
中国酶工程的兴旺与崛起
2015-03-08 06:28黎高翔
生物工程学报 2015年6期
黎高翔
自1970年开始,在酶制剂工业的基础上,我国酶工程研究只局限于当时兴起的固定化酶及细胞、生物传感器、生物反应器以及天然酶诱变育种的酶制剂工业。在1988年召开的全国固定化生物催化剂会议上,已有大量成果问世,为中国酶工程的发展打下了坚实的基础。
但是,早在1971年美国工程基金会就已组织了第一届国际酶工程会议,事隔10年,直至1981年在日本召开的第六届国际酶工程会议上,才以张树政院士为代表的我国科研人员首次参会。当时,我国酶工程领域研究大大落后于国际水平。
为了适应国际酶工程的迅猛发展,加强国内外同行的学术交流与合作,在邹承鲁院士、张树政院士和国内著名专家倡议下,中国微生物学会于 1989年 7月成立了酶工程专业委员会,当年就立即组织了12人代表团赴日本参加第十届国际酶工程会议,并与日方商约好每两年召开一次中、日酶工程会议。直至2004年韩国要求加入形成了中、日、韩三方的酶工程会议。至现在,25年中连续成功举办了十四届中、日、韩酶工程会议,已形成了亚洲酶工程的轴心。
1997年,经国际酶工程组委会批准,由我国酶工程专业委员会在北京主办了第十四届国际酶工程会议[1],会议由南开大学俞耀庭教授和国际酶工程组织委员会委员黎高翔研究员任大会主席。会上,邹承鲁院士、张树政院士、田波院士、王志珍院士等我国8位酶学专家作了特邀报告。显示了中国酶工程研究的初步成果,获得国际上的一致好评。此次大会云集了国际上很著名的酶工程专家教授,如美国Arnold F H教授、Klibanov A M教授、Blanch H B教授、Dordick J S教授、Russell A J教授、Clark D S教授、瑞典Mosbach K教授、德国Wandrey C教授、Scheller F教授、日本Tanaka A教授、Karube I教授、Shimizu S教授和以色列Katchalski-Katzir E教授等,是一次国际上高水平的酶工程盛会,与会者充分领略了国际酶工程的新领域、新思想、新技术。
这之后的 1989–2014年,我国共举办了 9届全国酶工程会议,酶工程的研究和应用在国内得到广泛重视,科研工作者们群策群力,扩展了酶工程研发的很多新领域,历经25年的奋斗拼搏,在我国各科研院校形成了许多优秀的科研队伍,建立了许多酶工程研发的技术平台和中心,学术水平迅速提高,在酶工程的主要领域,已赶上或有些已超越了国际先进水平,有些获得国家科学技术发明和进步奖,为国家酶工程产业化作出了重要贡献。
1 酶的基因工程与蛋白质工程
工业生物催化在20世纪 90年代兴起,与蛋白质定向进化、基因组学和蛋白质组学技术的发展有关。工业生物催化的核心是酶的应用,与传统的化学催化相比较,生物催化有位点专一性和立体专一性的优势,人们可以按人类的意愿进行“再进化”手段,而不需要了解酶的结构,可以采取基因克隆、随机突变或杂交、定点突变,用易错PCR方法构建突变库,结合高通量筛选策略来提高酶的活性、稳定性、立体选择性及非水反应性能。
我国在过去25年中 (至2014年),共9届的全国酶工程会议上,发表论文总数为978篇,其中酶基因工程与蛋白质工程论文达到356篇,占总论文数36.4% (表1)。是一次很大的飞跃和崛起[2-10]。《生物工程学报》曾于2009年与2012年结合酶工程会议出版了“酶工程”系列专刊[11-12]。
木聚糖酶是半纤维素降解的关键酶,我国用橄榄绿链霉菌基因转到毕赤酵母 Pichia pastoris获得高效表达。酶活性为1 200 U/mL,比活性为2 869 U/mg。具非常好的抗蛋白酶降解能力[3]。嗜酸真菌Biospora sp. MEY-1四个基因成功克隆到毕赤酵母异源表达,重组酵母XYL11酶比活力为1 8831 U/mg,90 10℃ min仍保留87%以上活力。降解燕麦木聚糖主要产木糖和木二糖,具有很好抗蛋白酶降解的能力[8]。用黑曲IME-216产木聚糖基因克隆到酿酒酵母中表达,活力提高到 90 000 U/mL[8],其余 30多篇木聚糖酶基因在Escherichia coli等中表达的论文不再一一详述。
饲料用酶已成为世界工业酶产业中增长最快、实力最强的酶工业。植酸酶是饲料中植酸降解为无机磷酸和肌醇的饲料添加剂,中国农业科学院饲料研究所将黑曲霉Aspergillus niger 963植酸酶基因重组到毕赤酵母中得到高效表达,酶活力达 8×105IU/mL,比原黑曲霉产量提高了3 000倍以上,大大高于国外报道的工程菌[4,11],国内已建立了多家生产企业。
纤维素酶是多酶的复合酶系,非理性设计是目前纤维素酶定向进化的方法,木霉纤维素外切酶和纤维素内切酶已在噬菌体展示功能。目前已经克隆表达一批中碱性纤维素酶的基因,可用于纺织和洗涤剂,造纸工业用途的中性内切纤维素酶工程菌优化培养,酶活可达32 529 U/mL[10],提高了原株的7.8倍。将福寿螺 Ampullaia crossean多功能纤维素酶基因在E. coli中表达,得到高比活力纤维素酶系,对木聚糖、对硝基酚纤维二糖苷、羧甲基纤维素有很好的水解活性[5]。克隆拟康木霉S38 Swollenin基因可能是破坏氢键,是真菌降解纤维素酶系的组成部分[6]。此外,我国还开展了黄翅大白蚁木质纤维素降解酶系的研究[10]。
脂肪酶是生物合成中一个重要酶类。目前我国已建立了通过理性设计成熟的 3个脂肪酶基因改造、生产和应用的技术平台。用基因改组技术将抗展青霉 Penicillium expansum FS 8486酶活力提高了317%[7]。对脂肪酶“蓋子”结构进行了定点突变,获得开蓋型脂肪酶,酶比活力提高5.7倍,两相催化效率提高1.8倍[8]。我国还构建了表面展示工程菌、E. coli工程菌、毕赤酵母工程菌、高密度培养的技术平台,假丝酵母 Candida sp. 99–125脂肪酶活力达到15 000 U/mL以上[9]。克隆米黑根霉 R. miehei脂肪酶在两个毕赤酵母成功表达,最高酶活力达到18 000 U/mL。4 6℃个月酶活力不下降,12 h生物柴油产率超过95%[9]。华根霉Rhizopus chinensis脂肪酶基因克隆到毕赤酵母中成功表达,两种共表达的伴侣蛋白可使酶活力提高30%,酶活力达16 200 U/mL[10-11]。同时,亦开展了对有机溶剂耐受性、热稳定性好的细胞结合脂肪酶的研究。
淀粉酶是非常重要的工业用酶,占酶制剂市场 25%。目前工业主要是高温酶,来自深海液口嗜热厌氧古细菌热球菌属Thermococcus产胞外耐热高温酶,最适温度95 ℃,100 ℃仍有60%活力,用PCR法克隆此酶基因,并在E. coli中得到表达[7]。又从云南腾冲分离两株耐热生淀粉生产的Geobacillus细菌,经纯化后,比活力分别为1 320和890 U/mg[7]。用PCR法克隆了嗜碱芽胞杆菌Bacillus alcalophilus基因,其中枯草芽胞杆菌异源表达,活性为450 U/mL[9]。中温酸性α-淀粉酶,对淀粉加工有节能降耗作用,Bacillus sp. α-淀粉酶基因与B. amyliquefaciens α-淀粉酶基因有98%的同源性[7]。
甘露聚糖酶属于半纤维素酶,适用于甘露寡糖制备。肇东市日成酶制剂公司用黑曲霉工程菌酸性β-甘露聚糖酶表达活力达20 000 U/mL,为目前生产菌株水平的25倍,处于真菌基因工程菌领先地位[10]。利用DNA shuffling定点突变获得耐高温耐酸的A. tabescens MAN 47 β-甘露聚糖酶突变体,高温80 ℃、pH 4.0酶活力为野生型的10倍。定向引入N-糖基化位点,实现了野生型和突变型在毕赤酵母中的表达,热稳定性、酸稳定性、蛋白酶抗性均得到改善。也获得了比野生型甘露聚糖酶活性高 3–5倍的 3个突变体[10-11]。从嗜热网球菌中克隆获得热稳定的β-1,4-甘露聚糖酶,在80 ℃高温低渗油井中、对羟基瓜尔胶有最高活力。该酶半衰期为46 h[10]。
漆酶为含铜多酚氧化酶,为木质素分解酶,亦可催化合成酚类、芳香胺的低聚物。担子菌漆酶酶基因克隆到毕赤酵母中表达活力为9.03 U/mL,为原始菌株的3倍[5]。野生革耳Panus rudis漆酶转化到毕赤酵母分泌表达,通过定点突变及随机突变后,酶比活力为16.17 U/mg,提高了4.4倍[7]。新型海洋细菌漆酶经氨基酸定点突变,突变体半衰期延长3倍,可溶性蛋白较优化前提高244%。发酵产率为野生型的 7.9倍[10]。热带白腐真菌菌株漆酶酶活力达11 400 U/L (愈创木酚法)[7]。
普鲁兰酶又称茁霉多糖酶,属解枝酶,分解α-1,6-葡萄糖苷键。嗜热古菌Thermococcus sp.HJ21产胞外高温普鲁兰酶,最适温度95 ℃,酶活力在100 ℃、2 h仍有50%以上活力[7]。通过PCR技术克隆了此酶基因。温泉中厌氧芽胞杆菌属Anoxy bacillus sp. p28克隆到普鲁兰酶基因序列,构建了重组质粒。转化到E. coli中,产物为单一麦芽三糖,为Ⅰ型普鲁兰酶。从腾冲热泉中分离出耐热厌氧芽胞杆菌普鲁兰酶基因导入枯草芽胞杆菌中得到表达。60 ℃、48 h仍保留 50%以上活力,胞外酶活力 42 U/mL,表达活力提高了40倍[10]。南京百斯杰生物工程公司通过基因敲除及重组技术,改造野生芽胞杆菌普鲁兰酶基因,与葡萄糖淀粉酶制成复合酶,可制备出DE值超过96.5的葡萄糖,商品名为High DEX系列,满足了葡萄糖生产,达到国际水平[10]。
青霉素酰化酶是 β-内酰胺类抗生素工业的关键酶。我国已成功进入酶合成抗生素工业,青霉素酰化酶通过诱变获得突变株活力为 820 U/mL[2]。克隆巨大芽胞杆菌青霉素酰化酶基因在枯草杆菌中表达活力为30 U/mL[4]。青霉素酰化酶在枯草芽胞杆菌中分泌表达,表达量为864 U/L,是野生型类产杆菌A. faecalis产量的144倍[5]。粪产杆菌的青霉素酰化酶在E. coli中分泌表达,改进培养的工程菌酶活力>2 000 U/L。7-氨基头孢烷酸酰化酶 (GL-7-ACA酰化酶) 可转化头孢菌素C,在E. coli中表达,最高酶活力达266 U/L[5],用Eupergitc载体固定化巨大芽孢杆菌青霉素酰化酶,连续转化生产30批次,活力未见下降[6]。
D-氨基酸氧化酶可催化D-氨基酸生成相应的酮酸和氨,此酶与 7-氨基头孢烷酸酰化酶(7-ACA酰化酶) 两步法生产头孢菌素重要原料7-氨基头孢烷酸 (7-ACA)。用甲醇酵母表达的D-氨基酸氧化酶,构建了高表达的毕赤酵母重组菌。14 L罐发酵活力达8 000–1 2000 U/L[5]。构建了毕赤酵母融合表达的 D-氨基酸氧化酶活力为1 700 U/L[5],还构建了两种重组GL-7-ACA酰化酶,组成型菌株达到1 500 U/L,诱导型菌株达900 U/L,固定化酶转化率达到97%[5]。三角酵母D-氨基酸氧化酶基因转到E. coli中,表达活力为23.3 U/mL[5],D-氨基酸氧化酶固定化在 Amberzyme环氧树脂上转化 14批次,活力未见下降[6]。对羟基苯甘氨酸是半合成β-内酰胺类抗生素的重要中间体,可通过海因酶和 D-氨甲酰水解酶两步催化制备,通过对D-氨甲酰水解酶随机突变,在E. coli中可溶性提高了6倍[6],重组表达E. coli的D-海因酶可溶性差,共表达分子伴侣可将可溶性表达量提高约3倍[7]。
羰基还原酶不对称还原羰基化合物广泛用于制备手性醇。天蓝色链霉菌羰基还原酶制备(S)-4-氯-3-羟基丁酸乙酯。其重组菌E. coli BL21制备了 (S)-4-氯-3-4-苯基丁酸乙酯和 (S)-邻氯扁桃酸甲酯,转化率和ee值均高达99%以上[9]。面包酵母羰基还原酶基因同源表达产物对映体ee值和产率均有提高。近平滑假丝酵母基因组中克隆出新型 (S)-型羰基还原酶,重组菌E. coli BL 21能制备 (S)-苯基乙二醇,光学纯达99.1%,产率为89.6%[8,11]。
β-葡萄糖苷酶是纤维素酶系的重要组分,可将纤维二糖、可溶性纤维寡糖水解成葡萄糖及相应配基,水解β-糖苷键及合成新的糖衍生物。瑞士木霉β-葡萄糖苷酶,通过基因敲除、氨基酸突变,突变体酶活力比野生型提高了 143倍[9]。新鞘氨醇杆菌β-葡萄糖苷酶基因在E. coli成功表达,可转化异黄酮糖苷生成相对应的苷元[10]。台湾乳白蚁肠道中的 β-葡萄糖苷酶,用原核表达系统进行异源表达,突变酶较野生型酶活力提高了2.6倍。亦提高了葡萄糖耐受性及热稳定性[10]。斜卧青霉 β-葡萄糖苷酶进行基因改造,通过转化获得 3个表达菌株,酶活力比原始菌株提高3.4–3.7倍[10],从非解朊栖热菌分离到耐热β-葡萄糖苷酶基因在E. coli中克隆表达,酶活力提高17倍[4]。从海洋宏基因组筛选到葡萄糖耐受的β-葡萄糖苷酶克隆到E. coli高效表达,葡萄糖浓度低于400 mmol/L,对酶有促进作用,达到 1 000 mmol/L葡萄糖浓度,酶活性为50%[8,11]。利用 DNA改组和定点突变提高 β-葡萄糖苷酶稳定性,4个突变体与野生型相比,61 ℃热失活半衰期提高了14.16、68和44倍。耐热性有明显提高[8]。
半乳糖苷酶,又名乳糖酶,能将乳糖分解成葡萄糖和半乳糖基,可以合成低聚糖。通过宏基因组法从海底泥中获得高效转糖基的 β-半乳糖苷酶,在E. coli中可溶性表达,对有机溶剂有耐受性,合成低聚半乳糖产量可达51.6%[9]。曲霉α-半乳糖苷酶固体发酵,酶活力达305 IU/g[6]。硫磺矿硫化叶菌Sulfolobus solfataricus P2的 β-半乳糖苷酶进行分子改造构建了突变体,突变酶F441Y生产低聚半乳糖,产率达61%,较野生型高10%左右[9]。
腈水合酶在酰胺、羧酸及其衍生物合成中有重要应用价值,催化丙烯腈生产丙烯酰胺,诺卡氏菌Nocardia sp. YS-2002发酵活力最高达6 000国际单位,在E. coli及毕赤酵母中得到表达[5]。
菊粉酶是水解菊粉的 β-2,1-D-果聚糖果糖苷键的水解酶,产低聚果糖,分内切酶和外切酶两种。将黑曲霉菊粉内切酶基因克隆到毕赤酵母得到高效表达,重组酶活力达128 U/mL,达到国际水平[9]。
海藻糖合成酶,海藻糖在食品、医学、轻工业领域广泛应用,可用单酶法从麦芽糖生产海藻糖,通过酶分子的理性设计和DNA shuffling技术从两个 GRAS菌种和两个嗜热菌中克隆到海藻糖合成酶基因并成功进行了高效表达,已实现了工业化生产[5]。
中性蛋白酶广泛用于工业化生产,枯草芽胞杆菌AS1398基因Npr的重组质粒转入B. subtilis AS 1398,获得多拷贝的重组菌,连续传代 30代,重组菌质粒稳定性保持100%,蛋白酶表达水平提高1倍以上,达到24 000–28 000 U/mL[10]。有高效杀线虫毒力的侧孢短芽胞杆菌株经优化培养获得蛋白酶的活力达12 379 U/mL,比优化前提高5倍,应用B. subtilis WB600表达体系,构建随机突变文库,获得热稳定的突变体,为生物杀虫剂提供依据[8]。人基质金属蛋白酶与肿瘤转移关系密切,在E. coli中成功克隆表达了该酶,为研究该酶与肿瘤转移机理和寻找该酶特异抑制剂奠定基础[5]。
葡聚糖酶属水解酶,分内切酶和外切酶,用于啤酒生产和饲料添加。嗜热青霉 β-1,3-1,4-葡聚糖酶基因克隆到原核表达载体并转入E. coli BL 21诱导表达,酶活力240 U/mL[8],链霉菌S27内切β-1,3-葡聚糖酶基因在E. coli中高效表达,可水解昆布多糖等,可抑制致病真菌和产毒素真菌[8]。长梗木霉内切葡聚糖酶基因导入毕赤酵母表达,重组菌Ⅲ酶活力达110 U/mL,优化后提高 50%[8]。点突变极耐热 Thermotoga maritima内切葡聚糖酶 Cell-12B,酶最适温度95 ℃,90 ℃仍保留70%活力[10]。
N-乙酰神经氨酸是生物体内一种最重要的唾液酸,唾液酸糖链参与许多生命过程,CMP-唾液酸能促进神经细胞再生,对CMP-唾液酸合成酶基因进行碱基改造,在E. coli中得到高效表达,酶表达量占总蛋白26.5%,酶活力为100 U/L,为出发菌株的850倍[4]。
黄曲霉毒素解毒酶为一种加氧酶,用重组技术构建基因在毕赤酵母中高密度发酵表达,酶占总蛋白量56%,表达量达814.5 mg/L[6]。将酶突变基因重组质粒引入E. coli扩增转入酿酒酵母,建立突变文库,突变体 A1773酶活提高5倍。突变体A1242耐高温性提高了3.5倍。突变体DS1474酶比活力56 U/mg,突变体DS896酶比活力为44 U/mg[9],已获准在饲料中作为除毒的酶产品。
烟曲霉真菌感染是困扰人类健康的难题,系统研究烟曲霉基因组中50多个糖基化途径相关基因,先后从烟曲霉中克隆了几丁质酶、磷酸甘露糖异构酶、O-甘露糖转移酶、α-1,4-N-乙酰氨基葡萄糖转移酶、α-葡萄糖苷酶I及CMP-唾液酸合成酶等基因、对了解真菌糖基化机制、开发基因工程药物、抗真菌感染药物有意义[6]。
L-天冬酰胺酶有明确治疗白血病的效果,通过DNA重组技术,将L-天冬酰胺酶特异单链抗体片段与酶构建融合蛋白,提高了酶在体内的稳定性,重组E. coli AS 1357工程酶活力提高达228 U/mL,相当野生菌的50多倍[4]。
用宏基因组学法将酯酶基因转入E. coli中,构建的工程菌提高了活力,表达量为200 mg/L,酶活力达180 U/mg,与野生型比较60 ℃热稳定性提高144倍和196倍,获得了可降解氯氰菊酯、溴氰菊酯、氰戊菊酯和氟氯氰菊酯的新型聚酯类农药降解突变酯酶[9]。
重组毕赤酵母发酵生产碱性果胶酶,1 t发酵罐产酶最高达1 315 U/mL[8]。黑曲霉EIM-6培养优化后果胶酶活力可提高至30 231 U/mL[8,11]。
假单胞菌水杨酸羟化酶基因克隆到 E. coli表达了两个萘降解酶,可降解水杨酸、乙酰水杨酸、磺基水杨酸、水杨醛、间硝基苯甲醛、5-氯水杨酸、辛醛及邻硝基苯甲醛等[5]。
有机磷酯类是一类高毒化合物,用于杀虫剂、除草剂或作为神经毒剂。克隆了假单胞菌有机磷酯水解酶,通过氨基酸位点突变,在E. coli中重组表达,对有机磷酯水解能力提高了约7 000倍[10]。
α-淀粉酶和葡萄糖淀粉酶基因转入酿酒酵母中获得能分解淀粉和生产酒精的 6株酵母工程菌——JL1082工程菌经固定化包埋,葡萄糖淀粉酶活力最高达170 U/mL,淀粉水解率80%以上,产酒率可达13.3%,在广西酒精厂年产5 000 t,工程菌AD (YIP19RGAn) 白酒实验,酒精度为43%,大米利用率为96.16%[5]。
将合成β-聚羟基丁酸酯 (PHB) 的酶基因,λ-噬菌体裂解细胞的酶基因及能合成血红蛋白VHb的酶基因——合成透明颤菌血红蛋白酶基因Vgb,共同转入E. coli中,发酵产聚β-羟基丁酯(可降解的高分子) 工程菌PHB产量达194 g/L,占细胞干重高达 90%,同时解决了破壁及高密度培养的生化工程问题,已进行投产[5]。
抗体酶,又叫催化抗体,具有抗体高选择性和酶的高效催化性。谷胱甘肽过氧化物酶(GPX) 可以清除体内活性氧 (ROS) 所带来的细胞损伤,治疗炎症、糖尿病和心血管病等。根据Pauling过渡态理论,设计了多种含硒的谷胱甘肽过氧化物酶模拟物,通过杂交瘤技术免疫小鼠,化学诱变成功制备了分子量小能穿透细胞膜有治疗价值的具GPX活力的单链抗体模拟物,为天然兔肝酶活力的8.5倍。用人源基因替换鼠非保守区,全基因片段在噬菌体展示,构建了人源化GPX单链抗体酶,还制备了含硒多肽GPX模拟物,活力为68.7 U/mg,为天然酶的80%,最适pH和温度与天然酶相近。通过鼠表皮细胞抗氧化实验,具强抗氧化能力。用化学修饰将θ型谷胱甘肽硫转移酶 (GST) 转化为含硒GPX酶,能有效还原H2O2,催化能力为国外临床试验的PZ51 (含硒杂环化合物) 的5 000倍[5]。通过分子印迹技术,变性卵清蛋白作为印迹分子,除去印迹分子后,印迹分子结合部位中Ser被Sec取代,该蛋白分子有高的GPX活力,为PZ51的820倍[5]。还合成一系列环糊精衍生物的有机硒化合物作为GPX模拟物,对膜通透性好,无免疫原,体内半衰期长[5]。GPX模拟物6-ImTeCD催化谷胱甘肽还原 H2O2活力为6.8 U/μmol,还原t-BuOOH级CuOOH活力是9.7和 15.2 U/μmol[10]。
核酶 (Ribozyme) 是小分子 RNA,也是一种多功能催化剂,可催化病毒RNA自我切割或断裂反应。我国亦进行了[R-]锤头核酶基因防治马铃薯纺锤病毒 (PSTVd) 和[R]锤头核酶基因防治椰菜花叶病毒 (CaMy) 的工作[1]。
此外,还有利用基因工程菌丝氨酸羧甲基转移酶生产 L-丝氨酸[4]。重组毕赤酵母生产腺苷甲硫氨酸药物[5]。重组菌生产 1,3-丙二醇及2,3-丁二醇[8]。构建 E. coli工程菌生产色氨酸[8]。芽短梗霉工程菌生产聚苹果酸[10]。芽胞杆菌突变株Y89D生产环糊精[8]。重组E. coli合成抗禽流感药物达菲前体草莽酸[10]。枯草芽胞杆菌突变株生产苯丙氨酸。利用菌株基因敲除提高工程菌生产化工产品原料丁二酸产量[10]。大肠杆菌工程菌全细胞转化L-苯丙氨酸生产α-苯丙酮酸[10]。
酶工程会议还有许多酶基因克隆与表达的论文,不能一一叙述,如:修复 DNA损伤的O6-甲基鸟嘌呤 DNA甲基转移酶,甜菜碱醛脱氢酶提高抗氧化酶活力,与噬菌体蛋白同源的新核酸酶。甜蛋白基因的表达,蓝藻蛋白裂合酶,抗虫胆固醇氧化酶,嘧啶核苷磷酸化酶[4],乙醇氧化酶,合成 D-氨基酸的 N-氨甲酰-D-氨基酸酰胺水解酶[5],辣根过氧化物酶[7],烟酸脱氢酶[8],乳糖酶[7],甲烷单加氧酶,苯乙烯单加氧酶,右旋糖酐蔗糖酶[8],脂肪醛去甲酰加氧酶,多聚乙酰合成酶,CO2关键酶与限速酶,耐热羧酸酯酶,硫氧化还蛋白谷胱甘肽还原酶[10],烟酸脱氢酶[7],毕赤酵母SASA冠状病毒的表达,癌转移有关的人基质金属蛋白酶,重组毕赤酵母表达人血清白蛋白,耐热单加氧酶,合成 r-亚麻酸的Δ6-脱氢酶在酵母中表达,E. coli表达1,3-丙二醇氧化还原酶生产多聚纤维单体1,3-丙二醇[5],壳聚糖酶,酰胺酶,甘油脱水酶,转谷氨酰酶,丙烯腈水解酶,嗜热菌角质酶-CBD融合蛋白[11],人细胞色素P450酶在E. coli中表达,耐碱性葡萄糖脱氢酶在E. coli中表达,丙氨酸消旋酶[8],果胶内切水解酶,手性药物合成的NADPH酮基还原酶,酪氨酸激酶,糖基合成酶,天冬酰胺合成酶,S-腺苷同型半胱氨酸水解酶,丙酮酸脱氢酶,无机焦磷酸酯酶[6],E. coli表达治疗肝癌与黑色素瘤的精氨酸脱亚胺酶。制备手性醇的酮还原酶。热稳定制备芳基醇的醇脱氢酶[9]。异源表达β-1,3-氮乙酰胺基葡萄糖转移酶制备人乳寡糖,克隆了 5种糖核苷酸合成的关键酶合成相应的糖核苷酸,E. coli表达细菌脂肪氧合酶用于绿色香料和植物激素合成。E. coli诱导表达对治疗癌症及高半胱氨酸症有价值的深海甲硫氨酸r-裂解酶。E. coli表达Bst DNA聚合酶[9]。抗生素骨架聚酮合成的迭代聚酮还原酶。肿瘤标记物环氧化酶-2的人源单链抗体。E. coli表达双功能D-乳酸脱氢酶,重组表达系列琼胶酶制备琼胶寡糖。里氏木霉异源表达木质素过氧化物酶[10],构建了 α-乙酰乳酸脱羧酶啤酒酵母工程菌用于啤酒生产[4]。聚酮合酶是最强大的化学合成酶[10],正在进行结构功能研究。
2 酶与微生物细胞的生物合成与催化
传统的化学催化已不能满足可持续发展的需要,高立体选择性的微生物细胞或酶已成为手性生物合成的亮点。在这25年中,我国工业生物催化已成为具有相当规模和技术水平的产业,在世界上占有举足轻重的地位。
在第一至九届全国酶工程会议的 978篇论文中,基因工程酶方面的有 356篇,生物催化合成论文有 186篇,占第二位。其中脂肪酶催化合成论文占55篇,占生物合成的第一位。
我国在有机相、离子液体、非水溶剂系统已成功利用脂肪酶进行了手性 2-辛醇[3,5,7-8]拆分,(R,S) 仲辛醇拆分[3,5-6],α-苯乙醇拆分[6],酮洛芬乙烯水解拆分[10],合成己酸乙酯[3-4,7,11],合成辛酸乙酯[7],油酸乙酯[7],庚酸乙酯[10],癸酸乙酯[7],生物柴油[7-9,11],脂肪酸甲酯[11],二元酸酯[8],单甘油酯[4],油酸油醇酯[3],香叶醇酯[3],油酸酯[3],乙酸乙烯酯[3],脂肪酸糖脂[9],(S)-α-氨基-3-苯氧基苄醇[5],1,3-丙二醇羧酸酯[3],棕榈异辛酯[8],癸酸甘油酯[5],维生素A棕榈酸酯[5],抗坏血酸脂肪酸酯[5],蔗糖酯[7],聚羟基丙酸酯[7],葡萄糖月桂酸单酯[8,11],果糖月桂酸单酯[8],及 (+)反式菊酸[4],以及新闻纸脱墨[7,10]等。
细胞展示技术是将外源蛋白锚定在细胞壁上,酶分子的固定化可提高酶的稳定性和对有机溶剂的耐受性,毕赤酵母对外源脂肪酶表面展示系统可催化合成果糖脂、己酸乙酯,葡萄糖酯、生物柴油、月桂酸单糖酯、脂肪酸甲酯[8,11]等。
离子液体体系是新型绿色非水溶剂,酶法微藻藻油生产生物柴油达 90%转化率。并合成了脂肪酸糖脂[9-10]。
利用酯酶催化合成酒香己酸乙酯[3],单一性2-辛醇拆分合成抗癌药物前体 (2S,3R)-3-苯基缩水甘油酸甲酯[10],固定化合成脂肪族聚酯[10],菌株不对称合成L-薄荷醇[7],拆分1,1,1-三氟-2-辛醇[10],有机合成壬酸香草醇酯 (辣椒素脂)[8]。
用蛋白酶与化学法相结合,在低水有机相中合成了 4种含 Arg-Gly-Asp(RGD) 的细胞粘附肽 RGDS、RGD(NH2)2、RGE-NH2和RGDS-NH2,有抗肿瘤作用[7]。
目前,已知酶有4 428种 (至2011年),工业上用酶大多数为水解酶,但氧化还原酶在酶中比例却约占 32%,很多氧化还原酶未能被开发,因此拓展氧化还原酶是工业技术中一项重要内容,但需解决辅酶循环再生的难题。为建立微生物细胞氧化还原酶催化体系,提高内源辅酶利用率,我国已利用E. coli重组菌双辅酶底物偶联固定化面包酵母羰基还原酶合成 (S)-苯基乙二醇手性模块物[8]。用双羰基还原酶偶联辅酶再生体系合成他汀类药物手性侧链——3R,5S二羟基-6-苄基己酸乙酯[8]。重组菌羰基还原酶合成医药合成用的手性醇 (S)-4-氯-3-羟基丁酸乙酯,(R)-邻氯扁桃酸甲酯和 (S)-2-羟基-4-苯基丁酸乙酯[9]。用烯酮/烯酯还原酶还原C=C双键合成有商品价值的底物,对甲基马来酰亚胺、R-香芹酮、衣康酸二甲酯、乙烯酰氧基丙烯酸甲酯、茶香酮、苄烯丙二腈都有较高的转化率[10]。用辅酶再生体系酵母醇脱氢酶不对称合成医药,农药光学手性醇中间体 (R)-扁桃体酸甲酯和 (R),(S)2-辛醇[8]。菌株 D-氨基酸脱氢酶和胺氧化酶合成手性胺 (如 D-丙氨酸、D-苯丙氨酸、D-叔亮氨酸)。已建立了 D-氨基酸合成的酶催化平台[10]。氧化葡萄糖酸杆菌高选择性氧化合成工业上第二位的芳香醛——苯甲醛[8]。嗜热酮还原酶辅酶再生系统不对称合成医药中药中间体S-1-苯基222三氟乙醇[9]。E. coli共表达全细胞甲酸脱氢酶催化茴香硫醚不对称合成手性亚砜——甲基苯基亚砜[9]。金黄杆菌烯醇还原酶催化C=C双键不对称还原环状烯酮产生光学纯烷烃化合物用于手性中间体合成[10]。金黄杆菌短链脱氢酶不对称合成匹瑞匹坦药物中间体(R)-[3,5-二 (三氟甲基) 苯基]乙醇[10]。芽胞乳杆菌D-乳酸脱氢酶催化谷氨酸合成α-酮戊二酸[10]。高活性全细胞醇脱氢酶不添加辅酶合成米格列醇[6],基因敲除提高酶表达量构建甘油脱氢酶合成1,3-二羟基丙酮 (最简单的多羟基酮糖)[8]。利用天然多脱氢酶体系甲烷氧化细菌生物催化二氧化碳制甲醇[5]。厦门大学挖掘了海洋微生物的8种氧化还原酶体系[9]。
国内已形成生物催化与微生物转化的科研开发群体,江南大学与浙江鑫富生化公司合作,用产 D-泛解酸内酯水解酶的串珠镰孢菌水解拆分得到光学纯的D-泛解酸内酯,成功地用于D-泛酸钙和D-泛醇的生产,D-泛酸钙产量全球第一[5,10]。此外,还固定化诺卡氏菌水解顺式环氧琥珀酸生产 L-酒石酸[7]。微生物转化生产香草酸与香草醛,又名香兰素 (3-甲氧基-4-羟基苯甲醛) 高档香料[5]。转化巴豆甜菜碱制L-肉碱[4],采用双水相合成用于倍他司汀抗组胺药物手性中间体-(S)-(4-氯苯基)-(吡喃-2-基) 甲醇[9]。定向进化E. coli菌株精氨酸脱亚胺酶为治肝癌药物[9]。
融合酶是利用融合蛋白将几种功能蛋白集成于一体的嵌合体。构建肝素酶的双重融合蛋白体系,如麦芽糖结合蛋白融合或荧光蛋白融合,促进了肝素酶的可溶性表达,用于生产低分子量抗栓药物——肝素[6-7,9,12]。双亲短肽融合环糊精糖基转移酶的N末端构建6种融合酶合成克服Vc氧化的2-氧-D-吡喃型葡萄糖基-L-抗坏血酸[10]。固定化自组装短肽18A与腈水解酶融合表达生产扁桃腈[10]。用嗜热子囊菌角质酶-CBD融合蛋白精炼棉织物[8]。
大孔树脂吸附法具有吸附容量大、选择性好、吸附速度快、时间短、物理化学稳定性好等优点,但其也有一定的局限性。树脂经过反复使用后,树脂的表面及内部会残留许多非吸附性成分或者是杂质使柱的颜色变深,柱效也会降低,而且经多次使用有时柱床挤压过紧还会导致树脂颗粒破碎,使用寿命较短。此外由于即使是同一生产企业生产的同一型号树脂,各批之间的比表面积和功能基团的含量差别也较大,导致实验的重现性较差,影响实验结果的准确性。
我国在 20世纪90年代就已开发利用过氧化物酶在有机相或反相胶束中催化酚类和芳香胺的聚合反应,如合成聚对羟联苯,并研究了聚合物的光电性能。在非水介质中催化去除苯酚达84.9%。在水乙醇中前列腺素合成酶催化合成前列腺素 E1[3]。青霉菌转化顺丙烯磷酸一步环氧化为磷霉素[4]。环氧化物水解酶合成药物手性中间体手性邻二醇[7]。浙江大学与海正药业公司合作克隆表达 R-酰胺酶和水合酶合成西司他汀手性中心 (S)2,2-二甲基环丙甲酰胺[8]。利用休止细胞甘油脱水酶生产聚酯、聚醚、聚氨酯合成的单体1,3丙二醇[5]。构建E. coli基因工程菌转化甘油生产3-羟基丙醛[8]。利用羟腈酶合成可制备β-氨基醇的羟腈化合物[4]。利用嘧啶核苷磷酸化酶合成抗肿瘤药物中间体-5-氟尿苷[4]。用反胶束单宁酶催化合成抗氧化剂没食子酸戊酯[5]。成都生物所用固定化粘红酵母苯丙氨酸解氨酶/肉桂酸途径生产苯丙氨酸[7]。腈转化酶是继酯酶和脂肪酶之后,又一制备手性化合物的工具酶,包括腈水解酶、腈水合酶、酰胺酶。产业化实例有腈水合酶/酰胺酶双酶偶联生产西司他汀钠关键手性中间体-(S)-(+)2,2-二甲基丙烷甲酰胺[8]。腈水解酶制备除草剂-(L)-单铵膦及农药甘膦中间体——亚氨基二乙酸[10]。用固定化细胞腈水解酶制备医药中间体-(R)扁桃酸[8]。非水相α-胰凝乳蛋白酶催化合成具有强抗氧化的类肌肽4-(5)-丙氨酰胺5 (4)-羧酸咪唑[8]。基因工程菌转化制备脱氧核苷三磷酸[8]。定向突变菌株环氧化物水解酶制备降压药普萘洛尔中间体邻位二醇[10]。甾体激素仅次于抗生素,用青霉菌转化 6种甾体生成 1,2-二氢睾内酯[10]。手性胺或非天然氨基酸是重要药物合成的手性模块,用 ω-转氨酶获得光学纯的 (R)手性胺 (R)-α-苯乙胺,(R)-1-苯丙胺,(R)-4-苯基-2-丁胺,(R)-α-四氢萘胺[10]。E.coli表达细胞 L-氨基酸脱氨酶催化合成 α-苯丙酮酸[10]。固定化细胞转化精氨酸为瓜氨酸,通过细胞谷氨酸氧化酶转化谷氨酸制备 α-酮戊二酸[10]。固定化含精氨酸脱亚氨酶的细胞拆分DL-精氨酸[9,12]。用内切型肝素酶制备抗平滑肌细胞增生的肝素寡糖 (0–8 K)[8]。突变菌株环糊精葡萄糖基转移酶合成6个葡萄糖的α-环糊精[8]。用4种酶法合成新生儿益生糖——氮乙酰氨基葡萄糖-1,3-乳糖[9]。嗜热β-葡萄糖苷酶在两相中合成非离子表面活性剂——辛基葡萄糖苷[9]。琼胶酶降解琼胶生成2、4、6、8和10糖[10]。右旋糖酐酶水解葡聚糖生成麦芽糖,异麦芽三糖和葡萄糖[10]。
新发现的人参皂苷糖苷酶,从人参二醇类皂苷酶转化法获得稀有皂苷Rh2,其得率比红参中提取提高了 500–700倍,在大连已产业化,年产Rh2皂苷300 kg,证明酶转化法可以生产中草药的有效成分[5,11]。β-糖苷酶水解人参二醇皂苷Rb1和Rb2的糖基生成抗癌人参皂苷Compound K。可提高免疫力,增加白血球等药效[10]。
硅基磁性纳米材料 Fe3O4SiO2 (Hb) 肌红蛋白具有过氧化物酶活性[9]。重组菌不对称催化合成手性环氧化物和手性邻位二醇产物 R-对氯苯二醇和R-对氯苯乙二醇[10]。聚酮合酶是自然界最小,但功能最强大的合成工厂,催化合成抗生素次级产物,正在进行结构域研究[5,10]。
长链二元酸是10个以上碳原子的脂肪族二羧酸,是化工合成高性能尼龙、麝香香料、润滑油、油漆、粘合剂、新型医药和农药的精细化工原料,长链二元酸的生物合成是中国独有的原创性新菌种、新工艺专利技术,已经大规模工业化生产,只有中国能应用这种技术生物合成生产一系列长链二元酸,并垄断了西方所没有的 7种长链二元酸,每年产值有十几亿人民币,荣获多项国家重大成果奖[4]。
3 生物传感器
生物传感器应用于各类重要生化物质快速分析,我国已研制成功 101种以上不同种类、台套的生物传感器,包括电极型、化学光敏型、场效应晶体管型、等离子体谐振型、光波导型、光纤型、压电晶体型、微悬臂列阵型、双分子质脂仿生型、椭扁光型、丝网印刷型,大多数采用酶膜、组织、微生物、受体蛋白、抗原抗体、叶绿体、DNA探针作为探头,测定物质为乳酸、谷氨酸、血糖、尿素氨、尿酸、肾功能、青霉素、次黄嘌呤、谷氨酰胺、维生素C、转氨酶、有机磷、BOD、丙酮酸、黄曲霉毒素、海洛因、酚、苯丙氨酸、胆固醇、细菌总数、乙醇、抗原抗体、嗅觉、硫化物、微生物、环境监测、二氧化碳、钾离子、胆碱、β-羟丁酸、农药、DNA等。十个研究所组成的中国科学院国家传感联合开放实验室及全国院校做出了许多成果,微型血糖仪已商品化,山东省科学院生物研究所制备了一系列 SBA-30乳酸分析仪、SBA-40谷氨酸葡萄糖双功能分析仪、SBA单电极分析仪、SBA60四电极在线分析仪、SBA70血糖乳酸自动分析仪及可检测糖化酶、鱼虾鲜度、转氨酶、尿素氨、维生素C的生物传感器,已经形成了商品化,实际应用于发酵生产和实验室的实用检验。最近,还研制了NADH生物传感器[6]。
经过多年的科研积累,我国酶工程研究已形成了许多有特色的科研队伍和领路人,建立了许多技术平台,中国科学院在北京、天津、上海建立了酶工程共性的技术平台,中国农业科学院构建了饲料酶技术开发平台,北京化工大学建立了脂肪酶化学品合成技术平台,华东理工大学建立了脂肪酶工业应用平台,福州师范大学构建了微生物脂肪酶产业化技术平台,中国药科大学建立了氧化还原酶研发技术平台,江南大学建立了氧化还原催化体系技术平台,厦门大学建立了氧化还原酶检测平台,清华大学建立了生化工程技术平台,华南理工大学构建了酵母细胞表面展示技术平台。山东大学建立了微生物工业技术平台,合肥学院建立了生物与环境技术平台,吉林大学建立了分子酶学与进化技术平台。
我国酶工程研究开发团队做出了许多突出的成果与贡献,获得了多项国家奖励。荣获国家科学技术发明二等奖的有:姚斌研究员、孙志浩教授、郑裕国教授、陈坚教授、徐岩教授、杨立荣教授、谭天伟院士、陈国强教授。荣获国家科学技术进步二等奖的有:黄日波教授、金凤燮教授、曲音波教授、陈远童研究员。此外,高培基教授荣获教育部科学技术进步一等奖。在会议上,荣获酶工程贡献奖的有:孙志浩教授、金凤燮教授、黄日波教授、许建和教授、冯雁教授。曾先后参加过酶工程会议的院士有:邹承鲁院士、张树政院士、范云六院士、欧阳平凯院士、杨胜利院士、沈寅初院士、赵国屏院士、王志珍院士、田波院士和谭天伟院士。酶工程已成为我国十分重视的现代生物技术。
4 我国酶制剂工业发展现状
据中国生物发酵产业协会统计,2014年,我国酶制剂商品产量21.9万t,产值28亿元人民币 (不包括外资企业)。
我国酶制剂工业起步于 20世纪 60年代,但真正得到发展是在 20世纪 80年代初改革开放后,特别是进入新世纪初,企业由地方国营改制为股份制或民营企业。改制后企业推行 GMP管理,引进人才,采用新工艺新设备,提高产品质量,降低生产成本,并与大专院校、科研院所合作或委托开发新产品。扩大应用领域,使酶制剂工业快速发展。
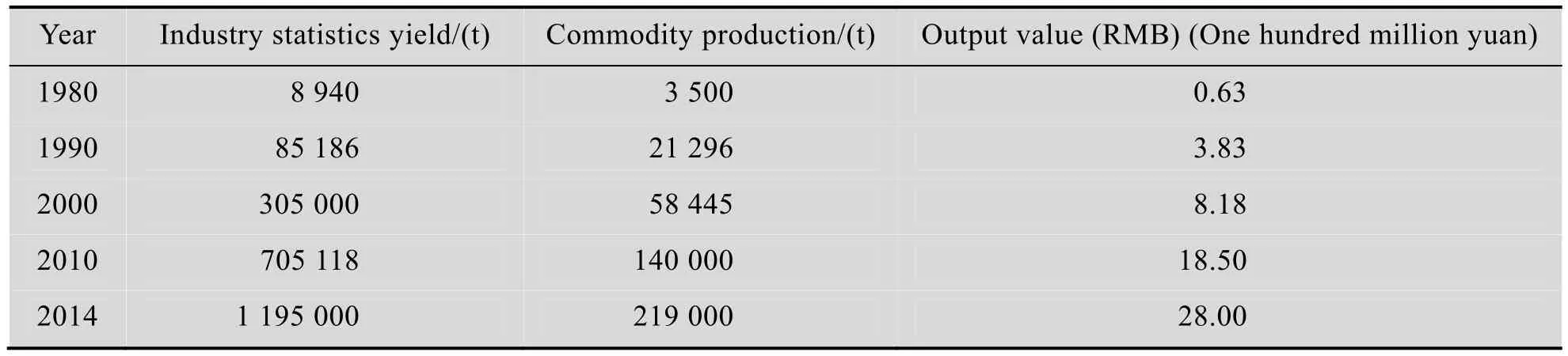
表2 我国酶制剂产量、产值 (不包括外资企业)[13]Table 2 The production and the output value of enzyme preparation production in China (not including foreign capital enterprises)
近10年,在市场激烈竞争中,行业中一些规模很小又无特点的企业逐渐边缘化,有的被淘汰出局。同时,出现了一些规模较大具有发展潜力的企业,引人关注。如:山东隆大生物工程有限公司[14],这家企业目前已拥有发酵总容积3 000 m3,生产高温α-淀粉酶 (普通、酸性)、中温 α-淀粉酶、真菌淀粉酶、β-淀粉酶、糖化酶、普鲁兰酶、复合糖化酶、纤维素酶 (酸性、中性)、木聚糖酶、β-葡聚糖酶、蛋白酶 (酸性、中性、碱性)、风味蛋白酶、过氧化氢酶、葡萄糖氧化酶、果胶酶 (酸性、碱性) 近20种。应用于多个行业,并有较大出口量,是行业中生产品种最多的企业。2014年产值5亿元人民币。该企业2014年还投入专项资金1 000多万元用于完善治理生产中产生污染环境的三废(废水、废渣、废气)。广东溢多利生物科技有限公司[14]是行业中最早生产饲料酶的企业,除在珠海建有国内最大的固体发酵生产基地,并在内蒙托克托建有液体发酵生产线,生产多种产品。近年来产品应用已由饲料延伸至食品、纺织、造纸等领域。2014年产值4亿元人民币。该企业已成功上市。这两家企业,加上湖北新华扬生物股份有限公司、湖南鸿鹰祥生物工程股份有限公司、湖南尤特尔生化有限公司、北京挑战饲料科技集团、山东康地恩生物科技有限公司、北京昕大洋科技发展有限公司、江苏博立生物制品有限公司,共9家企业,2014年总产值24.5亿元,占全国酶制剂企业当年产值28亿元的87.5%。企业发展规模化已成趋势。
根据海关统计:2014年进口酶产品14 578 462 kg,金额为2.327亿美元,出口酶产品86 593 934 kg,金额3.369亿美元[15]。
上述酶制剂产品进出口包括国内生产企业、在我国独资生产的诺维信、杰能科、帝斯曼、天野等公司、合资企业苏州宏达酶制剂有限公司以及在我国设办事处做贸易的一些国外公司。我国酶制剂生产企业出口酶制剂金额约占我国酶制剂出口总金额的1/3。
酶制剂出口产品中,国内企业出口的品种依次为:糖化酶、植酸酶、碱性蛋白酶、碱性脂肪酶、高温α-淀粉酶、纤维素酶、木聚糖酶、果胶酶等。糖化酶数量最多,年出口约3.0万t,其次为植酸酶,年出口约1.0万t。
植酸酶:植酸酶研发项目 (包括禽类、鱼类、猪等专用植酸酶),由中国农业科学院饲料研究所姚斌研究员负责的课题组承担[13],列入国家“863”计划,于1998年取得重大突破,实现工业化生产,是用基因工程菌毕赤酵母生产。2005年研究又取得重大进展,开发出新一代高比活植酸酶,处于国际领先地位。2010年,该产品生产销售超过15 000 t,产值超过3亿元人民币,占据国内植酸酶市场 90%以上。据我国饲料行业统计,由于饲料中添加了植酸酶,年节约饲料原料磷酸氢钙20万t以上,节约饲料成本约9亿元,并使动物粪便中排出的可污染环境的磷减少了约30万t,创造了较大的经济效益和生态效益。主要生产企业:北京挑战饲料科技集团、湖北新华扬、广东溢多利、四川禾本生物等。
纤维素酶:主要生产企业为湖南尤特尔生化有限公司、山东隆大生物工程有限公司、湖南利尔康生物科技有限公司等。酸性纤维素酶是用基因工程菌里氏木霉生产,由毕业于湖南农业大学的留美李新良博士研制,2001年实现工业化生产,产品的性价比居国内领先地位。
高温 α-淀粉酶 (普通、酸性):主要生产企业为山东隆大生物工程有限公司、江苏博立生物制品有限公司。
普鲁兰酶:该产品可水解液化淀粉中的α-1,6-D糖苷键产生 α-1,4-D直链多聚糖,用于与糖化酶配合生产高含量葡萄糖,与 β-淀粉酶配合生产麦芽糖含量高于 70%的麦芽糖浆主要生产企业:山东隆大生物工程有限公司。
α-乙酰乳酸脱羧酶:该产品可降解双乙酰的前驱物 α-乙酰乳酸,减少双乙酰的生成,确保啤酒中低水平双乙酰含量,对提高啤酒质量有重要作用。该产品研发项目由广西大学生物技术有限试验中心研究[13],课题负责人为留英归国黄日波博士,项目列入国家“863”计划,通过基因工程菌重组构建,发酵工艺优化,酶的提取分离纯化等技术攻关,1999年取得突破,实现工业生产。主要生产企业:南宁邦尔克生物技术有限公司。邦尔克生产的该产品2014年占据国内市场50%以上,并出口欧美和亚洲。
啤酒复合酶:主要生产企业为宁夏夏盛实业集团有限公司[16],该产品于2000年研究成功,投入生产。该产品含有β-葡聚糖酶、木聚糖酶、果胶酶、细菌中性蛋白酶、淀粉酶等。糖化时添加,用量为麦芽干重的0.03%–0.05%。可降低麦汁粘度,提高过滤速度,澄清麦汁,增加收率,提高氨基氮含量,有利酵母生长。
脂肪酶:主要生产企业为深圳市绿微康生物工程有限公司,生产菌种扩展青霉,由福建师范大学生物工程学院吴松刚教授提供。北京凯泰新世纪生物技术有限公司,菌种假丝酵母,由北京化工大学谭天伟院士提供。此外还有百圣龙生物工程有限公司,四川禾本生物技术有限公司和山东隆大生物工程有限公司 (国外引进,菌种黑曲酶)。
截至2014年,我国酶制剂行业生产的产品已有 20多个原酶品种 (糖化酶、植酸酶、α-淀粉酶、蛋白酶、纤维素酶、木聚糖酶、β-葡聚糖酶、果胶酶、脂肪酶、β-淀粉酶、真菌淀粉酶、普鲁兰酶、葡萄糖氧化酶、过氧化氢酶、甘露聚糖酶、半乳糖苷酶、α-乙酰乳酸脱羧酶、乳糖酶、凝乳酶、角质酶等)。五大酶种占酶制剂总产量82%,其中糖化酶为30%、植酸酶为22%、淀粉酶为 14%、蛋白酶为 7%,纤维素酶为6%[17]。产品应用于淀粉糖工业、纺织工业、饲料工业、酒精工业、啤酒工业、果汁果酒工业、面食制品加工、皮革工业、造纸工业等多个领域。
行业存在的主要问题:酶的品种还比较单一,由不同原酶组成具有不同用途的复合酶制剂发展滞后,不能满足市场的需要;一些产品的性价比不高,发酵酶活力较低,有些产品提取工艺和装备水平落后,产品还是粗制品,缺乏市场竞争力;酶制剂生产过程存在三废对环境的污染问题,比如生产1 t液体糖化酶 (100 000 U/mL)可产生COD (化学需氧量) 高达5 000 mg/L以上的废水10 t。酶制剂生产企业必须推行国家清洁生产政策,投入专业资金,做好自身的环境控制。重视这些问题的解决,将推动我国酶制剂工业持续健康发展。
致谢:感谢中国食品科技学会酶制剂分会侯炳炎先生提供我国酶制剂工业发展状况数据。
[1] Enzyme 2. Engineering XIV 1997, Beijing.
[2] 全国首次酶工程学术讨论会论文摘要汇编,1991, 威海.
[3] 吉林大学自然科学学报, 特刊, 第二次全国酶工程学术讨论会论文集, 1993, 长春.
[4] 第三次全国酶工程学术讨论会论文摘要集,2000, 成都.
[5] 第四届中国酶工程学术交流讨论会论文集,2003, 无锡.
[6] 第五届中国酶工程学术研讨会论文集, 2005,海口.
[7] 第六届中国酶工程学术研讨会论文集, 2007,昆明.
[8] 第七届中国酶工程学术研讨会论文摘要集,2009, 合肥.
[9] 第八届中国酶工程学术研讨会论文集, 2011,广州.
[10] 第九届中国酶工程学术研讨会论文摘要集,2013, 南宁.
[11] 生物工程学报, 酶工程专刊, 2009.
[12] 生物工程学报, 酶工程专刊, 2012.
[13] 孟广震. 中国生物工程学会组织. 新中国工业生物技术发展史略. 北京: 化学工业出版社,2013: 47–50.
[14] 侯炳炎. 结缘酶制剂工业四十年. 第四届全国酶制剂研究开发应用技术研讨会论文集. 无锡,2014, 10: 13–14.
[15] 中国生物发酵产业协会主办. 发酵工业, 2015,2: 42–44.
[16] 段钢, 姜锡瑞, 周红伟. 酶制剂应用技术问答.北京: 中国轻工业出版社, 2008: 163.
[17] 石维忱. 持续推进我国酶制剂工业健康发展.无锡第四届全国酶制剂研究开发应用技术研讨会论文集, 2014, 10: 1–2.
猜你喜欢
疯狂英语·新读写(2021年10期)2021-12-07
当代陕西(2020年9期)2020-08-04
中华诗词(2020年8期)2020-02-06
奥秘(2019年8期)2019-08-28
中国化妆品(2018年6期)2018-07-09
商周刊(2017年7期)2017-08-22
小猕猴智力画刊(2016年6期)2016-05-14
中国工程咨询(2016年6期)2016-01-31
中国卫生(2015年7期)2015-11-08
浙江人大(2014年1期)2014-03-20
