
家庭养育环境与儿童发展:差别易感性假说
2015-02-05 03:40
心理科学进展 2015年11期
(首都师范大学心理学系,北京 100048)
家庭养育环境在儿童的发展过程中发挥着至关重要的作用。差别易感性假说认为,家庭养育环境对儿童的影响无论是有利的还是不利的,其程度都是不同的。本文首先介绍了差别易感性模型的基本观点和统计标准,以及关于差别易感性模型的一些实证研究,最后总结了该领域研究的局限性,并对未来的研究方向进行了展望。
1 差别易感性模型的基本观点和检验步骤
1.1 差别易感性模型的基本观点
Rosenthal(1963)首次使用素质压力理论(diathesis-stress theories)来解释精神分裂症的病因,认为精神分裂症是个体遗传素质与环境中的压力因素相互作用的结果。这里的素质是脆弱性的同义词,包括基因、生物、生理、认知和人格等相关因素(Sigelman&Rider,2009)。素质压力理论认为,个体对特定心理障碍的素质倾向可以由生活中的压力事件激发,如果个体是弹性的,那么只有较高的生活压力才能使个体患病;相反,如果个体的脆弱性很高,那么即使较低的压力也会使个体患病(Goforth,Pham,&Carlson,2011)。该理论可以解释为什么在经历相同的压力环境时并不是所有人都会出现心理障碍。
之后发展心理学家也用精神病理学领域中的素质压力模型来解释那些处于消极环境中的脆弱性个体的发展问题,Belsky和Pluess(2009a)将之称为双重风险模型(dual-risk model)。素质压力或双重风险模型的重点是,一些个体由于自身具有的“脆弱性”而对环境中的压力事件更易感,即图1中黑色实线所代表的个体,这些“脆弱性”可能是个性特征中的行为或气质因素(例如,困难气质)、本质上的生理或内表型因素(例如,高生理反应)以及起源上的基因因素(例如5-HTTLPR短等位基因)(Belsky&Pluess,2009a),还有一些弹性个体由于缺乏这些脆弱性因素而不易受到不利环境的影响(Cicchetti&Garmezy,1993)。这一理论观点得到很多领域中实证研究的支持(Belsky,Hsieh,&Crnic,1998;Caspi et al.,2003;Cummings,El-Sheikh,Kouros,&Keller,2007)。但素质压力或双重风险模型主要关注的是消极环境对脆弱性个体的不利影响(见图1中左边黑色实线方框部分),并未说明个体对积极环境的反应差异问题,实际上该模型暗含着弹性个体和脆弱性个体对积极环境或非逆境环境的影响没有差异(Ellis,Boyce,Belsky,Bakermans-Kranenburg,&van IJzendoorn,2011)。然而,有证据表明某些个体在消极环境和积极环境中都具有发展可塑性,而这种情形并不符合素质压力或双重风险模型的假定。
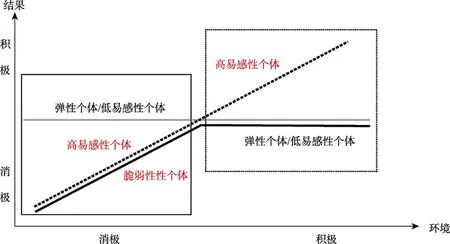
图1 素质压力/双重风险模型和差别易感性模型
1997年,Belsky首次提出了差别易感性模型(Differential Susceptibility Model)(图1中黑色虚线和灰色实线部分),认为高易感性的个体(图1中黑色虚线所代表的个体)虽然具有某种“脆弱性”,但是这种“脆弱性”也可被称作是“发展可塑性”,它使个体在易受消极环境的不利影响的同时,也会不成比例地易受到积极环境的积极影响,即在素质压力或双重风险模型的基础上加上了右边黑色虚线方框部分,这也是差别易感性模型区别于素质压力或双重风险模型的部分。换言之,环境是以一种更好或更差的方式在影响着个体的发展(Belsky,1997a,2005;Beklsy&Pluess,2009a,2009b,2013a)。这与上述众所周知的素质压力或双重风险模型不同,差别易感性模型强调个体对环境影响的差异性并不仅仅局限于逆境的负面影响中,个体受积极经历或体验的影响也存在着差异性。自从差别易感性模型提出之后,研究者对该模型进行了多方面的探索和验证,尤其是对儿童在家庭养育环境中的差别易感性进行了大量研究。实证研究的证据表明,确实有些儿童无论是暴露在消极养育环境中还是积极养育环境中都具有发展可塑性(Bradley&Corwyn,2008;Pluess&Belsky,2009)。那么,究竟如何通过实证研究证实差别易感性模型?其检验步骤和诊断标准是什么?
1.2 差别易感性模型的检验步骤和诊断标准
我们不能假定所有的易感性因素(如气质、基因等)×环境的交互作用研究都符合差别易感性理论模型,为了将差别易感性从其他交互作用中区分出来,Belsky,Bakermans-Kranenburg和van IJzendoorn(2007)提出了检验差别易感性模型的5个步骤和统计标准。根据差别易感性模型的假设,环境对个体发展的影响受到个体易感性因素的调节。因此,正如模型所述,个体的各种易感性因素(包括行为的、生理的和基因的等因素)为调节变量,环境因素为预测变量,个体的行为发展结果为因变量(Belsky et al.,2007)。2005年,Belsky只是根据数据模型的视觉显示这样一个简单的评估方法来检验差别易感性模型,而且为了提高差别易感性的预期,Belsky使用了一个比较自由宽松的标准。后来,为了完善差别易感性的评估方式,Belsky等人(2007)设定了更具体、更严格的统计标准,除了检测环境预测变量和调节变量之间的显著交互作用,也需要保证高易感性群体的斜率显著不等于0,同时其斜率的绝对值显著大于低(非)易感性群体斜率的绝对值。
Belsky等人(2007)提出了差别易感性模型的具体检验步骤。第一,检验易感性因素与环境因素之间是否存在交互作用(Belsky et al.,2007);第二,检验易感性因素和环境因素之间没有相关,这是检验差别易感性的关键步骤。例如,如果消极情绪性和父母的教养方式之间相关,则不能证明父母教养方式对于高消极情绪性婴儿具有更大的预测力度(Belsky et al.,2007);第三,检验易感性因素和行为结果之间的相关关系。如果其相关不等于0,那么差别易感性模型不成立。这一步的目的是为了把差别易感性模型从素质压力或双重风险模型中分离出来;第四,将回归图与差别易感性经典图形进行比较。只有符合图1中黑色虚线和灰色实线部分才证明是差别易感性模型,并且要求高易感性群体的斜率显著不等于0,同时其绝对值显著大于低(非)易感性群体斜率的绝对值;若符合图1黑色实线和灰色实线部分,则证明是素质压力或双重风险模型(Ellis et al.,2011);最后,替换易感性因素和行为结果,检验模型的特异性。
但是,有研究者认为以往所使用的检验差别易感性的方法存在着严重的局限性,例如Roisman等人(2012)对差别易感性的解释提出了视觉检查、量化、Ⅰ型错误和非线性等4点批评。Belsky等人(2015)对前人的批评建议进行了总结,概括出几种诊断差别易感性的更严格的标准:
第一,使用显著性域(RoS)检验来决定易感性因素M(如,困难气质)的值域,并且结果变量Y对预测变量X的回归在易感性因素的值域上是统计显著的,预测变量的范围应界定为X±2SD(Roisman et al.,2012);第二,使用交互效应比例(PoI)和受影响比例(PA)两个新标准。PoI指在预测变量值域为X±2SD范围内的交互效应线形图中位于交叉点之上的面积占总面积的比例。在差别易感性模型图中,交叉点位于预测变量平均值附近,因此PoI约为50%,代表“更好”的区域;PA指数与PoI相似,但PA衡量的是位于交叉点之上即受到“更好”影响的人数的比例,在差别易感性条件下,50%的人将由于预测变量(例如,教养)的影响而受易感性因素的“更好”影响。在使用这种方法时,差别易感性理论要求PoI的范围在40%-60%或PA≥16%(Roisman et al.,2012);第三,涉及非线性验证。在得出差别易感性结论之前要证明X2、MX2在统计上不显著,且X2和MX2之和也不显著;如果一项或多项是显著的,那么须证明对这些非线性项进行了统计控制之后X和M的交互作用仍然存在(Roisman et al.,2012);第四,当对多波数据集检查多元交互效应时应当考虑Ⅰ型错误。可用多层次模型来检验跨发展过程的差别易感性,考虑多重比较时可适当调整α系数(Roisman et al.,2012)。此外,还有重新参数化回归模型拟合(Widaman et al.,2012)和将数据与先验模型进行拟合(Belsky,Pluess,&Widaman,2013)等诊断差别易感性模型的方法。
2 差别易感性模型的证据
根据近年来有关养育环境或养育经历下的差别易感性的实证研究,我们选择了一些能够较好地证明儿童对养育环境的影响具有差别易感性的证据,这些证据均表明儿童自身具有不同的可塑性,一些儿童会以一种更好或更差的方式受到家庭养育环境的影响。以往该领域的研究将儿童的易感性因素主要分为行为、生理和基因因素三种,下面我们将对上述这三种易感性因素的有关实证研究进行回顾和梳理。
2.1 对养育环境差别易感性的行为标志
令发展心理学家最感兴趣的话题之一,就是儿童的个性特征和养育经历如何共同塑造着儿童的发展(Rothbart&Bates,2006),差别易感性模型就是要回答上述问题。其中,在差别易感性行为因素的研究中,研究者关注最多的就是儿童自身的困难气质。有关儿童气质与养育环境的交互作用的研究为差别易感性模型提供了强有力的证据,这些研究既包括儿童的消极情绪性或困难气质与父母教养的交互作用,也涉及儿童气质对儿童保育经历的调节作用,研究表明儿童的气质和情绪特征改变着后天养育环境或经历对儿童发展的影响(Belsky,2005;Belsky et al.,2007)。
2.1.1 儿童气质与父母教养环境
基于差别易感性模型的基本观点,不同气质类型的儿童对亲代教养环境的影响的敏感性是不同的。对于困难气质的儿童来说,他们更容易受到不良教养经历的不利影响,同时他们也可以从积极的、支持性的教养经历中获得更多的益处(Belsky,1997a,1997b)。一般地,困难气质的儿童具有倾向于消极情绪表达、低适应性和高活动性等特性。有研究发现,与那些在12个月大时没有表现出高水平痛苦的儿童相比,父母消极的教养方式更能预测那些表现出高痛苦水平的儿童在36个月时的外化问题和抑制行为,这一研究结果很好地支持了差别易感性模型(Belsky et al.,1998)。关于提高母亲对不安全型依恋婴儿风险的敏感性的干预研究发现,只有对高消极反应性的婴儿来说,父母教养敏感性的提高有助于增进他们的依恋安全性(Velderman,Bakermans-Kranenburg,Juffer,&van IJzendoorn,2006)。新近的研究也发现,困难气质的儿童如果早期经历了严厉的父母教养则会表现出更多外化问题,如果经历的是积极的父母教养方式,则会得到相反的研究结果(Poehlmann et al.,2012)。
上述研究对婴儿困难气质的测量均采用的是实验室观察法,提供了差别易感性模型的证据,此外基于母亲对婴儿气质报告的研究也提供了相关证据。例如,关于儿童9个月时母婴互动同步性和儿童2岁时自我控制的研究表明,对婴儿期就具有困难气质的儿童来说,相互同步的母婴互动更好地预测了儿童的自我控制(Feldman,Greenbaum,&Yirmiya,1999)。还有研究发现,对痛苦敏感性得分较高的的儿童来说,严厉和敌对的母亲教养是儿童1~2年级时的外化行为的一个很好的预测因素。对困难气质的儿童来说,父母的教养除了能够很好地预测其外化行为之外,还在一定程度上能够预测儿童的社会情感问题(Morris et al.,2002)。
美国儿童健康和人类发展研究所(NICHD)使用婴儿气质问卷测量了6个月大儿童的气质。结果表明,对于困难气质儿童,母亲敏感性和儿童行为问题之间的关系更为强烈(Bradley&Corwyn,2008)。之后,研究进一步发现,教养质量能反向预测儿童一年级时教师报告的问题行为,但这种预测作用要受到儿童早期气质的调节,对于困难气质儿童,较高质量的教养能预测更少的问题行为,较低质量的教养会预测更多的问题行为,但是对中等和低困难气质儿童的预测作用较弱(Bradley&Corwyn,2008)。在我国,也有证明差别易感性模型的实证研究。对学前儿童的父亲情绪表达、儿童气质与儿童社会适应的关系的调查表明,对消极情绪性较高的儿童来说,父亲消极情绪表达能够显著正向预测其内隐行为问题,父亲积极情绪表达可以显著正向预测其社会能力;而对消极情绪性较低的儿童则没有显著的预测作用(梁宗保,孙铃,张光珍,陈会昌,张萍,2011))。因此,上述研究结果均表明,儿童的气质会以一种更好或更差的方式调节家庭教养环境和儿童发展之间的关系。
2.1.2 儿童气质与儿童保育
上述研究重复表明,消极情绪性或困难气质的儿童会以更好或更差的方式受到家庭教养环境的影响,但差别易感性的证据并不局限于对家庭教养环境的研究中,儿童早期的保育经历也是儿童气质的函数(Belsky,2005)。早在1995年,对儿童社会恐惧和保育质量的研究发现,母亲报告的儿童社会恐惧和儿童保育质量之间有明显的交互作用,儿童保育质量可以预测一年以后儿童的非社会性活动(如独自玩耍、旁观者行为)。由于儿童保育质量的高低不同,高恐惧儿童会有最低和最高的非社会性活动,然而这种影响在低恐惧儿童中并未出现(Volling&Feagans,1995)。还有研究发现,相比于其他类型的看护,当儿童受到儿童保育看护时,易受挫的婴儿在30个月大时会表现出更多外化行为,但是对那些在面对挫败时有较少苦恼的儿童来说并没有这样的影响(Crockenberg&Leerkes,2005)。一项将儿童的社交恐惧作为调节因素的研究发现,没有压力的儿童保育环境对困难气质儿童来说尤为重要。对高恐惧的儿童来说,当看护者的皮质醇水平较低时,高恐惧的儿童会报告更高的幸福感,而当看护者的皮质醇水平上升时,儿童报告的幸福感会降低(Groeneveld,Vermeer,van IJzendoorn,&Linting,2012)。这些证据也都表明,困难气质的儿童以更好或更差的方式受到儿童保育质量的影响。
上述研究均为横断研究的结果,来自NICHD大型追踪数据库的研究也明确验证了差别易感性假说。例如研究发现,前4.5年的儿童保育质量对儿童行为问题和社会能力的影响会受到儿童气质的调节,相比于容易气质的儿童,那些在婴儿时期就表现出困难气质的儿童在低质量的保育环境中会有更多的行为问题,在高质量的保育环境中会有更少的行为问题,这一结果很好地验证了差别易感性假说(Pluess&Belsky,2009)。当研究者把儿童的年龄范围扩展到童年中期末时,也发现了差别易感性的证据,即儿童4.5岁之前的保育质量能够有效地预测10~11岁有困难气质的儿童的认知-学业成绩和社会功能(Pluess&Belsky,2010)。这些使用NICHD追踪数据库的研究,不但有力地支持了差别易感性模型,同时将差别易感性的研究从童年早期延伸到了童年中期末。
尽管关于儿童气质与教养环境交互作用的研究已经很多,但是儿童自身的某些气质特性可能本来就与父母的教养存在着一定的相关,这样差别易感性个体的可塑性差异的解释就值得重新考虑,因此,在测量气质和教养的交互作用之前,研究者有必要控制前者对后者的唤起效应(Belsky&Pluess,2009a)。还有,对于儿童保育来说,只有当保育质量作为预测因素时出现了差别易感性的证据,而对儿童接受的保育量来说并没有出现差别易感性的情况(Pluess&Belsky,2010)。
2.2 对养育环境差别易感性的生理标志
环境生物敏感性理论(biological sensitivity to context theory)认为,对压力表现出高生理反应性的儿童具有最大的发展可塑性(Boyce&Ellis,2005),该理论观点强调了个体差别易感性的生理特征,这些生理因素可能会调节环境对个体产生的影响,是可塑性的标志。这些差别易感性的生理标志主要来自于两个独立的生理系统——自主神经系统和内分泌系统。
2.2.1 自主神经系统引起的生理反应
与差别易感性假说相一致,交感神经系统(SNS)引起的某些生理反应能够调节环境和个体外化问题之间的关系。Boyce等人(1995)对儿童呼吸系统疾病的研究发现,有较高血压反应的儿童在高压力环境中要比其他儿童有更高的发病率,但是在低压力环境中发病率则会显著降低。El-Sheikh,Keller和Erath(2007)在一个问题解决任务中研究了SCL(受交感神经系统单独控制)反应性,发现和较低SCL反应性女孩相比,高冲突家庭中的高反应性女孩的外化问题会有最大程度的增加,而低冲突家庭中的高反应性女孩则会有最小的增加。关于社会支持和迷走神经监管能力的研究也支持了差别易感性假说。迷走神经监管力是面对各种压力事件时迷走神经灵活反应的一个指标。Wolff,Wadsworth,Wilhelm和Mauss(2012)对4~5岁儿童的研究发现,相对于迷走神经监管能力低的儿童,高迷走神经监管力儿童在支持性的社会环境中有较低的SNS压力反应,而在非支持环境中的SNS压力反应则会较高。这些交感神经系统引起的生理反应似乎也是以一种更好或更差的方式影响着儿童的发展。
同时,副交感神经系统(PNS)引起的生理反应也作为调节因素发挥着作用。有研究者使用迷走神经张力和迷走神经抑制来调查副交感神经系统是否以及如何调节家庭中婚姻冲突与童年中期儿童的适应能力的关系。他们发现相比于高迷走神经张力的儿童,那些生长在较少婚姻冲突的家庭中的低迷走神经张力的儿童会出现更低的焦虑,而当处于高婚姻冲突的家庭中时则会表现出更高的焦虑;对迷走神经抑制的研究也出现了相似的证据(El-Sheikh,Harger,&Whitson,2001)。Obradović,Bush,Stamperdahl,Adler 和 Boyce(2010)以5岁儿童为研究对象,发现了呼吸窦性心律不齐(RSA,迷走神经张力的一个指标)对养育环境影响的调节作用。相比于呼吸窦性心律不齐反应性较低的儿童,那些有高呼吸窦性心律不齐反应的儿童,在高逆境环境中长大时会有更少的亲社会行为和学校活动参与,而当处于低逆境环境中时则相反。一项对生活在贫困环境中的婴儿的研究发现,对呼吸窦性心律不齐基线水平高的婴儿来说,当他们处于安全的成长环境中时会出现最低水平的问题行为,而当成长环境混乱时会有最高水平的问题行为(Conradt,Measelle,&Ablow,2013)。这些研究结果都与差别易感性理论相一致。还有一些研究支持或部分支持了差别易感性假说(Elisabeth de Neuf,2012;Eisenberg et al.,2012)。上述研究均表明副交感神经系统对环境的影响具有调节作用。
2.2.2 内分泌系统引起的生理反应
关于内分泌系统的差别易感性研究中主要集中于皮质醇反应性。应对压力的神经内分泌系统主要受到下丘脑-垂体-肾上腺皮质轴(Hypothalamic-Pituitary-Adrenal cortex axis)的控制,HPA轴对个体情绪调节、行为控制等过程具有重要作用。皮质醇作为HPA轴的终产物经常作为测量压力的生物学指标(聂瑞虹,许颖,韩卓,2015)。有研究者考察了皮质醇反应性对养育环境影响的调节作用,例如,Obradović等人(2010)以 5~6岁儿童为调查对象,考察了家庭磨难对儿童亲社会行为的影响,该研究将唾液皮质醇反应性作为神经生物应激反应性的指标之一,即易感性的生理因素,结果表明,相比于低水平皮质醇反应性的儿童,皮质醇反应高的儿童在低逆境环境中有较多的亲社会行为,但是在高逆境环境中则会有较少的亲社会行为。虽然许多研究将皮质醇反应性作为差别易感性的生理指标,但也有研究者认为皮质醇反应性是环境因素的结果,而不是环境影响的调节因素(Fernald,Burke,&Gunnar,2008;Gunnar&Quevedo,2007),例如,Martin,Kim,Bruce和Fisher(2014)发现不良父母教养方式会通过减缓皮质醇日常节律而增加儿童的外化问题行为。因此,关于皮质醇反应性是否是家庭环境与儿童发展的调节变量尚需进一步研究。
2.3 对养育环境差别易感性的基因型标志
由于受传统的素质压力或双重风险模型的影响,人们忽视了当那些携带“风险等位基因”的儿童当暴露于非风险环境中时实际上可能比其他儿童发展得更好这一事实。近些年,关于基因与养育环境交互作用的研究为差别易感性模型提供了基因型证据。其中,研究最多的当属5-羟色胺转运体多态性、多巴胺D4受体基因和单胺氧化酶A基因。
2.3.1 5-羟色胺转运体多态性(5-HTTLPR)
五羟色胺转运体基因连锁多态性区域上的五羟色胺转运蛋白基因负责编码五羟色胺转运体,一般五羟色胺转运体短等位基因(s/s,s/l)与五羟色胺转运体因子的表达性相关。2003年,Caspi等人在对26岁有自杀观念和重度抑郁发作的成年人的研究中首次发现,两个短等位基因(s/s)携带者最易受到生活压力事件的不利影响,抑郁水平的得分最高,而当没有生活压力事件时这些人的抑郁水平最低,但是生活压力事件对两个长等位基因(l/l)携带者的抑郁水平的影响很小甚至没有影响,这说明5-HTTLPR能够调节生活压力事件和抑郁之间的关系。Pluess,Belsky,Way和Taylor(2010)以118名15~33岁的成年人为调查对象,研究了5-HTTLPR对生活事件和神经质之间关系的调节效应,结果发现了差别易感性的证据,即对纯合子短等位基因的携带者来说,高消极的生活事件与高神经质相关,而高积极的生活事件与低神经质表现出相关。近年来的一项元分析也发现了5-HTTLPR的这种调节作用,为差别易感性模型提供了强有力的证据支持。对54项研究的元分析发现5-HTTLPR能够调节压力和抑郁之间的关系,携带5-HTTLPR短等位基因(s/s,s/l)的个体在压力环境下会增加患抑郁症的风险(Karg,Burmeister,Shedden,&Sen,2011)。此外,国内外的研究还发现,5-HTTLPR对家庭教养和青少年积极情感之间的关系也发挥着调节作用,携带两个短等位基因(s/s)的青少年在消极的、非支持性的教养环境中表现出低水平的积极情感,而在积极的、支持性的教养环境中会表现出高水平的积极情感,从而支持了差别易感性假说(Hankin et al.,2011;张卫,甄霜菊,2013)。最近一项以高加索人为研究对象的元分析研究也支持了5-HTTLPR是差别易感性的基因标志,研究发现短等位基因(s/s,s/l)携带者比两个长等位基因(l/l)携带者会更多受到消极环境的影响,然而也可以从积极环境中受益更多(van IJzendoorn,Belsky,&Bakermans-Kranenburg,2012)。
2.3.2 多巴胺D4受体基因(DRD4)
大量研究表明,携带多巴胺D4受体基因的儿童更易受到消极环境的不利影响,在积极的、支持性的环境中他们也会受益更多。例如,一项纵向研究发现,对携带7-重复多巴胺D4受体基因的儿童来说,10个月大时的母亲敏感性能预测儿童2年后的外化问题,当母亲敏感高时会有最少的外化问题,这一研究支持了差别易感性模型(Bakermans-Kranenburg&van IJzendoorn,2006)。2007年,一项以18~21个月大的儿童为研究对象的横断研究也得到了相似的结果,与没有携带7-重复多巴胺D4受体基因的儿童相比,携带者在高质量的教养环境中长大时会有更少的感觉寻求行为,而在低质量的教养环境中则相反(Sheese,Voelker,Rothbart,&Posner,2007)。同样,在一项研究中对1~3岁有较严重外化问题儿童的母亲的教养方式进行了干预,结果发现只有携带7-重复多巴胺D4受体等位基因的儿童会从中受益(Bakermans-Kranenburg,van IJzendoorn,Pijlman,Mesman,&Juffer,2008)。对儿童依恋安全性和捐赠行为的研究也发现了多巴胺D4受体基因的调节作用,只有那些携带7-重复等位基因的安全型依恋的儿童会表现出更多的捐赠行为(Bakermans-Kranenburg&van IJzendoorn,2011)。2014年,对早期的母亲看护、多巴胺D4受体基因和儿童青少年时期的外化行为的研究表明,当经历低反应性和低刺激性的母亲看护时,携带7-重复等位基因的青少年会出现更多的注意缺陷多动障碍、破坏性行为障碍和精神错乱行为,当他们成长于积极的早期看护环境中时,则会出现较少的注意缺陷多动障碍和精神错乱行为,然而那些没有携带7-重复等位基因的青少年则对母亲看护质量的影响不敏感(Nikitopoulos et al.,2014)。一项专门从差别易感性视角探讨多巴胺D4受体基因和母亲敏感性对儿童外化行为的预测作用的研究发现,儿童14个月时的母亲敏感性与其18个月时的外化行为以一种更好或更差的方式相关联,但这一结果只局限于携带至少一个7-重复基因的儿童,从而也支持了差别易感性模型(Windhorst et al.,2015)。此外,除了在家庭教养中发现了差别易感性模型的基因型证据之外,在有关儿童保育的研究中也发现了相似的证据,如有研究发现携带7-重复等位基因的儿童比那些未携带者更易受低质量的儿童保育的不利影响,表现出更多行为问题,同时也更易从高质量的儿童保育中受益(Belsky&Pluess,2013b)。
2.3.3 单胺氧化酶A基因(MAOA)
神经递质代谢酶单胺氧化酶A基因,位于X染色体上,编码产生MAOA酶,进而产生与人们活动状态相关的神经递质。与差别易感性模型的假设相一致,相关的研究发现,拥有MAOA活性基因的个体当遭遇逆境时会产生最多的问题行为,而在非逆境中则会有最少的问题行为,表现出最大的可塑性。例如,Caspi等人(2002)的研究为MAOA基因对童年期虐待和成年后的反社会行为之间关系的调节作用提供了相关证据,遭受过童年期虐待且携带低MAOA活性基因的男性会表现出高暴力倾向,而对于那些高MAOA活性基因携带者童年期虐待的影响则很小。国内侯金芹、李新影、杨小冬和陈祉妍(2010)的研究也发现,携带短链MAOA基因的男孩对不良的家庭教养环境更敏感,更容易出现不良的问题行为,而长链MAOA携带者在不良的家庭教养环境中则较少出现外化问题。
以上研究都是基于单个基因和环境的交互作用,事实上一个人可能携带多个可塑性等位基因,这些携带多个可塑性等位基因的个体可能表现出对环境更高的易感性(Sonuga-Barke et al.,2009)。例如,Beaver和Belsky(2012)对教养代际传递的研究发现,个体携带可塑性基因越多,其青少年时期的教养经历越能预测其未来的教养行为,这为差别易感性假说提供了多基因型的证据。一项对5-羟色胺转运体基因、多巴胺受体基因和单胺氧化酶基因等三个基因型的研究也支持了差别易感性观点。在预测攻击行为时,消极环境条件与累计可塑性基因指数产生了交互作用,累积可塑性基因指数越高的个体,在不利环境中会出现更多的侵犯行为,同时在良好的社会环境中出现更少的侵犯行为(Simons et al.,2012)。还有研究发现,当成长于消极的教养环境中时,累计遗传指数高的青少年会对他们的恋人表现出更多的敌意,但是当他们成长于积极的家庭教养环境中时,这些青少年成年后会对恋人表现出更少的敌意(Masarik et al.,2014)。
3 本领域研究的局限性和未来研究的展望
长期以来,由于受到素质压力或双重风险模型的广泛影响,人们更多地关注那些更易受到消极养育环境不利影响的“脆弱”儿童,而忽视了这些儿童同时更容易受到积极的、支持性养育环境的有利影响的可能性,直到差别易感性模型的提出这一趋势才得到改变。这一模型为我们理解个体发展的可塑性提供了有价值的参考。但是,目前关于儿童对养育经历差别易感性的研究仍然存在一些问题和局限性。
第一,虽然到目前为止支持差别易感性的研究很多,但也有相当一部分研究并不符合差别易感性模型,这就可能涉及到一个问题:是否有些儿童只对逆境具有易感性,有些儿童只对积极的、支持性环境具有易感性,而有些儿童对两者都有或都没有易感性?同时,如何区分这些易感性不同的四类儿童群体?还有,目前研究者使用行为、生理和基因等不同的标志来区分更低或更高的易感性个体,但高消极情绪性、高生理反应性以及5-HTTPLR短等位基因携带者是否可能是同一个体,这一点是需要我们考虑到的(Auerbach et al.,1999)。
第二,为什么有些儿童比其他儿童对养育环境更具易感性?原因可能是这些个体拥有高度敏感的神经系统,使得他们比其他儿童有更强的易感性和反应性,对养育环境有更低的反应下限。当然,这也不能排除儿童对养育环境的高度易感性本身可能就是由其养育经历诱导的(Boyce&Ellis,2005)。对这一问题未来需要更多的研究加以探讨。
第三,儿童的皮质醇反应是否是环境影响的调节因素。虽然已有许多研究将皮质醇反应作为差别易感性的生理指标之一,但目前关于皮质醇反应是否确切是环境影响的调节变量仍然存在争议,有人认为皮质醇反应水平是环境因素的结果变量,而不是环境影响的调节因素(Fernald et al.,2008;Gunnar&Quevedo,2007)。例如,有研究表明一些年轻哺乳动物的皮质醇指数是由胎儿经历塑造的(Gluckman&Hanson,2005)。
最后,关于差别易感性的基因和环境交互作用的研究仍是一个难题。基因与环境交互作用的机制是怎样的,如何区分出具有某种“风险基因”的个体,某种“风险基因”的携带者与某种发展结果之间是否有确切的联系,多个可塑性基因累计遗传性的测量等问题都需要进一步的探讨(Belsky&Fearon,2002)。
尽管相对于素质压力或双重风险理论来说,差别易感性理论确实考虑到了对积极和消极环境的影响同时具有高易感性的儿童,但是差别易感性理论本身也存在着一定的局限性,它并未解释为什么一些儿童比其他儿童在积极环境中发展得更好。然而有研究表明确实存在一些具有“优势敏感性”特质的儿童,他们相比于那些具有“优势阻力”特质的儿童更容易受到积极环境或经历的有利影响,而对消极或中性环境来说并不敏感(Pluess&Belsky,2013)。尽管差别易感性理论存在着自身的局限性,并且未来研究中也依然面临着诸多的挑战性问题,但差别易感性模型确实为我们理解儿童发展的个体差异提供了一个独特而崭新的视角。鉴于目前我国该领域的研究很少,笔者希望可以将差别易感性模型更好地引入我国家庭养育环境对儿童发展影响的研究中,冀望为我国的儿童养育实践提供理论指导。
侯金芹,李新影,杨小冬,陈祉妍.(2010).父母教养与外化行为关系的行为遗传学研究.中国临床心理学杂志,18(6),742–744,795.
梁宗保,孙铃,张光珍,陈会昌,张萍.(2011).父亲情绪表达与儿童社会适应:气质的调节作用.心理发展与教育,27(4),351–358.
聂瑞虹,许颖,韩卓.(2015).皮质醇日常节律与儿童问题行为及心理社会因素的关系.心理科学进展,23(4),591–601.
张卫,甄霜菊.(2013).发展可塑性的个体差异——基因与环境的交互作用.华南师范大学学报(社会科学版),(5),106–111.
Auerbach,J.,Geller,V.,Lezer,S.,Shinwell,E.,Belmaker,R.H.,Levine,J.,&Ebstein,R.(1999).Dopamine D4 receptor(D4DR)and serotonin transporter promoter(5-HTTLPR)polymorphisms in the determination of temperament in 2-month-old infants.Molecular Psychiatry,4,369–373.
Bakermans-Kranenburg,M.J.,&van IJzendoorn,M.H.(2006).Gene-environment interaction of the dopamine D4 receptor(DRD4)and observed maternal insensitivity predicting externalizing behaviorin preschoolers.Developmental Psychobiology,48,406–409.
Bakermans-Kranenburg,M.J.,&van IJzendoorn,M.H.(2007).Genetic vulnerability or differential susceptibility in child development:The case of attachment.Journal of Child Psychology and Psychiatry,48,1160–1173.
Bakermans-Kranenburg,M.J.,&van IJzendoorn,M.H.(2011).Differential susceptibility to rearing environment depending on dopamine-related genes:New evidence and a meta-analysis.Development and Psychopathology,23,39–52.
Bakermans-Kranenburg,M.J.,van IJzendoorn,M.H.,Pijlman,F.T.A.,Mesman,J.,&Juffer,F.(2008).Experimental evidence for differential susceptibility:Dopamine D4 receptor polymorphism(DRD4 VNTR)moderates intervention effects on toddlers’externalizing behaviorin a randomized controlled trial.Developmental Psychology,44,293–300.
Beaver,K.M.,& Belsky,J.(2012).Gene-environment interaction and the Intergenerationaltransmission of parenting:Testing the differential-susceptibility hypothesis.Psychiatric Quarterly,83,29–40.
Belsky,J.(1997a).Theory testing,effect-size evaluation,and differential susceptibility to rearing influence:The case of mothering and attachment.Child Development,68,598–600.
Belsky,J.(1997b).Variation in susceptibility to environmental influence:An evolutionary argument.Psychological Inquiry,8,182–186.
Belsky,J.(2005).Differentialsusceptibility to rearing influences:An evolutionary hypothesis and some evidence.In B.Ellis&D.Bjorklund(Eds.),Origins of the social mind:Evolutionary psychology and child development(pp.139–163).New York.NY:Guildford.
Belsky,J.,Bakermans-Kranenburg,M.J.,&van IJzendoorn,M.H.(2007).For better and for worse:Differential susceptibility to environmental influences.Current Directions in Psychological Science,16,300–304.
Belsky,J.,& Fearon,R.M.(2002).Infant-mother attachment security,contextual risk,and early development:A moderational analysis.Development and Psychopathology,14,293–310.
Belsky,J.,Hsieh,K.H.,&Crnic,K.(1998).Mothering,fathering,and infant negativity as antecedents of boys’externalizing problems and inhibition at age 3 years:Differential susceptibility to rearing experience?Development and Psychopathology,10(2),301–319.
Belsky,J.,Newman,D.A.,Widaman,K.F.,Podkin,P.,Pluess,M.,Fraley,R.C.,… Roisman,G.(2015).Differential susceptibility to effects of maternal sensitivity?A study of candidate plasticity genes.Development and Psychopathology,27,725–746.
Belsky,J.,&Pluess,M.(2009a).Beyond diathesis stress:Differential susceptibility to environmental influences.Psychological Bulletin,135,885–908.
Belsky,J.,&Pluess,M.(2009b).The nature(and nurture?)of plasticity in early human development.Perspectives on Psychological Science,4,345–351.
Belsky,J.,&Pluess,M.(2013a).Beyond risk,resilience,and dysregulation:Phenotypic plasticity and human development.Development and Psychopathology,25,1243–1261.
Belsky,J.,&Pluess,M.(2013b).Genetic moderation of early child-care effects on social functioning across childhood:A developmentalanalysis.Child Development,84(4),1209–1225.
Belsky,J.,Pluess,M.,&Widaman,K.F.(2013).Confirmatory and competitive evaluation of alternative gene-environment interaction hypotheses.Journal of Child Psychology and Psychiatry,54,1135–1143.
Boyce,W.T.,Chesney,M.,Alkon,A.,Tschann,J.M.,Adams,S.,Chesterman,B.,....Wara,D.(1995).Psychobiologic reactivity to stress and childhood respiratory illnesses:Results of two prospective studies.Psychosomatic Medicine,57,411–422.
Boyce,W.T.,&Ellis,B.J.(2005).Biological sensitivity to context:I.An evolutionary-developmental theory of the origins and functions of stress reactivity.Development and Psychopathology,17,271–301.
Bradley,R.H.,&Corwyn,R.F.(2008).Infant temperament,parenting,and externalizing behavior in first grade:A test of the differential susceptibility hypothesis.Journal of Child Psychology and Psychiatry,49,124–131.
Caspi,A.,McClay,J.,Moffitt,T.E.,Mill,J.,Martin,J.,Craig,I.W.,....Poulton,R.(2002).Role of genotype in the cycle of violence in maltreated children.Science,297,851–854.
Caspi,A.,Sugden,K.,Moffitt,T.E.,Taylor,A.,Craig,I.W.,Harrington,H.,....Poulton,R.(2003).Influence of life stress on depression:Moderation by a polymorphism in the 5-HTT gene.Science,301,386–389.
Cicchetti,D.,&Garmezy,N.(1993).Prospects and promises in the study of resilience.Development and Psychopathology,5,497–502.
Conradt,E.,Measelle,J.,&Ablow,J.C.(2013).Poverty,problem behavior,and promise:Differential susceptibility among infants reared in poverty.Psychological Science,24(3),235–242.
Crockenberg,S.C.,& Leerkes,E.M.(2005).Infant temperament moderates associations between childcare type and quantity and externalizing and internalizing behaviors at 2½ years.Infant Behavior&Development,28,20–35.
Cummings,E.M.,El-Sheikh,M.,Kouros,C.D.,&Keller,P.S.(2007).Children’s skin conductance reactivity as a mechanism of risk in the context of parental depressive symptoms.Journal of Child Psychology and Psychiatry,48,436–445.
Eisenberg,N.,Sulik,M.J.,Spinrad,T.L.,Edwards,A.,Eggum,N.D.,Liew,J.,…Hart,D.(2012).Differential susceptibility and the early development of aggression:Interactive effects of respiratory sinus arrhythmia and environmental quality.Developmental Psychology,48(3),755–768.
Elisabeth de Neuf,C.(2012).Differential susceptibility to rearing influences:The role of infant autonomic functioning(Unpublished doctorial dissertation).University of Oregon.
Ellis,B.J.,Boyce,W.T.,Belsky,J.,Bakermans-Kranenburg,M.J.,& van IJzendoorn,M.H.(2011).Differential susceptibility to the environment:An evolutionary-neuro developmental theory.Development and Psychopathology,23,7–28.
El-Sheikh,M.,Harger,J.,&Whitson,S.M.(2001).Exposure to interparentalconflictand children’s adjustmentand physical health:The moderating role of vagal tone.Child Development,72,1617–1636.
El-Sheikh,M.,Keller,P.S.,&Erath,S.A.(2007).Marital conflict and risk for child maladjustment over time:Skin conductance level reactivity as a vulnerability factor.Journal of Abnormal Child Psychology,35,715–727.
Feldman,R.,Greenbaum,C.W.,&Yirmiya,N.(1999).Mother-infant affect synchrony as an antecedent of the emergence of self-control.Developmental Psychology,35,223–231.
Fernald,L.C.,Burke,H.M.,&Gunnar,M.R.(2008).Salivary cortisol levels in children of low-income women with high depressive symptomatology.Development and Psychopathology,20,423–436.
Gluckman,P.,& Hanson,M.(2005).The fetal matrix:Evolution,development and disease.Cambridge,England:Cambridge University Press.
Goforth,A.N.,Pham,A.V.,&Carlson,J.S.(2011).Diathesis-stress model.Inencyclopedia of child behavior and development(pp.502–503).US:Springer.
Groeneveld,M.G.,Vermeer,H.J.,van IJzendoorn,M.H.,&Linting,M.(2012).Stress,cortisol and well-being of caregivers and children in home-based child care:A case for differential susceptibility.Child:Care,Health and Development,38(2),251–260.
Gunnar,M.,&Quevedo,K.(2007).The neurobiology of stress and development.Annual Review of Psychology,58,145–173.
Hankin,B.L.,Nederhof,E.,Oppenheimer,C.W.,Jenness,J.,Young,J.F.,Abela,J.R.Z.,…Oldehinkel,A.J.(2011).Differential susceptibility in youth:Evidence that 5-HTTLPR x positive parenting is associated with positive affect‘for better and worse’.Translational Psychiatry,1,e44.
Karg,K.,Burmeister,M.,Shedden,K.,&Sen,S.(2011).The serotonin transporter promoter variant(5-HTTLPR),stress,and depression meta-analysis revisited:Evidence of genetic moderation.Archives of General Psychiatry,68,444–454.
Martin,C.G.,Kim,H.K.,Bruce,J.,&Fisher,P.A.(2014).Child diurnal cortisol rhythms,parenting quality,and externalizing behaviors in preadolescence.Psychoneuroendocrinology,40,170–180.
Masarik,A.S.,Conger,R.D.,Donnellan,M.B.,Stallings,M.C.,Martin,M.J.,Schofield,T.J.,…Widaman,K.F.(2014).For better and for worse:Genes and parenting interact to predict future behavior in romantic relationships.Family Psychology,28(3),357–367.
Morris,A.S.,Silk,J.S.,Steinberg,L.,Sessa,F.M.,Avenevoli,S.,&Essex,M.J.(2002).Temperamental vulnerability and negative parenting as interacting predictors of child adjustment.Journal of Marriage and Family,64,461–471.
Nikitopoulos,J.,Zohsel,K.,Blomeyer,D.,Buchmann,A.F.,Schmid,B.,Jennen-Steinmetz,C.,…Laucht,M.(2014).Are infants differentially sensitive to parenting?Early maternal care,DRD4genotype and externalizing behavior during adolescence.Journal of Psychiatric Research,59,53–59.
Obradović,J.,Bush,N.R.,Stamperdahl,J.,Adler,N.E.,&Boyce,W.T.(2010).Biological sensitivity to context:The interactive effects of stress reactivity and family adversity on socioemotional behavior and school readiness.Child Development,81(1),270–289.
Pluess,M.,&Belsky,J.(2009).Differential susceptibility to rearing experience:The case of childcare.Journal of Child Psychology and Psychiatry,50(4),396–404.
Pluess,M.,&Belsky,J.(2010).Differential susceptibility to parenting and quality child care.Developmental Psychology,46(2),379–390.
Pluess,M.,& Belsky,J.(2013).Vantage sensitivity:Individual differences in response to positive experiences.Psychological Bulletin,139(4),901–916.
Pluess,M.,Belsky,J.,Way,B.M.,&Taylor,S.E.(2010).5-HTTLPR moderates effects of current life events on neuroticism:Differential susceptibility to environmental influences.Progressin Neuro-Psychopharmacology &Biological Psychiatry,34,1070–1074.
Poehlmann,J.,Hane,A.,Burnson,C.,Maleck,S.,Hamburger,E.,&Shah,P.E.(2012).Preterm infants who are prone to distress:Differential effects of parenting on 36-month behavioral and cognitive outcomes.Journal of Child Psychology and Psychiatry,53(10),1018–1025.
Roisman,G.I.,Newman,D.A.,Fraley,R.C.,Haltigan,J.D.,Groh,A.M.,&Haydon,K.C.(2012).Distinguishing differential susceptibility from diathesis-stress:Recommendations for evaluating interaction effects.Development and Psychopathology,24,389–409.
Rosenthal,D.(1963).A suggested conceptual framework.In D.Rosenthal(Ed.),The Genain quadruplets(pp.505–516).New York:Basic Books.
Rothbart,M.K.,&Bates,J.E.(2006).Temperament.In N.Eisenberg,W.Damon,&R.M.Lerner(Eds.),Handbook of child psychology:Vol.3.Social,emotional,and personality development(6th ed.,pp.99–166).Hoboken,NJ:Wiley.
Sheese,B.E.,Voelker,P.M.,Rothbart,M.K.,&Posner,M.I.(2007).Parenting quality interacts with genetic variation in dopamine receptor D4 to influence temperament in early childhood.Development and Psychopathology,19,1039–1046.
Sigelman,C.K.,&Rider,E.A.(2009).Developmental psychopathology.InLife-span human development(6th ed.,(pp.468–495).Belmont,CA:Wadsworth Cengage Learning.
Simons,R.L.,Lei,M.K.,Stewart,E.A.,Beach,S.R.H.,Brody,G.H.,Philibert,R.A.,&Gibbons,F.X.(2012).Social adversity,genetic variation,street code,and aggression:A genetically informed model of violent behavior.Youth Violence and Juvenile Justice,10(1),3–24.
Sonuga-Barke,E.J.S.,Oades,R.D.,Psychogiou,L.,Chen,W.,Franke,B.,Buitelaar,J.,....Faraone,S.V.(2009).Dopamine and serotonin transporter genotypes moderate sensitivity to maternal expressed emotion:The case of conduct and emotional problems in attention deficit/hyperactivity disorder.Journal of Child Psychology and Psychiatry,50(9),1052–1063.
van IJzendoorn,M.H.,Belsky,J.,&Bakermans-Kranenburg,M.J.(2012).Serotonin transporter genotype 5HTTLPR as a marker of differential susceptibility?A meta-analysis of child and adolescent gene-by-environment studies.Translational Psychiatry,2,e147.
Velderman,M.K.,Bakermans-Kranenburg,M.J.,Juffer,F.,&van IJzendoorn,M.H.(2006).Effects of attachmentbased interventions on maternal sensitivity and infant attachment:Differential susceptibility of highly reactive infants.Journal of Family Psychology,20,266–274.
Volling,B.L.,&Feagans,L.V.(1995).Infant day care and children’s social competence.Infant Behavior&Development,18,177–188.
Widaman,K.F.,Helm,J.L.,Castro-Schilo,L.,Pluess,M.,Stallings,M.C.,& Belsky,J.(2012).Distinguishing ordinal and disordinal interactions.Psychological Methods,17,615–622.
Windhorst,D.A.,Mileva-Seitz,V.R.,Linting,M.,Hofman,A.,Jaddoe,V.W.V.,Verhulst,F.C.,…Bakermans-Kranenburg,M.J.(2015).Differential susceptibility in a developmental perspective:DRD4 and maternal sensitivity predicting externalizing behavior.Developmental Psychobiology,57,35–49.
Wolff,B.C.,Wadsworth,M.E.,Wilhelm,F.H.,&Mauss,I.B.(2012).Children's vagal regulatory capacity predicts attenuated sympathetic stress reactivity in a socially supportive context:Evidence for a protective effect of the vagal system.Development&Psychopathology,24(2),677–689.
猜你喜欢
昆明医科大学学报(2021年6期)2021-07-31
启蒙(3-7岁)(2019年8期)2019-09-10
幸福(2019年21期)2019-08-20
幸福(2018年33期)2018-12-05
妈妈宝宝(2018年9期)2018-12-05
中华家教(2018年10期)2018-10-30
创新作文·初中版(2017年6期)2017-12-06
医学研究杂志(2015年6期)2015-07-01
医学研究杂志(2015年8期)2015-06-22
中国当代医药(2015年33期)2015-03-01
