
蜱组胺结合蛋白与渗透肽TAT在杆状病毒表达体系中的融合表达和功能分析
2015-03-09 01:54王亚楠周勇志张厚双龚海燕周金林
中国预防兽医学报 2015年6期
王亚楠,曹 杰,周勇志,张厚双,龚海燕,周金林*
(1.上海师范大学 生命与环境科学学院,上海 200234;2.中国农业科学院上海兽医研究所农业部动物寄生虫学重点实验室,上海 200241)
蜱组胺结合蛋白(Histamine binding protein,HBP)是由蜱的唾液腺分泌的具有特异性结合组胺的一类功能性蛋白的总称,属于脂质运载蛋白(Lipocalins)家族的一员[1]。HBP 最早发现于扇头蜱属的唾液腺中,目前已在网纹革蜱(D.reticulatus)、肩突硬蜱(I.scapularis)、扇头蜱属(Rhipicephalus)、美洲花蜱(A.americanum)的唾液腺中发现了HBP[2]。本实验室从亚洲璃眼蜱的唾液腺组织中克隆了684 bp 编码HBP 的基因,并原核表达了HBP,命名为亚洲璃眼蜱组胺结合蛋白(HBP),根据其药物开发应用潜力,申请并获得了中国发明专利[3]。
TAT 蛋白转导肽是人类免疫缺陷病毒1 型(HIV-1)编码的一段富含碱性氨基酸、带正电荷的多肽,属于蛋白转导域家族的一员[4]。其全长及11 个碱性氨基酸富集区的核心肽段(YGRKKRRQRRR),不仅能够在包括蛋白质、多肽及核酸等多种外源生物大分子的跨膜转导过程中具有重要作用,而且能够携带这些外源生物大分子通过活体细胞的各种生物膜性结构(如细胞膜和血脑屏障等)并发挥生理功能,TAT 作为一种重要的渗透肽,在生物大分子及药物传递等领域具有广泛应用前景[5]。
本研究根据TAT 具有跨膜转导的作用,为了增强HBP 在生物被膜传递和吸收效率,进行TAT 与HBP 融合表达,为蜱组胺结合蛋白的开发应用奠定基础。
1 材料和方法
1.1 主要实验材料 pGEX-HBP 重组质粒由本实验室构建;pFastBacHTa、DH10Bac 感受态细胞及sf9细胞由本实验室保存;His 单克隆抗体(MAb)购自康为世纪生物科技有限公司;FITC 标记的标记羊IgG(IgG-FITC)购自KPL 公司;羊抗鼠IgG-HRP 购自北京鼎国昌盛生物技术有限公司;SF900-Ⅱ无血清培养基和购自Gibco 公司;Cellfectin®ⅡReagent 转染试剂、杆粒中抽试剂盒购自Invitrogen;Ni-NTA His Bind Restin、Ni-NTA Buffer Kit 购自Novagen 公司;CytoBuster 蛋白抽提试剂购自购自Millipore 公司;组胺检测ELISA 试剂盒购自NeogenVeratox®;免抗HBP 阳性血清由本实验室制备。
1.2 p Fast-HBP-TAT重组转移质粒的构建 以pGEX-HBP 为模板对HBP 的序列进行PCR 扩增,设计克隆引物时在TAT 的3' 端和HBP 的5' 端加入Eco RⅠ酶切位点。
TAT 的合成:TAT 正链5'-TACGGCCGTAAGA AGCGCCGCCAGCGCCGTCGT-3';TAT 反链5'-GA ATTCACGACGGCGCTGGCGGCGCTTCTTACGGCC GTA-3',将合成的TAT 的正反链等摩尔浓度混合,95 ℃加热10 min,室温放置1 h,使之形成编码TAT 的双链DNA。
对合成的TAT 双链DNA 片段和经Eco RⅠ限制性内切酶酶切后的HBP 胶回收产物连接,将连接产物克隆于pMD18-T 中构建重组质粒pMD-HBPTAT。并以其为模板,参照pFastBacHTa 载体多克隆酶切位点序列设计引物:5'-CGGGATCCGTAC GGTCGTAAAAAA-3'(Bam H Ⅰ)/5'-GCTCTAGAGCG ATTTCGGCAATATTTTC-3'(Xba Ⅰ),以pMD-HBP为模板,经PCR 扩增HBP-TAT 片段。并将其通过Bam HⅠ和XbaⅠ双酶切,克隆于pFastBacHTa 中,构建昆虫重组转移质粒pFast-HBP-TAT。并分别用特异性引物和载体测序通用引物进行pFast-HBPTAT 的PCR 鉴定。
1.3 rBac-HBP-TAT重组杆粒的构建 将pFast-HBP-TAT 转化至DH10Bac 感受态细胞中,通过蓝白斑筛选,PCR 筛选阳性重组杆粒rBac-HBP-TAT。并按照Bac-to-Bac 杆状病毒表达系统使用手册合成M13 上下游引物,进行重组杆粒的PCR 鉴定。
以杆粒抽试剂盒提取鉴定为阳性的rBac-HBPTAT,使用转染试剂将其转染于sf9 细胞单层中,同时做空白对照,转染后于不同时间观察细胞病变(CPE),对重组杆状病毒进行病毒蚀斑纯化和扩增并对P2 代重组杆状病毒进行鉴定。
1.4 HaHBP-TAT融合蛋白的检测和鉴定 分别收集P2 代重组杆状病毒扩增后的sf9 细胞的培养基上清液和细胞,进行重组蛋白的SDS-PAGE 检测。并采用半干法转印至PVDF 膜上、5 %脱脂奶粉封闭后,分别以His MAb(1∶4 000)和HBP 多抗血清(1∶200)为一抗,羊抗鼠IgG-HRP(1∶4 000)为二抗,最后经DAB 显色后进行western blot 鉴定。
1.5 重组蛋白HaHBP-TAT的间接免疫荧光(IFA)检测 鉴定为阳性的P3 代重组杆状病毒感染sf9 细胞,同时做正常sf9 细胞空白对照,48 h 后弃培养液,以预冷的甲醇固定细胞20 min,以His MAb(1∶4 000)为一抗,羊抗鼠IgG-FITC(1∶1 000)为二抗,进行融合蛋白表达的IFA 鉴定。
1.6 表达的融合蛋白(HaHBP-TAT)纯化和组胺结合试验 通过离心收集大批量P4 代重组杆状病毒感染的细胞沉淀,以CytoBuster 细胞裂解液悬浮,并加入一定量的蛋白酶抑制剂,低功率超声后,离心,收集上清,0.22 μm(低蛋白结合)滤膜过滤,并参照Ni-NTA Buffer Kit 蛋白纯化说明书对表达的融合蛋白进行纯化。
将真核表达纯化的融合蛋白HBP-TAT 纯化后在不同蛋白浓度(10 μg/mL、5 μg/mL、2.5 μg/mL、1.25 μg/mL)的条件下与等体积的HRP 标记的组胺共同孵育后,检测OD650nm值,设TAT 为阴性对照。
2 结果
2.1 pFast-HaHBP-TAT重组转移质粒的构建 将HBP(5' 端引入Eco RⅠ位点)和TAT(3' 端连连接有酶切位点Eco RⅠ)的基因片段的胶回收产物经限制性内切酶Eco RⅠ酶切连接,将其克隆于pMD18-T中,经测序后确定重组基因片段构建正确。
同时使用两组不同的引物对扩大培养的样品进行PCR 鉴定,结果表明,特异性引物扩增出的基因片段约为650 bp,而使用交叉引物扩增出的片段大小大于特异性引物扩增出的片段大小,表明重组基因片段HBP-TAT 成功连接至真核表达载体pFast-BacHTa(图1)。
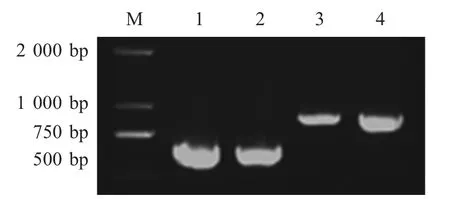
图1 pFast-HBP-TAT 转移重组质粒的PCR 鉴定Fig.1 Identification of recombinant pFast-HBP-TAT by PCR
2.2 rBac-HBP-TAT重组杆粒的构建 pFast-HBPTAT 转化于DH10Bac 感受态细胞中,通过蓝白斑筛选,菌液PCR 筛选重组杆rBac-HaHBP-TAT。PCR鉴定结果表明:阳性重组杆粒rBac-HBP-TAT 通用引物M13 扩增后片段约为3 000 bp(2430+642=3072 bp),特异性引物扩增后片段约为650 bp;空杆粒rBac 通用引物M13 扩增后片段约为2 500 bp,特异性引物扩增阴性(图2)。

图2 rBac-HBP-TAT 重组杆粒构建菌液PCR 鉴定Fig.2 Identification of rBac-HBP-TAT by PCR
2.3 真核表达蛋白的SDS-PAGE和western blot鉴定 将收集的细胞裂解液进行SDS-PAGE 电泳分析,显示在约31 ku 处出现条带(图3A)。分别使用His MAb 和HBP 免疫小鼠后获得的多抗血清作为一抗进行western blot 分析,结果出现与预期分子量大小相符的特异性条带,表明His 标签的融合蛋白HBP-TAT 在sf9 细胞中得到表达(图3B、C)。

图3 HBP-TAT 的SDS-PAGE(A)和western blot(B)鉴定Fig.3 SDS-PAGE(A)and western blot analysis(B)of rHBP-TAT protein
2.4 真核表达蛋白的IFA检测 重组杆状病毒接种于sf9 细胞后,经IFA 检测结果显示,感染重组杆状病毒的sf9 细胞产生绿色荧光而野生型的sf9 细胞无绿色荧光,证明重组蛋白HBP-TAT 在sf9 细胞中表达(图4)。

图4 间接免疫荧光检测重组蛋白的表达(400×)Fig.4 The IFA assay of the recombinant protein(400×)
2.5 组胺结合试验 真核表达融合蛋白(HBP-TAT)经Ni-NTA His·Bind Resin 亲和纯化后,在不同浓度的条件下与HRP 标记的组胺共同孵育后检测OD650nm值,结果表明真核表达的融合蛋白HBP-TAT 具有与组胺结合的能力,而对照组TAT 不具备与组胺结合的能力(图5)。

图5 HBP-TAT 在不同蛋白浓度条件下的组胺结合试验Fig.5 The histamine binding assay of HBP-TAT at different concentrations
3 讨论
不同种蜱细胞中的HBP 高度保守,即均含有一个Lipocalin 折叠,HBP 结构中均存在一个由多条多肽链围绕而成的β 桶状结构,在桶状结构内部存在一个或两个组胺结合位点。由于结合位点的数目和在桶内的位置不同,不同的组胺结合蛋白和组胺的亲和力也不相同[6]。研究表明,亚洲璃眼蜱HBP 重组蛋白的高级结构对于研究组胺结合蛋白和组胺结合的机制非常重要,Paesen 等已从具尾扇头蜱(R.appendiculatus)细胞内成功克隆并经真核表达(杆状病毒表达系统)结晶出结构接近于天然蛋白的RaHBP、RaHBP2 及RaHBP3[7],为使表达的融合蛋白的结构更接近于天然蛋白,所以本研究选择杆状病毒蛋白表达系统对融合蛋白进行表达。
杆状病毒蛋白表达系统具有与高等真核生物相似的转录和翻译蛋白的能力并能在翻译后进行修饰和加工,在蛋白结构上更加接近天然蛋白,具有良好的生物学活性[8]。本实验对TAT 融合的HBP 蛋白进行了真核表达,结果显示表达的融合蛋白分子量与预期结果一致,分子量大小基本相同。经western blot 分析显示,真核表达的融合蛋白HBP-TAT 能够与由原核表达的重组蛋白制备的多抗血清发生反应,证明其与原核表达的HBP 具有相同的免疫原性。
此外,组胺结合试验表明,真核表达纯化的HBP-TAT 具有与组胺结合的能力,表明TAT 融合未影响其结合组胺功能,可以进行后续的实验研究。
[1]Mans B J.Tick histamine-binding proteins and related lipocalins:potential as therapeutic agents[J].Curr Opin Invest Drugs,2005,6(11):1131-1135.
[2]Chinery W A,Ayitey-Smith E.Histamine blocking agent in the salivary gland homogenate of the tick Rhipicephalussanguineus[J].Nature,1977,265(5592):366-367.
[3]周金林,李庄,石磊,等.一种亚洲璃眼蜱的组胺结合蛋白HaHBP 基因序列和重组表达与应用[P]:中国CN101928709A,2010-12-29.
[4]Mbonye U,Karn J.Transcriptional control of HIV latency:cellular signaling pathways,epigenetics,happenstance and the hope for a cure[J].Virology,2014,454-455:328-339.
[5]吴永红,张成岗.HIV-1 TAT 蛋白转导肽的研究进展[J].中国生物工程杂志,2010,(10):66-73.
[6]Schlehuber S,Skerra A.Lipocalins in drug discovery:From natural ligand-binding proteins to 'anticalins?[J].Drug Dis Today,2005,10(1):23-33.
[7]Paesen G,Adams P,Harlos K,et al.Tick histamine-binding proteins:isolation,cloning,and three-dimensional structure[J].Molecular cell,1999,3(5):661-671.
[8]付凡,陈蔚.利用Bac-to-Bac 杆状病毒表达系统在家蚕中表达人生长素[J].蚕业科学,2011,(02):320-324.
猜你喜欢
山东畜牧兽医(2022年10期)2022-12-06
当代水产(2021年7期)2021-11-04
昆明医科大学学报(2021年5期)2021-07-22
中国合理用药探索(2020年9期)2020-10-13
健康必读(上旬刊)(2019年3期)2019-10-15
江苏农业学报(2019年1期)2019-09-10
河南农业科学(2018年9期)2018-09-28
飞碟探索(2016年12期)2016-12-10
浙江大学学报(医学版)(2013年6期)2013-12-09
浙江海洋大学学报(自然科学版)(2012年1期)2012-07-19
