片类型对它的影响不显著。可见季节变化是影响内生细菌物种丰富度的重要因素。
2.4 撑×绿杂交竹细菌数量状况与生理指标的关系
细菌数量除受季节和叶片类型的影响外,还受叶片微环境的影响。这个微环境主要就是指叶片上的pH值、蛋白质质量分数和可溶性糖质量分数。
表7 健、病叶类型和季节变化对叶部细菌物种丰富度的方差分析
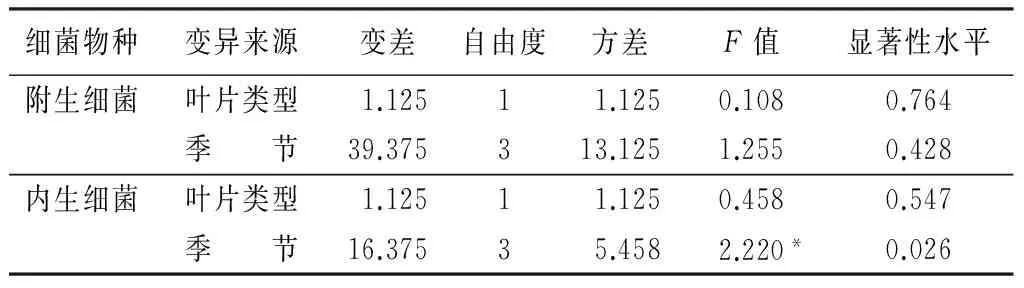
细菌物种变异来源变差自由度方差F值显著性水平附生细菌叶片类型1.12511.1250.1080.764季 节39.375313.1251.2550.428内生细菌叶片类型1.12511.1250.4580.547季 节16.37535.4582.220*0.026
注:*表示a=0.05时显著。
从表8可以看出,健康叶附生细菌的数量和pH值呈正相关,且关系显著,说明健康叶叶面附生细菌的数量随pH的升高而增加。健康叶内生细菌和病叶附生细菌的数量和蛋白质质量分数呈显著关系,而可溶性糖质量分数与4种来源的细菌数量之间关系不显著。总的来看,叶面微环境对附生细菌数量的影响大于内生细菌。
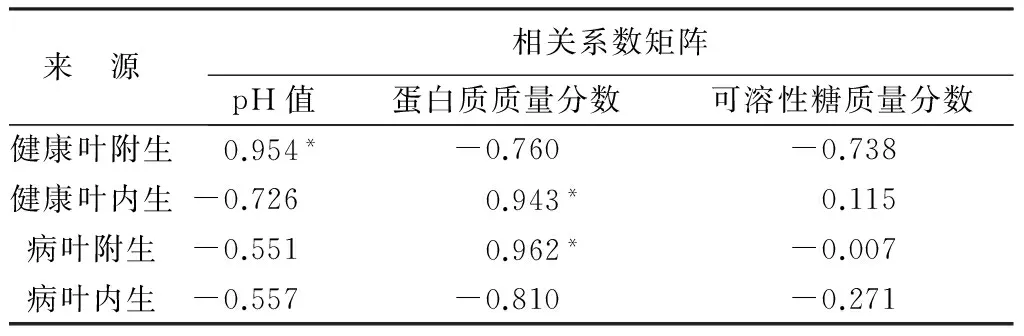
表8 叶部细菌数量状况与生理指标的相关性
注:*表示a=0.05时相关。
3 结论与讨论
通过本研究发现,撑×绿杂交竹病叶叶面附生和叶片内生细菌数量的年平均值均大于健康叶。健、病叶间细菌数量的差异说明杂交竹发病与否与叶部细菌的关系紧密。据Withers H[15]对“细菌数量应答系统”的研究表明:细菌通过可自由扩散的小分子化合物作为信号分子进行个体细胞间信息交流,感应其自身群体密度的变化,当细菌达到一定浓度时,调整整个群体的行为,做出相应的应答。其中AHLs(N-acylhomoserine lactones)介导的细菌数量应答系统参与许多生物学功能的调节,如生物发光,孢子形成,菌体游动,抗生素合成,质粒接合转移,动植物病原菌致病因子的产生等[16]。宋水山等[17]的研究发现AHLs细菌数量应答系统参与病原菌的发病,并发挥着关键作用。这很好地解释了杂交竹病叶叶片细菌数量多于健康叶的原因,即病叶叶部细菌多,AHLs细菌数量应答系统参与病原菌发病的作用也越强。夏、秋两季一般为植物病害的高发期,细菌活动频繁,因此生长于病叶上的细菌数量要高于健康叶,夏、秋两季的细菌数量也远大于春季和冬季。
对于附生细菌,健康叶的平均物种丰富度大于病叶,而对于内生细菌,则病叶的平均物种丰富度大于健康叶。在健康叶上相对分离频率高的物种大多数为芽孢杆菌,如枯草芽孢杆菌(B.subtilis)、蜡样芽孢杆菌(B.cereus)和纺锤芽孢杆菌(B.fusiformis)。芽孢杆菌突出的特征是能产生耐热抗逆的芽孢,这有利于生防菌剂的生产、剂型加工及在环境中存活、定殖与繁殖[18]。此外,它产生的脂肽抗生素对多种真菌和细菌具有强烈的拮抗作用[19-20],而且它还能产生多种细菌素,可抗真菌和细菌病害[21-23]。因此健康叶上分离得到大量的芽孢杆菌。值得注意的是病叶上分离到梨火疫病病原菌(E.amylovora),且为优势种。Hueck[24]和Galan[25]的研究表明:E.amylovora含有三型分泌系统(type III secretion system,TTSS),TTSS能包装及运输一系列效应蛋白穿越宿主的防卫体系,进入膜内释放效应蛋白,产生毒性或者引起一系列过敏性反应,具有强致病性。除此之外,在病叶上分离得到的其他优势种,如S.siyangensis、M.testaceum、X.bacterium等,国内外对其研究相对较少,它们是否为致病菌,仍需进一步的研究。
通过对4个季节健康叶和病叶pH值的测定,发现健康叶叶部的pH值变化范围在6~7,病叶叶部的pH值略高于健康叶,带碱性。这与细菌喜偏碱的环境有关。在酸性条件下细菌一般生长缓慢或者不生长。这也从另一方面解释了病叶叶部细菌数量多于健康叶的原因。氮源是细菌主要的营养来源,因此健康叶内生细菌和病叶附生细菌的数量都和蛋白质质量分数相关。为此我们可以利用改变叶部微环境的方法,调节pH值和蛋白质质量分数来抑制病原菌的生长。

[10] Egan S, Thomas T, Holmstrom C, et al. Phylogenetic relationship and antifouling activity of bacterial epiphytes from the marine algaUlvalactucabrief report[J]. Environmental Microbiology,2000,2(3):343-347.
[11] 何红,邱思鑫,胡方平,等.植物内生细菌生物学作用研究进展[J].微生物学杂志,2004,24(3):40-45.
[12] 方中达.植病研究方法[M].北京:中国农业出版社,1998.
[13] 龙雯,陈存社.16SrRNA测序在细菌鉴定中的应用[J].北京工商大学学报:自然科学版,2006,24(5):10-12.
[14] 熊庆娥.植物生理学实验教程[M].成都:四川科学技术出版社,2003.
[15] Withers H, Swift S, Williams P. Quorum sensing as an integral component of gene regulatory networks in gram-negative bacteria[J]. Current Opinion in Microbiology,2001,4(2):186-193.
[16] Swift S, Throup J P, Williams P, et al. Quorum sensing: a population-density component in the determination of bacterial phenotype[J]. Trends in Biochemical Sciences,1996,21(6):214-219.
[17] 宋水山,贾振华,高振贤,等.植物伴生细菌数量应答系统的研究进展[J].微生物学通报,2004,31(2):117-120.
[18] 于淑池.植物真菌病害生防芽孢杆菌的研究进展[J].通化师范学院学报,2007,28(8):52-54.
[19] Dimkic' I, Zivkovic' S, Beric' T, et al. Characterization and evaluation of two bacillus strains, SS-12.6 and SS-13.1, as potential agents for the control of phytopathogenic bacteria and fungi[J]. Biological Control,2013,65(3):312-321.
[20] 张荣胜,王晓宇,罗楚平,等.解淀粉芽孢杆菌Lx-11产脂肽类物质鉴定及表面活性素对水稻细菌性条斑病的防治作用[J].中国农业科学,2013,46(10):2014-2021.
[21] Martin D F, Priest F G, Todd C, et al. Distribution of beta-glucanases within the genus Bacillus[J]. Applied and Environmental Microbiology,1980,40(6):1136-1138.
[22] Tanaka H, Watanabe T. Glucanases and chitinases of bacillus circulans WL-12[J]. Journal of Industrial Microbiology,1995,14(6):478-483.
[23] 王倩,孙珊,郝林.枯草芽孢杆菌W321产抑菌物质的理化性质及抑菌谱研究[J].食品工业, 2012,33(11):110-112.
[24] Hueck C J. Type III protein secretion systems in bacterial pathogens of animals and plants[J]. Microbiology and Molecular Biology Reviews,1998,62(2):379-433.
[25] Galan J E, Collmer A. Type III secretion machines: bacterial devices for protein delivery into host cells[J]. Science,1999,284:1322-1328.
Foliar Bacterial Diversity ofBambusapervariabilis×Dendrocalamopsisdaii
Peng Yan, Mao Cui, Zhu Tianhui, Liu Yang(Sichuan Agricultural University, Ya’an 625014, P. R. China)//Journal of Northeast Forestry University,2015,43(6):67-71,97.
Bambusapervariabilis×Dendrocalamopsisdaii; Healthy leaves; Diseased leaves; Bacterial diversity; Biological control
1)国家科技基础条件平台项目(2005DK21207-13)。
彭艳,女,1992年12月生,四川农业大学林学院,硕士研究生。E-mail:1362582576@qq.com。
朱天辉,四川农业大学林学院,教授。E-mail:zhuth1227@126.com。
2013年12月1日。
S763.13
责任编辑:潘 华。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
湖北农机化(2020年4期)2020-07-24
世界农药(2019年4期)2019-12-30
今日农业(2019年11期)2019-08-15
山东林业科技(2019年2期)2019-06-03
乡村地理(2018年2期)2018-09-19
林业科技(2017年6期)2017-12-06
中国果业信息(2017年4期)2017-05-09
光散射学报(2016年1期)2016-04-19
园艺与种苗(2015年10期)2015-02-27
