
白鹤幼鹤行为发育
2015-02-05 01:23:38钱法文
生态学报 2015年21期
张 琼,钱法文
中国林业科学研究院森林生态环境与保护研究所, 北京 100091
白鹤幼鹤行为发育
张 琼,钱法文*
中国林业科学研究院森林生态环境与保护研究所, 北京 100091
2012年10月至2013年5月,采用焦点取样法对在吉林莫莫格国家级自然保护区内秋季和春季迁徙停歇的以及在江西鄱阳湖国家级自然保护区内越冬的白鹤(Grusleucogeranus)幼鹤行为进行了研究。结果表明,幼鹤取食行为比例从秋季迁徙时的23.0%增加到越冬晚期时的82.7%,但春季迁徙期减少为61.9%;幼鹤乞食行为比例从秋季迁徙时的58.2%减少至春季迁徙时的1.2%;幼鹤警戒行为比例从秋季迁徙时的1.0%增加至春季迁徙时的7.1%。幼鹤在春季迁徙期的理羽及静栖行为显著高于越冬期,这种行为的差异可能与食物的丰富度相关。环境因子中,风速对幼鹤静栖行为有显著影响,幼鹤取食及警戒行为与气温呈显著正相关。秋季迁徙及越冬晚期,幼鹤行为日节律在13:00—14:00 时出现明显的取食低谷期,而春季迁徙期时则推后1小时。整个观察期间,幼鹤从雄鹤和雌鹤获得食物频次无显著差异,但不同时间段幼鹤从雄、雌成鹤处获得食物频次差异是显著的。研究从行为学的角度展示白鹤幼鹤的生长过程及不同性别成鹤对幼鹤生长的贡献。
白鹤幼鹤;行为;生长;取食;栖息地
白鹤(Grusleucogeranus)属鹤形目鹤科鹤属鸟类,为大型涉禽。白鹤是一种迁徙性鸟类,主要分布在亚洲大陆,目前全世界仅有3 000余只[1]。被IUCN红皮书列为极度濒危物种[2],中国濒危物种红皮书列为濒危级物种[3], 为我国国家Ⅰ级重点保护野生动物。现存的白鹤可分为东部和西部2个种群。东部种群为数量最大的种群,在俄罗斯西伯利亚东北部繁殖,在中国长江中游流域越冬,主要越冬地为江西的鄱阳湖[1]。近年来野外调查结果显示,在鄱阳湖区越冬白鹤种群数量占世界种群数量的近99%[4- 5]。黑龙江省的扎龙湿地、吉林省的莫莫格湿地、内蒙古自治区的图牧吉湿地、辽宁省的獾子洞湿地和盘锦湿地以及渤海湾湿地等是白鹤东部种群迁徙路线上的重要中途停歇地[6- 11]。莫莫格湿地为白鹤迁徙途中最重要的中途停歇地[12- 13],在此迁徙停歇的白鹤数量记录达到3800余只,迁徙期停留持续长达60余天。
白鹤主要在浅水区域活动,以植物性食物为主[14- 17]。常见的白鹤家庭构成为2成1幼或2成[18]。当年的幼鹤由于其身体和生理的不成熟及行为模式的不稳定,自身获得的食物的能力和数量有限,必须从其父母处获得食物来补充营养物质[16,19]。Sauey在印度克拉迪奥国家公园对越冬白鹤幼鹤行为研究显示,白鹤幼鹤的乞食占其所有用于获得食物时间的22.5%,且从雄性成鹤获得食物量是从雌性成鹤获得食物量的2.5倍[14]。
目前对白鹤行为研究主要集中在研究单一时间段白鹤的个体行为[20- 21],未见从行为变化方面来研究白鹤幼鹤的生长过程。为了掌握白鹤幼鹤生长过程中的行为变化及其生长轨迹,本研究从以家庭为单位的白鹤日活动规律入手,展开对白鹤幼鹤生长过程行为变化的研究。
1 研究地概况
吉林莫莫格国家级自然保护区(简称“莫莫格保护区”)位于吉林省镇赉县东南部, 东与黑龙江省杜尔伯特、泰来县隔嫩江相望;南以洮儿河为界, 与吉林省大安市相邻。地理坐标为:122°27′—1244′E,45°45′—46°10′N。全区总面积1440 km2,其中湿地面积占80%,是典型的湿地类型保护区,为鹤类等珍稀水鸟的栖息繁殖提供了良好的生存环境。莫莫格保护区属温带大陆季风气候, 春旱风大、夏季多雨、秋季冷爽、冬季雪少。年均气温4.4℃,1月均温-17.5℃,7月均温23.5℃。年均降雨量377.1 mm, 无霜期137 d。
江西鄱阳湖国家级自然保护区(以下简称“鄱阳湖保护区”)位于江西省北部,中国第一大淡水湖鄱阳湖的西北角,其地理坐标为:115°55′—116°03′E,29°05′—29°15′N。鄱阳湖保护区总面积为224 km2,包括有大湖池、沙湖、蚌湖、大汊湖等9个湖泊,是生物多样性十分丰富的国际重要湿地,是白鹤东部种群的主要越冬地[4,22]。鄱阳湖保护区属亚热带湿润性季风型气候,气候温暖,四季分明,无霜期长约270d。年均气温为17.1℃,7月份气温最高,平均为29.1℃;1月份气温最低,平均4.5℃。鄱阳湖是季节性过水湖泊,湖水位受到入湖五河来水和长江水位双重影响,呈现出水陆交替的特征;通常鄱阳湖在4—9月间在丰水季节的面积约3000 km2,而10月至翌年3月枯水季节的面积仅为500 km2,露出大部分的泥滩及浅水区域,为鹤类等水鸟提供良好的越冬栖息地[17,23]。
2 研究方法
2012年秋季的10月至11月初和2013年春季的4月至5月,以及2012年冬季的11月下旬至12月和2013年1月至3月,分别对在莫莫格保护区秋、春季迁徙的和在鄱阳湖保护区越冬的白鹤家庭行为,采用焦点取样法进行了观察。用10倍双筒望远镜(Eagle Optics 10×50)随机选取离其他白鹤群超过20 m的2成1幼白鹤家庭,采用20—60倍单筒望远镜(Kowa 20—60×82)进行观察并记录每个个体的行为。白昼观察时间一般为8:00—17:00 时,在无干扰情况下,1天固定观察1个白鹤家庭。雨天停止观察。
白鹤秋季迁徙时一般于9月下旬到达莫莫格保护区,11月上旬离开莫莫格迁往越冬地;越冬白鹤于11月上旬至中旬到达鄱阳湖区,次年3月底至4月上旬离开越冬地开始春季迁徙;春季迁徙的白鹤于4月中旬到达莫莫格保护区,5月中旬离开莫莫格保护区迁往繁殖地[7- 9,24]。根据幼鹤的生长阶段及观察时间,将整个观察期间分为5个时间段:2012年10月至11月初为秋季迁徙期,总计观察7 d;2012年11月下旬至12月5日为越冬早期,总计观察8 d;2012年12月8日至12月末为越冬中期,总计观察9 d;2013年1月和3月为越冬晚期,总计观察16 d;2013年4月中旬至5月中旬为春季迁徙期,总计观察13 d。从当地气象站获得观察区域的气温及风速数据,将风速分为3个等级:等级1为风速小于3级,等级2为风速3—4级,等级3为风速大于4级。白鹤主要以集大群或者家庭群方式活动,本次研究对象为以2成1幼家庭群的活动行为。
根据行为分类,将白鹤幼鹤行为分为取食、乞食、警戒、游走、理羽、静栖和其他:
取食 将头插入水中寻找食物,在水中清洗食物及吞咽过程,在草地或水边觅食。
乞食 跟着其父母身边等待、鸣叫或从成鹤嘴中抢食。
警戒 抬头、伸直脖子四处张望。
游走 行走、跑或短距离飞行。
修饰 包括啄理羽毛、在水中清洗头颈部、用爪抓挠头颈部、用喙啄脚、单腿站立并抬头、蓬羽等。
静栖 单腿站立将头埋在翅下不动,跪或卧倒在旱地上。
其他 包括领域行为,如鸣叫、驱赶入侵的其他白鹤,及原地扇翅、跳跃,非警戒行为的站立等。
野外观察收集到白鹤幼鹤活动时间共计420 h。用Excel 2003软件对观察数据进行初步整理,数据分析采用SPSS 20.0统计分析软件来完成。
3 结果
3.1 白鹤幼鹤行为的变化趋势
统计幼鹤各个时间段的活动时间比例(表1),可以发现随着幼鹤的生长,其各行为比例都出现明显变化。在越冬期,幼鹤各行为比例呈递增或减的趋势,而秋季及春季迁徙期与越冬期对比,并不随时间增加或减少。通过单因素方差分析,不同时间段幼鹤的各行为比例差异极显著。

表1 不同时间段白鹤幼鹤各行为比例分配(Mean±SD)及单因素方差分析Table 1 Time budget of the behaviors of juvenile Siberian Crane at different periods and its ANOVA (**P<0.01, *P<0.05)
不同时间段幼鹤各行为比例明显变化,通过LSD两两比较发现,幼鹤的取食行为比例从秋季迁徙期至越冬晚期呈递增趋势,但春季迁徙期有所降低;秋季迁徙期与越冬早期差异显著(P=0.025),而其他时间段之间的差异极显著(P<0.001)。幼鹤乞食行为比例随时间递减,越冬晚期与春季迁徙期乞食行为比例无显著差异(P=0.661),其他时间段间差异极显著(P<0.001)。警戒行为比例呈整体递增趋势,在秋季迁徙期、越冬早期和越冬中期与越冬晚期和春季迁徙期间差异极显著(P≤0.001),其他时间段间无显著差异(P>0.05)。游走行为比例先减少再增加,在越冬早期与越冬中、晚期之间存在显著差异(P<0.01),其他时间段间无显著差异(P>0.05)。幼鹤理羽行为在秋季及春季迁徙期显著高于越冬期(P≤0.001),秋季迁徙期与春季迁徙期间也存在显著差异(P=0.023),而整个越冬期幼鹤理羽行为无差异(P>0.05)。静栖行为比例在春季迁徙期显著高于其他时间段(P≤0.003),而除春季迁徙期的其他时间段间无差异(P>0.05)。越冬期的其他行为比例显著高于迁徙期的(P<0.001)。
3.2 不同时间段白鹤幼鹤行为的日节律
取食行为比例在秋季迁徙期、越冬晚期及春季迁徙期出现明显的低谷波动,其他时间段波动较小;秋季迁徙期和越冬晚期在13:00—14:00 时有明显低谷,而春季迁徙期的低谷出现在14:00—15:00 时。乞食行为比例在秋季迁徙期呈从早到晚的微弱递增趋势,越冬早期在14:00 时后明显减少,其他时间段波动很小。警戒行为比例波动很明显,低谷一般出现在10:00—13:00 时;越冬早期及越冬晚期在13:00—14:00 时出现明显的高峰。游走行为比例在秋季及春季迁徙期波动极明显,秋季迁徙出现在8:00—9:00、13:00—14:00 时2个高峰,春季迁徙的2个高峰往后推迟1h。理羽行为比例在秋季迁徙期的8:00—9:00 时出现较小高峰,越冬晚期在13:00—14:00 时出现特别明显的高峰,春季迁徙期在9:00—10:00 时出现明显高峰。静栖行为比例秋季迁徙期出现早高峰,春季迁徙期较其他时间段多,且在14:00—15:00 时出现明显的高峰。

图1 白鹤幼鹤各行为日节律Fig.1 Daily rhythm of juvenile Siberian Crane
3.3 白鹤幼鹤从雄、雌成鹤获得食物的差异
采用独立样本T检验分析幼鹤从雄、雌成鹤处获得食物频次差异及单因素方差分析不同时间段从雄、雌成鹤处获得食物频次差异(表2),整个观察期间,幼鹤从雄鹤处获得食物的频次与从雌鹤处获得食物频次无显著差异,但越冬早期幼鹤从雄鹤处获得食物频次显著多于从雌鹤处获得食物频次,越冬晚期则相反。随着时间的变化,幼鹤从雄、雌成鹤处获得食物的频次都出现显著变化。
比较幼鹤从雄、雌成鹤处获得食物频次随时间变化(图2),在秋季迁徙期及越冬早期,从雄鹤处获得食物频次比雌鹤多;在越冬中期及晚期,从雄鹤处获得食物频次比雌鹤少;春季迁徙期,幼鹤从雄、雌成鹤处获得食物频次都减少至接近0 次/h。
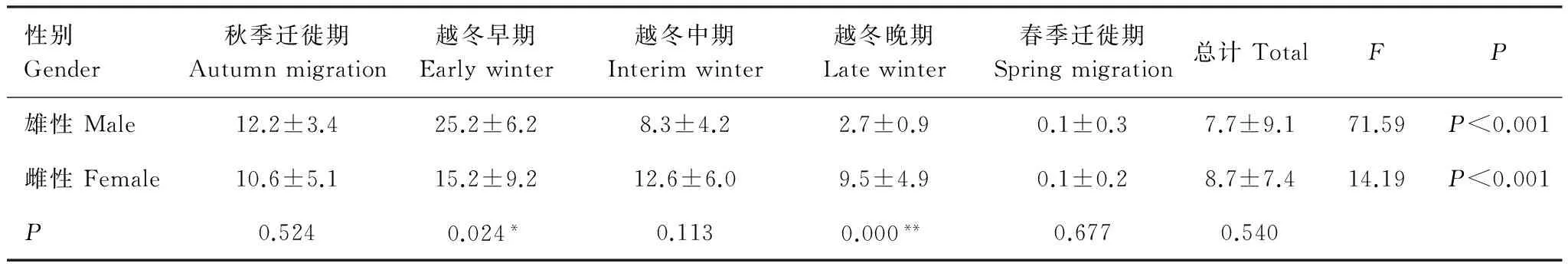
表2 白鹤幼鹤从成鹤处获得食物频次比较(次/h)Table 2 The frequency of juvenile crane obtained food from adult Siberian Cranes

图2 白鹤幼鹤从雄、雌成鹤处获得食物频次比较 Fig.2 The frequency of juvenile crane obtained food from adult Siberian Cranes
3.4 风速和气温对白鹤幼鹤行为的影响
通过单因素方差分析(表3)及LSD比较,在不同风速下,幼鹤仅有静栖行为有显著变化,即当风速大于4级时其静栖行为比例显著增加(P<0.001)。
统计不同时间段日最高气温与最低气温(表4),并采用单因素方差分析不同时间段之间气温的差异,表明不同时间段的最高气温和最低气温都存在极显著差异。LSD多重比较分析显示,秋季迁徙期的最低气温极显著低于其他时间段的最低气温(P<0.001),而越冬期的3个时间段及春季迁徙期的最低温之间无显著差异。秋季迁徙期的最高气温极显著低于越冬晚期及春季迁徙期(P<0.001),显著低于越冬早期(P=0.036);越冬中期的最高气温显著低于越冬晚期(P=0.015);春季迁徙期的最高气温极显著的高于秋季迁徙期和越冬早、中期(P<0.001),且显著高于越冬晚期(P=0.013)。采用Pearson相关分析最高气温及最低气温对幼鹤行为的影响(表5)表明,幼鹤取食行为和警戒行为与气温呈极显著的正相关,乞食行为与气温呈极显著的负相关,游走行为与当日最低气温呈显著的负相关;其他行为与气温无显著相关性。

表3 不同风力下白鹤幼鹤各行为分配比例的单因素方差分析Table 3 ANOVA of wind velocity on juvenile Siberian Crane activity time(**P<0.01, *P<0.05)

表4 各时间段平均气温及差异Table 4 ANOVA of temperature at different periods
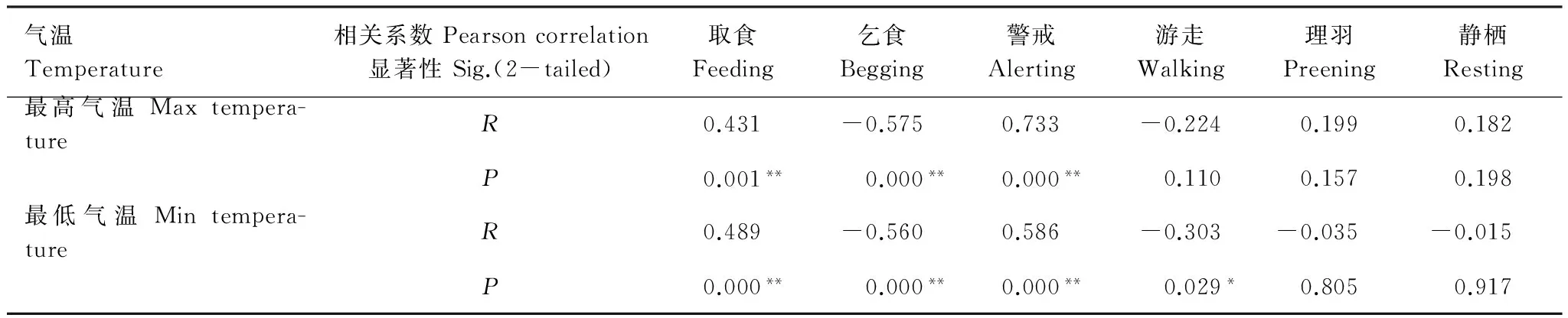
表5 气温对白鹤幼鹤行为影响的相关性分析Table 5 Correlation analysis on behaviors of juvenile Siberian Crane with changes in temperature(**P<0.01, *P<0.05)
4 讨论
随着白鹤幼鹤的生长,其取食行为比例逐渐增加,乞食行为比例逐渐减少;到越冬晚期,即2013年1月,幼鹤取食行为比例达到81.3%,基本与成鹤的取食行为比例达到一致[25],此时幼鹤乞食行为比例减少至2.4%。说明幼鹤取食行为在其出生次年的1月份就基本稳定,具备独自获取食物的能力。但幼鹤取食+乞食的时间比例为83.7%,高于成鹤,表明幼鹤获取食物的效率低于成鹤,因此需要花费更多的时间觅食。此结果与Jia等[21]于2011年3月在鄱阳湖区的野外观察结果一致;云南纳帕海越冬黑颈鹤幼鹤取食效率也同样低于成鹤[26]。
食物资源是影响鸟类取食时间的重要因素[27]。2011年3月,贾亦飞等在鄱阳湖保护区采用焦点取样法观察浅水区域活动的白鹤幼鹤,其取食行为比例为88%[21],明显高于本次研究结果。推测原因可能与鄱阳湖区2011年与2013年的白鹤食物丰富度差异有关。其次本研究观察是以家庭为单位的活动群,这类明显的家庭群一般与白鹤大群的间距在20 m以上。有研究表明,与集群活动的黑颈鹤对比,以家庭为单位活动的黑颈鹤的优势主要体现在有利于幼鹤的成长[26],具有领域的繁殖家庭其幼鹤的成活率要明显高于集群中的幼鹤[28]。
白鹤幼鹤的警戒行为随时间推移而逐步增加。越冬后期及春季迁徙期比越冬中期之前的时间段其警戒行为增加显著,但幼鹤的警戒行为比例仍然明显低于成年白鹤[24]。在鄱阳湖保护区,在草洲生境觅食的白鹤幼鹤其警戒行为比例低于在浅水区觅食的幼鹤[21];采用瞬时扫描法观察单个个体白鹤行为时,幼鹤的警戒行为比例也是明显低于成鹤[29]。
最适觅食理论预测动物应选择最好的觅食斑块[30]。白鹤在刚到达一个新的栖息地时,会搜寻最好的觅食生境,因此其游走行为比例会比较高,如秋季迁徙期、越冬早期及春季迁徙期的游走行为比例都显著高于越冬中期和晚期。越冬晚期游走行为比例稍高于越冬中期,与鄱阳湖区渔民的捕鱼活动有关,1月份是鄱阳湖区渔民捕鱼的最佳时节,渔民的捕鱼活动会引起白鹤游走和警戒行为的增加。
2013年1月之后,幼鹤行为模式基本稳定,取食行为比例接近于成鹤,具备独自获取食物的能力,并能够满足身体的营养需求。在春季迁徙期,幼鹤取食行为比例显著下降且理羽和静栖行为比例显著增加,推测与春季迁徙期莫莫格保护区日最高气温显著增加有关。当白昼最高气温高于某一值时,幼鹤用于维持身体热量散失的能量减少,幼鹤只需花费较少的时间取食就可以获得足够的能量,在春季迁徙期幼鹤取食行为主要受最高气温的影响。
白鹤幼鹤在秋季及春季迁徙期的理羽行为及静栖行为比例显著高于越冬期,且在春季迁徙期显著高于其他时间段。东北地区春季和秋季为多风时节,而且风力较大,白鹤在觅食过程中常常需要整理被风吹乱的羽毛,从而导致理羽行为比例增加。幼鹤静栖行为比例在迁徙停歇地较高,可能与食物种类及丰富度及季风有关。有研究显示,莫莫格保护区白鹤主要食物藨草球茎的密度为(46.0±44.3)个/m2(2010年)[31],鲜重为(1.60±0.86)g,干重为(0.83±0.61)g;而鄱阳湖保护区白鹤主要食物苦草冬芽的密度为(38.6±38.8)个/m2(2012年)、(18.2±13.6)个/m2(2011年)、19.0 个/m2(1988年),鲜重为(1.03±0.30)g,干重为(0.30±0.10)g[32- 33]。莫莫格保护区的藨草球茎密度及单个球茎的营养物资含量都要稍高于鄱阳湖保护区的苦草冬芽,幼鹤可以花费较少的时间获得足够的食物。因此,在莫莫格保护区幼鹤取食+乞食行为比例减少,可以用更多的时间进行理羽和静栖。灰鹤、东方白鹳等都有类似的为应对风速增大而增加理羽及静栖时间的行为[34- 35]。本研究显示,当风速大于4级时,幼鹤的静栖行为比例显著增加,幼鹤可能为减少风引起的羽毛凌乱及热量散失而增加静栖行为。
幼鹤取食行为日节律在秋季迁徙期、越冬晚期及春季迁徙期有明显的低谷,且春季迁徙期的取食低谷比秋季迁徙及越冬晚期的时间推后1h,而在取食低谷出现了幼鹤的游走高峰,可能与气温升高及光照时间延长有关。幼鹤的理羽行为只在越冬晚期的13:00—14:00 时出现一个明显的高峰,而这时是幼鹤取食行为的低谷。
从秋季到春季整个抚育期来看,幼鹤从雌、雄成鹤处获得食物的频次无显著差异,但不同时间段幼鹤从雄、雌成鹤处获得食物的频次差异显著,在越冬早期之前幼鹤从雄鹤处获得食物频次较高,之后是从雌鹤处获得更多食物。随着幼鹤的生长,幼鹤从成鹤处获得食物的频次显著减少,幼鹤乞食占获得食物总时间比例,即乞食/(取食+乞食)从秋季迁徙期的71.7%逐渐减少为春季迁徙期的1.9%,此结果与Sauey[14]在印度对白鹤野外观察的结果基本一致。
最适觅食理论说明动物从食物中获得的能量必须多于为获取食物所消耗的能量[36]。鸟类会权衡其能量获得与散失来调整其行为[37],在低温环境下,水鸟可能会放弃包括取食在内的行为活动,从而降低自身的能量消耗[38]。白鹤幼鹤的取食行为与气温呈显著正相关,当气温降低时,幼鹤会减少其主要活动来减少热量的散失,同时幼鹤也会通过减少取食行为但增加乞食行为,以此来获得足够的能量和营养。
白鹤幼鹤的各行为变化充分反映出了它的生长轨迹。幼鹤在秋季迁徙期至越冬早期,其生理及行为都不成熟,获得食物的能力有限,需要依赖其父母才能获得足够食物;此时失去父母的孤幼鹤将无法顺利完成秋季迁徙及越冬[16,19]。到越冬后期及春季迁徙时,幼鹤已具备独自获得足够食物的能力,且此时幼鹤各行为日节律基本与成鹤一致。本次研究结果显示,幼鹤生长到能独立生存的时间节点为其出生次年的1月。Sauey[14]也得出大致同样的结论,在印度克拉迪奥国家公园越冬的白鹤幼鹤,于次年的1月其自己取食所获得的食物量超过从其父母处获得食物量,具备了能离开其父母独立生存的能力。
[1] ICF. Siberan Crane. [2014-4-8]. http://www.savingcranes.org/siberian-crane.html.
[2] 郑光美. 中国濒危动物红皮书:鸟类. 北京:科学出版社, 1998:22- 214.
[3] IUCN. IUCN Red List of Threatened Species. (2013.1) [2013- 6- 26]. http://www.iucnredlist.org.
[4] Meine C D, Archibald G W. The Cranes Status Survey and Conservation Action Plan. Gland, Switzerland: IUCN. 1996: 1- 294.
[5] 单继红, 马建章, 李言阔, 钱法文, 涂晓斌. 近十年来鄱阳湖区越冬白鹤种群数量与分布. 动物学研究, 2012, 33(4): 355- 361.
[6] 许杰, 蒋星星, 宋胜利. 濒危动物白鹤. 野生动物, 1985, 6(3): 30- 31.
[7] 吴志刚, 韩晓东. 莫莫格自然保护区的珍稀水禽.野生动物, 1992, 13(3): 20- 23.
[8] 何春光, 宋榆钧, 郞惠卿, 李鸿凯, 孙效维. 白鹤迁徙动态及其停歇地环境条件研究. 生物多样性, 2002, 10(3): 286- 290.
[9] Kanai Y, Ueta M, Germogenov N, Nagendranc M, Mitad N, Higuchie H. Migration routes and important resting areas of Siberian cranes (Grusleucogeranus) between northeastern Siberian and Chian as revealed by satellite tracking. Biological Conservation, 2002, 106(3): 339- 346.
[10] 崔茂欢, 郑礼琼. 白鹤秋季迁徙期行为观察. 林业调查规划, 2006, 31(4): 94- 97.
[11] 刘小强, 朱成伟, 赵玉海, 姜亮. 沈阳獾子洞湿地白鹤及其停歇地生境调查. 野生动物杂志, 2008, 29(4): 184- 186.
[12] 邹畅林,王波,王永. 莫莫格春季白鹤迁徙报告. 鹤类通讯, 2008, 12(1): 24.
[13] 江红星. 吉林莫莫格国家级自然保护区2010年春季白鹤迁徙日最高统计达3128只. 中国鸟类研究简讯, 2010, 19(1): 6, 28.
[14] Sauey R T. The feeding biology of juvenile Siberian Cranes in India. Proceedings, 1978 Crane Workshop. Ed. James C. Lewis. [Fort Morgan, Colo.]: Colorado State Univ and National Audubon Soc, 1979: 205- 210.
[15] Spitzer P R. The Siberian Crane at Bharatpur. Proceedings of 1978 Crane Workshop. Ed. James C. Lewis. [Fort Morgan, Colo.]: Colorado State Univ. and National Audubon Soc, 1979, 249- 253.
[16] Archibald G W, Mirande C. Status and conservation of the Siberian CraneGrusLeucogeranus. Vogelwelt, 1999, 120 (5/6): 377- 381.
[17] 胡振鹏. 白鹤在鄱阳湖越冬生境特性及其对湖水位变化的响应. 江西科学, 2012, 30(1): 30- 36.
[18] Archibald G W, Shigata Y, Matsumoto K, Momose K. Endangered cranes. Crane Research around the World. Proceedings of the International Crane Sympfosium at Sapporo, Japan, in 1980 and Papers from the World Working Group on Cranes, International Council for Bird Preservation. Eds. J.C. Lewis and H. Masatomi. International Crane Symposium at Sapporo, Japan, 1981: 1- 12.
[19] Sauey R T. The range, status, and winter ecology of the Siberian Crane (GrusLeucogeranus). PhD. Thesis. Cornell University, 1985: 212- 284.
[20] 李枫, 汪青雄, 卢珊, 蔡勇军. 扎龙湿地白鹤春季停歇地昼间行为时间分配及活动规律. 动物学杂志, 2007, 42(3): 68- 72.
[21] Jia Y, Jiao S, Zhang Y, Zhou Y, Lei G, Liu G. Diet Shift and Its Impact on Foraging Behavior of Siberian Crane (GrusLeucogeranus) in Poyang Lake. PLoS ONE, 2013, 8(6): e65843.
[22] 苏化龙, 林英华, 李迪强, 钱法文. 中国鹤类现状及其保护对策.生物多样性, 2000, 8(2): 180- 191.
[23] 周福璋,丁文宁. 白鹤(Grusleucogeranus)越冬习性. 动物学杂志, 1982, 17(4): 19- 21.
[24] 严丽,丁铁明. 江西鄱阳湖区白鹤越冬调查. 动物学杂志, 1988, 23(4): 34- 36.
[25] 张琼, 钱法文. 鄱阳湖白鹤家庭行为研究. 动物学杂志, 2013, 48(5): 759- 768.
[26] 王 凯, 杨晓君, 赵健林, 余红忠, 闵龙. 云南纳帕海越冬黑颈鹤日间行为模式与年龄和集群的关系. 动物学研究, 2009, 30(1): 74- 82.
[27] Enoksson B. Time budgets of NuthatchesSittaeuropaeawith supplementary food. Ibis, 1990, 132: 575- 583.
[28] Alonso J C, Bautista L M, Alonso J A. Family-based territoriality vs flocking in wintering common cranes (Grusgrus). Journal of Avian Biology, 2004, 35: 434- 444.
[29] 王宇嘉, 范巍巍, 李晓民. 吉林莫莫格白鹤行为研究. 野生动物, 2012, 33(2): 67- 70.
[30] 尚玉昌. 动物行为学. 北京:北京大学出版社, 2005: 152- 181.
[31] 刘春悦, 江红星, 孙效维, 邹畅林, 王波, 钱法文, 吕宪国. 白鹤中途停歇地主要食物藨草球茎密度的空间差值方法研究. 动物学杂志, 2013, 48(3): 382- 390.
[32] 贾亦飞. 水位波动对鄱阳湖越冬白鹤及其他水鸟的影响研究[D]. 北京:北京林业大学, 2013.
[33] 熊秉红, 李伟. 鄱阳湖自然保护区蚌湖和中湖池苦草冬芽的调查. 水生生物学报, 2002, 26(1):19- 24.
[34] Yang C, Hou Y, Zhou L. Behaviors of the Oriental White Stork(Ciconiaboyciana) in a semi-natural enclosure. Chinese Birds, 2013, 4(2): 161- 169
[35] 张希明, 麻友俊,何百锁, 李晓明. 灰鹤秋季迁徙行为研究. 四川动物, 2010, 29(1): 105- 108.
[36] MacArthur R H, Pianka E R. On optimal use of a patchy environment. Am. Not, 1966, 100:603- 609.
[37] Cherel Y, Robin J, Maho Y. Physiology biochemistry of long-term fasting in birds. Can J Zool, 1988, 66:159- 166.
[38] Paulus S L. Time-activity budgets of non-breeding Anatidae. University of Minnesota. Waterfowl in winter, 1988, 135- 152.
Behavioral development of the juvenile siberian crane
ZHANG Qiong, QIAN Fawen*
ResearchInstituteofEcology,EnvironmentandProtection,ChineseAcademyofForestry,Beijing100091,China
From October 2012 to May 2013, juvenile Siberian Cranes (Grusleucogeranus) were studied by the focal sampling method at Momoge National Nature Reserve in Jilin Province and Poyang Lake National Nature Reserve in Jiangxi Province, China. The results showed that the juvenile Siberian Cranes spent 23.0% of their time budget on feeding in the fall migration, increasing to 82.7% in late winter, but reduced to 61.9% in the following spring at Momoge National Nature Reserve. In comparison, the begging behavior reduced from 58.2% in the fall migration to 1.2% in the following spring migration, and the alert behavior increased from 1.0% to 7.1%. The behaviors of preening and resting in the following spring were significantly more than those in the wintering period, the reason for the difference may come from the different food availability. For environmental factors, wind velocity had a significant impact on resting behavior, while temperature was positively correlated with feeding and alert behaviors. The daily activity rhythm of juvenile Siberian Cranes presented a trough in feeding behavior at 13:00—14:00 in the fall migration and late wintering period; however, this trend was delayed by one hour in the next spring migration period. Throughout the whole observation period, there was no difference in the frequencies of juvenile Siberian Cranes obtaining food from their parents, but differences did occur between the parental contributions in the different periods. This study aimed to elucidate the growth process of juvenile Siberian Cranes and parental contributions to the growth of their offspring through ethological observations.
juvenile siberian crane; behavior; growth; feeding; habitat
国家林业局保护司珍稀濒危物种调查监管项目(1691300000020)
2014- 02- 18;
日期:2015- 04- 14
10.5846/stxb201402180278
*通讯作者Corresponding author.E-mail: cranenw@caf.ac.cn
张琼,钱法文.白鹤幼鹤行为发育.生态学报,2015,35(21):7236- 7243.
Zhang Q, Qian F W.Behavioral development of the juvenile siberian crane.Acta Ecologica Sinica,2015,35(21):7236- 7243.
猜你喜欢
心声歌刊(2023年2期)2023-07-16 05:56:02
哈哈画报(2022年1期)2022-04-19 11:27:20
今日农业(2021年21期)2021-11-26 05:07:00
黄河之声(2021年7期)2021-06-21 13:59:24
今日农业(2020年13期)2020-08-24 07:35:24
现代园艺(2017年11期)2017-06-28 11:32:51
意林(2017年8期)2017-05-02 17:40:37
中国农业文摘-农业工程(2016年5期)2016-04-12 05:38:13
现代农业(2016年4期)2016-02-28 18:42:06
医学研究杂志(2015年5期)2015-06-10 06:43:26
