
视觉适应及其神经机制*
2015-02-05 01:51忆鲍
心理科学进展 2015年7期
高 忆鲍 敏
(1中国科学院行为科学重点实验室,中国科学院心理研究所,北京 100101)(2中国科学院大学,北京 100039)
人眼感受到的外界环境是不断变化的,我们的视觉系统必须根据外界环境的变化做出适应性的调整,这就是视觉适应(visual adaptation)(Clifford et al.,2007)。视觉适应是一种广泛存在的视觉现象,比如盯着下落的瀑布看一段时间后再去看旁边静止的岩石会觉得岩石在向上“飞”,这就是经典的瀑布后效(waterfall aftereffect)。我们从露天进入关灯的电影院时会感觉一片漆黑,需要一段时间之后才能逐渐看清电影院里的人,这种现象属于暗适应(dark adaptation);当我们从电影院走出来时,一瞬间感觉阳光十分刺眼,但很快就恢复正常,这是明适应(light adaptation)的过程。上述的例子分别属于运动适应(motion adaptation)和亮度适应(luminance adaptation)。
视觉适应最显著的影响是能够改变我们对物体外观的知觉,有时甚至产生视错觉。因此,视觉适应的研究对于理解神经系统如何加工外部视觉信息具有重要的现实意义。此外,视觉适应也是研究大脑可塑性的重要手段之一。一般认为,视觉适应是短时程可塑性的一个代表(Kohn,2007,例外见 Bao&Engel,2012),其他形式的可塑性有长时程的知觉学习以及脑损伤后的功能恢复等。研究视觉适应有助于理解大脑可塑性。
近年来,ERP、TMS、fMRI等技术越来越被研究者们广泛使用,这些技术手段结合行为实验以及电生理实验的结果使得人们对视觉适应的认识逐步深入。本文从视觉适应发生的神经机制、理论模型以及不同时间尺度的控制机制这几个方面来综述视觉适应的研究现状,并对未来的研究方向进行展望。
1 视觉适应的神经机制
1.1 亮度适应和对比度适应
白天的光照强度可能是夜晚的几百万倍,人眼必须适应性地对具有巨大光强差异的外周环境进行反应,这就是亮度适应。视网膜的基本功能之一就是适应亮度。视网膜上神经元的反应范围是有限的,亮度适应使得神经元通过改变自身反应的增益和时间进程来适应外界环境中的亮度变化,使神经元以外周环境的平均亮度为中心进行反应,从而使视觉系统保持对外界环境的敏感性(Rieke&Rudd,2009)。
对比度是指图像中所有亮度相对于图像平均亮度的变异程度。对比度能够帮助我们分辨刺激的朝向、运动方向、空间频率等特征。对比度适应(contrast adaptation)是指在观察某个高对比度的刺激一段时间之后对相似的低对比度刺激的敏感性降低(Duong&Freeman,2007;Kohn,2007),它是生活中最为常见的一种适应形式。许多研究都表明视网膜上的神经元不仅能够适应各种亮度,也能够适应不同对比度(Baccus&Meister,2002;Demb,2008)。研究者利用图形视网膜电图(pattern electroretinogram,PERG)记录人的视网膜神经节细胞的反应,结果发现,对比度适应除了使得反应振幅减弱外还能够减少反应的潜伏期(Heinrich&Bach,2001)。Brown和Masland(2001)利用单细胞记录(single cell recording)技术对兔子视网膜的研究表明,双极细胞似乎是对比度适应发生的可能位置。
视网膜之后、视觉皮层之前的外侧膝状体(lateral geniculate nucleus,LGN)内的神经元也能够适应对比度。在麻醉猫的外侧膝状体中,Y细胞比X细胞的适应程度更大,但是Y细胞的对比度适应受到皮层反馈的影响,而X细胞的对比度适应不受其影响(Li,Ye,Song,Yang,&Zhou,2011)。这两类细胞功能的显著差异在于空间信息(例如空间频率)主要由X细胞传递,时间特征(例如运动)主要由Y细胞传递(Wright&Ikeda,1974)。类似地,猕猴LGN内对视觉运动和轮廓敏感的大细胞(magnocellular cell)也存在对比度适应,并且这种作用很可能承袭自视网膜,而对颜色和细节敏感的小细胞(parvocellular cell)则几乎不存在对比度适应(Solomon,Peirce,Dhruv,&Lennie,2004)。研究表明,猕猴LGN中背侧小细胞层中的细胞类似猫LGN中的X细胞,腹侧大细胞层中的细胞类似猫LGN中的Y细胞(Dreher,Fukada&Rodieck,1976;Sherman,Wilson,Kaas&Webb,1976;Schiller&Malpeli,1978)。这两类细胞适应程度的差别为研究从两类细胞到皮层区域输入信号的相对强度提供了依据。视觉加工通路中一个阶段的适应效果可能受之前加工阶段的影响,也可能受到随后阶段的反馈调节,如何将不同加工阶段的适应效果单独区分出来是值得进一步研究的问题。
视觉信息加工分为不同的层级,从较低级的视皮层逐渐传递到较高级的视皮层,在这个通路中神经元的感受野逐渐变大,并且越是高级的视皮层对诸如视网膜位置、朝向、空间频率等基本刺激属性越不敏感,而对复杂刺激属性则更加敏感(Desimone&Ungerleider,1989)。研究者们常用适应效果的迁移性(transfer)与特异性(specificity)来探求视觉适应的发生阶段。初级视觉皮层(V1)具有大量的单眼神经元,而且对视网膜位置、空间频率、朝向等基本属性的调谐(tuning)的带宽较窄,亦即这里的神经元相当多的具有更为局限的感受野。因此,如果适应的效果完全不发生迁移,例如仅在适应的眼或者适应的刺激的基本属性上可以观察到,那就说明该适应仅发生在初级视觉皮层及更早的加工阶段(例如外侧膝状体、视网膜等)。反之,如果适应的效果百分之百地发生了迁移,则说明适应发生在较晚的视觉加工阶段。研究发现,当适应刺激和探测刺激朝向相同时,探测刺激的对比度侦察阈限提高最多(Fang,Murray,Kersten,&He,2005)。当适应刺激和探测刺激呈现在不同眼时,适应效果仍然存在,但相比呈现在同眼时更弱(Baker&Meese,2012)。单细胞记录发现,成年猫和猕猴的初级视觉皮层的适应具有朝向选择性(Carandini,Movshon,&Ferster,1998;Dragoi,Sharma,&Sur,2000)。这些发现都说明初级视觉皮层在对比度适应中起到了十分重要的作用,但迁移的发生说明对比度适应不仅仅发生在初级视觉皮层。Boynton和 Finney(2003)运用fMRI技术研究发现,在对光栅刺激适应以后,视觉区域对朝向相同或垂直的探测光栅的反应都会减弱,而且减弱程度在V1和V2没有显著差别,而在视觉通路的更高级区域V3和V4V则差异显著,这表明V3和V4V在对比度适应的朝向选择性中也起着重要作用。从以上研究可以发现,视网膜、LGN和视皮层都存在着对比度适应。
1.2 颜色适应
尽管外界环境中的光照强度和光谱组成在变化,视觉系统对颜色的知觉始终保持相对不变,这就是颜色恒常性(color constancy)(Foster,2011)。例如,不管是明亮的晴天还是昏暗的阴天,我们总是认为一个熟透的苹果是红色的。从一个更长的时间尺度上来说,老年人视网膜对光谱的感受性相对于其青年时期已经发生了变化,但是老年人依旧会知觉到书本的纸张是白色的(Wuerger,2013)。颜色恒常性是颜色适应(color adaptation)的表现形式之一。
视网膜上存在分别对光谱中的长波、中波、短波最敏感(即分别对红色、绿色、蓝色最敏感)的三种视锥细胞(Nathans,Thomas,&Hogness,1986),这是我们知觉到不同颜色的生理基础。颜色适应包含了对亮度的适应和对对比度的适应,亮度适应能够调节对环境中平均亮度和颜色的敏感性,而对比度适应通过改变对颜色空间中不同方向上对比度的知觉来影响我们的颜色知觉(Webster,1996)。在视网膜感受器细胞之后,来自视网膜的颜色信号以红-绿、黄-蓝、明-暗的拮抗形式进行传递和加工(Krauskopf,Williams,&Heeley,1982)。单细胞记录发现,猕猴的LGN以及V1内存在以拮抗形式传递颜色敏感信息的神经元(Derrington,Krauskopf,&Lennie,1984;Conway,2001)。fMRI研究发现,在适应红-绿光栅后,初级视觉皮层对红-绿光栅的激活小于对明-暗光栅的激活;类似地,在适应明-暗光栅后,对明-暗光栅的激活小于对红-绿光栅的激活。这说明初级视觉皮层上存在能够进行颜色适应的神经元(Engel&Furmanski,2001)。另外,单眼颜色适应的效果能部分迁移到另一只眼睛,表明大脑视觉皮层参与了颜色适应(Neitz,Carroll,Yamauchi,Neitz,&Williams,2002)。尽管关于颜色适应的神经机制研究还不是很多,但根据已有研究可以发现,视网膜、视网膜感受器细胞之后的拮抗机制以及皮层机制都在颜色适应中起作用。
1.3 运动适应
运动适应是指长时间观察向某个方向运动的物体之后会损害运动检测任务的成绩,比如运动方向辨别、速率估计等(Morgan,2012;McGovern,Roach,&Webb,2012),并且会产生运动错觉。由运动适应产生的错觉之一是运动后效(motion aftereffect,MAE)。用不同类型的探测刺激来测量运动适应程度的大小能够使我们了解运动适应发生的阶段。用静态探测刺激测得的是静态运动后效(static motion aftereffect,sMAE),用动态探测刺激测得的是动态运动后效(dynamic motion aftereffect,dMAE)。sMAE只能部分迁移到另一只眼睛(Moulden,1980),这说明至少有部分单眼神经元参与sMAE,视觉加工的较低级区域的运动感受器存在着运动适应;而dMAE的眼间迁移则分不同情况,当刺激呈现在中央视野并且被试注意适应刺激时dMAE具有完全的眼间迁移,而当刺激呈现在外周视野并且在适应的同时被试完成干扰任务时dMAE只有部分的眼间迁移(Nishida,Ashida,&Sato,1994;Nishida&Sato,1995),这说明dMAE包含两种成分:较为低级的对单眼运动感受器的适应以及较为高级的对双眼刺激敏感的运动感受器的适应。当适应刺激是辐射状时,探测刺激和适应刺激分别呈现在注视点的两侧仍能产生“虚幻”的运动后效,而平移运动的适应刺激则不能,这种后效被认为可能产生于感受野较大且对辐射或旋转刺激敏感的颞中区(middle temporal area,MT)或内侧颞区(medial superior temporal area,MST)(Meng,Mazzoni,&Qian,2006)。许多单细胞记录研究都发现猴子MT神经元存在具有朝向选择性的运动适应(Petersen,Baker,&Allman,1985;van Wezel&Britten,2002)。然而,Kohn和Movshon(2003)发现,运动适应的效果具有空间位置特异性,而MT细胞感受野较大,不足以区分如此精细的位置,表明适应效果有可能承袭自感受野较小的V1。这暗示着在视觉加工较高级区域发现的运动适应效果可能受到较低级脑区适应的影响。利用fMRI技术发现,对一阶运动(由图像亮度的时空变化引起,可以由视觉系统的简单的“运动感受器”加工,这些“感受器”觉察到视网膜上一点的亮度改变,然后将其与一个短时间隔后视网膜上临近的一点上的亮度改变关联起来进行分析)和二阶运动(运动轮廓由图像的其他属性例如对比度、图案、闪烁等决定,且这些并不会造成上述的一阶运动)的适应都能显著减少诸如V1、V2、V3、VP、V3A、V4v以及MT在内的广大视觉区域的BOLD信号强度(Seiffert,Somers,Dale,&Tootell,2003;Nishida,Sasaki,Murakami,Watanabe,&Tootell,2003)。综上所述,视觉加工通路较低级到较高级的多个脑区都存在运动适应(Mather,Pavan,Campana,&Casco,2008)。
1.4 面孔适应
不同于对比度、颜色等,面孔是具有复杂特征的刺激,它是我们赖以辨别人的性别、表情、种族等信息的根据。研究表明视觉系统对面孔的长宽比、表情以及性别、种族等都能产生适应(Strobach&Carbon,2013),这就是面孔适应(face adaptation)。面孔适应会产生面孔扭曲后效(face distortion aftereffect,FDAEs),例如,观察一个被横向拉伸的面孔一段时间之后会感觉这个面孔不那么扭曲了,并且会认为正常的面孔向反方向扭曲(Zhao&Chubb,2001)。面孔适应还会产生面孔身份后效(face identity aftereffects,FIAEs),这种后效基于对面孔空间(在多维度上利用计算模型改变距离面孔原型的距离)中与面孔原型有不同距离面孔的适应(Valentine,2001),并被证明能改善对适应维度上面孔的识别(Leopold,O’Toole,Vetter,&Blanz,2001)。
与视觉加工低级阶段的适应相比,许多研究都发现对面孔的适应效果能在不同位置、朝向、年龄以及图像大小,甚至是不同性别之间迁移(Kovács,Zimmer,Harza,Antal,&Vidnyánszky,2005;Watson&Clifford,2003;Zhao&Chubb,2001;O’Neil&Webster,2011)。用同一个人的不同照片去测面孔适应后效发现这种后效仍然存在(Fox&Barton,2007),这些发现表明视觉加工的高级阶段在面孔适应中起着重要作用。重复观看相同面孔相对于观看新面孔梭状回面孔区(fusiform face area,FFA)的BOLD信号强度显著减弱(Eger,Schyns,&Kleinschmidt,2004),改变适应面孔的位置或大小后这种BOLD信号强度的减弱仍然存在(Grill-Spector et al,1999),这说明FFA能产生面孔适应,并且这种适应与图像的低级特征无关。然而,有研究发现,对简单的、本身并不像面孔的曲线的适应也能影响对知觉到的面孔表情的判断(Xu,Dayan,Lipkin,&Qian,2008;Dickinson&Badcock,2013),这让研究者们推测面孔适应也可能继承自视觉加工的低级阶段。综上所述,研究者们认为面孔适应至少部分取决于视觉加工高级阶段反应的变化(Webster&Macleod,2011)。
不止对面孔,视觉系统对动物、树木等日常生活中出现的复杂刺激也存在适应后效(Daelli,van Rijsbergen,&Treves,2010)。由此看来,视觉系统不仅对基本的刺激属性如亮度、对比度、运动、颜色等存在适应,对复杂的高级的刺激输入也存在适应。这暗示着视觉适应可能是一种普遍存在的机制,是视觉系统的内在功能。
2 视觉适应的理论模型
视觉适应最显著的效应就是会影响对随后呈现刺激的知觉。首先,视觉适应会导致知觉偏差(Levinson&Sekuler,1976),上述提到的运动后效就是适应导致知觉错觉的典型例子。其次,适应能改变人们辨别刺激强度的能力:适应能提高与适应刺激相似刺激的辨别能力(Oruç&Barton,2011),损害与适应刺激水平不相似刺激的辨别能力(Phinney,Bowd,&Patterson,1997)。可以说,视觉系统如何适应决定着个体对外部世界的最终知觉。视觉适应究竟如何改变神经元的反应?已有研究提出了疲劳和再标准化这两种假说。
2.1 “疲劳”假说
“疲劳”假说认为,视觉适应的本质是神经元的“疲劳”(fatigue)。视觉系统存在许多对刺激属性反应的通道,每个通道对特定刺激水平敏感,这些通道的调谐范围都很窄。刺激由许多不同通道的总反应性来表征,视觉适应导致对适应刺激敏感的通道中神经元发放“疲劳”,也就是使这些神经元的反应相对于未适应时减弱,从而减少了对该种刺激属性加工“通道”的敏感性(Graham,1989;Sekuler&Pantle,1967)。研究表明,这种“疲劳”的本质是皮层神经元的超级化(Sanchez-Vives,Nowak,&McCormick,2000)。“疲劳”假说最直接的证据来自单细胞记录技术对猫的研究,研究发现,适应高对比度光栅刺激一段时间后,初级视觉皮层(V1)相应区域的神经元反应减弱(Maffei,Fiorentini,&Bisti,1973)。利用fMRI技术的研究发现,适应时初级视皮层和纹外皮层相关区域的BOLD信号强度减弱,且适应导致的BOLD信号强度的减少程度取决于探测刺激和适应刺激之间的角度差,探测刺激与适应刺激朝向相同时信号强度减少最多(Fang et al.,2005)。这里不同于Boynton等人(Boynton et al.,2003)在初级视觉皮层未发现朝向选择性适应的结果,这可能是由于两者的适应时长不一样(Fang et al.,2005),Boynton等人的实验中只适应了1 s而Fang等人的实验中适应了20 s,这说明各个区域适应的程度与适应的时长有关。疲劳理论可以解释对单一刺激水平如空间频率、对比度、朝向等的适应。以对某个朝向的适应为例,“疲劳”假说可以用来解释“倾斜后效”(tilt aftereffect),即观察向某个朝向倾斜的线段一段时间之后再看竖直的线段觉得该线段向相反朝向倾斜。视觉适应减少对适应朝向敏感的神经元的反应,从而导致敏感性调谐的排斥性(repulsion)移动(Dragoi et al.,2000),因此,知觉到随后呈现的竖直刺激向与适应朝向相反的朝向倾斜。
然而,神经元“疲劳”假说既不足以解释非常短时间如67 ms内的适应(Glasser,Tsui,Pack,&Tadin,2011),也不足以解释长达几小时甚至几天的适应,因为神经元疲劳的发生既不能快速到在67ms的时间内发生,同时在几小时内也早已恢复。另一方面,既然“疲劳”的本质是是神经元的超极化,那么对不同探测刺激的反应应该相同,然而当探测刺激与适应刺激朝向相同时信号强度减少最多(Fang et al.,2005)违背了这一点,因此“疲劳”假说对视觉适应的解释已渐渐淡出主流研究观点。
2.2 再标准化假说
另一种观点认为视觉适应的机制是神经元反应的再标准化(renormalization)。该理论认为,视觉系统内部存在一个知觉模板,这个模板也就是场景中的平均刺激水平,刺激由距离模板的距离来表征,视觉适应使得知觉模板向当前刺激水平靠近。知觉模板在对自然界物体的表征中是普遍存在的。例如,在面孔适应中,适应一个男性化的面孔使得我们视觉系统内部的知觉模板向男性化的面孔移动,随后判断一个中性面孔的时候这个中性面孔距离知觉模板更远了,就会认为这个中性面孔更女性化(Webster,Kaping,Mizokami,&Duhamel,2004)。
再标准化理论可以用来解释刺激由多个加工宽调谐的(broadly tuned)“通道”来表征的情况。例如,在颜色适应中,适应红色会使知觉模板向红色移动,随后呈现一个白色背景的时候会认为这个白色背景是绿色的。颜色知觉中的知觉模板即白色,白色是三种调谐范围较宽的颜色通道反应相同时的情况,即外周环境的平均值。再标准化理论也可以用来解释对本身具有许多刺激强度的图像的适应。例如,自然场景中包含许多宽调谐的振幅谱,对自然场景的适应使得许多宽调谐通道的反应再标准化(Webster,2011)。
再标准化假说和“疲劳”假说对适应后效有不同的预测。以模糊适应(blur adaptation)为例,再标准化假说认为,适应模糊刺激使得视觉系统向知觉模板(norms)调整,这个模板受当前适应水平的影响,因此适应模糊刺激会使得随后呈现的模糊刺激看起来更清晰,而适应中性刺激本身则不会对随后呈现刺激的模糊程度的知觉产生影响;而根据“疲劳”假说,模糊适应使得视觉系统在适应的模糊等级附近敏感性降低,进而出现“排斥”现象,那么适应模糊程度为中性的刺激会使得视觉系统对中性刺激的反应减弱,从而使得模糊刺激知觉起来更模糊。研究发现,适应中性刺激不会改变对中性刺激的知觉,这支持了再标准化假说,适应中性刺激只是强化了已有的模板,不会改变对随后刺激的知觉。在此基础上进行的模型拟合结果也支持了再标准化假说而非“疲劳”假说(Elliott,Georgeson,&Webster,2011)。
3 视觉适应的时间尺度
外界刺激的变化有的转瞬即逝,有的则非常缓慢。同样地,视觉适应也能在多种时间尺度上发生,短到几秒钟,长到几个小时甚至几天。研究发现,对比度适应在50 ms内就能发生(Pavan,Marotti,&Campana,2012)。适应向某个方向运动的高对比度且尺寸较大的光栅67 ms足以影响对随后呈现的探测刺激方向的判断(Glasser et al.,2011),这说明适应刺激呈现非常短的时间也能产生适应效果。长时程适应的一个最常见的例子就是白内障患者的颜色适应(color adaptation),手术前晶状体上的黄色浑浊使得视网膜接收到的短波较少,在去除黄色浑浊后,到达视网膜的短波大幅增加,因此在完成消色差任务时被试需要用黄色去抵消蓝色使之看起来无色,几个月后消色差点才逐渐变成白色。这里视觉适应表现为颜色系统的长时程再标准化(Delahunt,Webster,Ma,&Werner,2004)。
近年来,一些巧妙的实验设计证明了长时程和短时程两种视觉适应时间机制的存在。麦考勒效应(McCollough effect)是一种经典的颜色适应后效,适应红色竖直光栅和绿色水平光栅使得随后呈现的竖直和水平中性光栅分别看起来呈现绿色和红色(McCollough,1965)。研究发现,麦考勒效应包含两种时间机制,适应效果在较短的时间内(约30s)形成和衰退反映了一种快速适应机制,而随着时间增加这种适应效果的建立和衰退达到渐进水平,并且不会完全饱和或消退,就像一个理想的积分器,这证明了视觉适应的慢速机制的存在(Vul,Krizay,&MacLeod,2008)。
Bao和Engel(2012)的研究进一步了提出视觉适应的多重机制理论。他们之前的研究发现,利用更改现实技术剥夺环境中的竖直信息4小时后,视觉系统对竖直信息响应的增益会被调高(Zhang,Bao,Kwon,He,&Engel,2009;Bao&Engel,2012)。若这时再观看未经过滤的拍摄图像15分钟,被作者称为“去适应”,视觉系统实际得到的竖直信号的输入应该比之前更强。根据视觉适应的多重机制预测,短时程机制检测到竖直的信号增强就会减低其控制的对竖直朝向敏感的神经元的增益;而15分钟不足以使得长时程机制根据这种变化作出增益调整,因此它们所控制的神经元仍然保持对竖直朝向的较强的增益。短时程机制和长时程机制的增益调节作用在15分钟的去适应阶段是相反的。需要注意的是,Vul等人对McCollough效应的模型拟合结果表明,朝向依赖的颜色适应(Orientation-contingent color adaptation)的快速和慢速机制是以并行而非串行方式起作用的(Vul et al.,2008)。因此这两种机制是相互独立的,适应效果可以线性相加,即总的适应效果等于二者之和。对比度适应的多重机制理论也遵循了这一规律,因此去适应阶段长短时程机制的相反输出互相抵消,使得去适应结束时观测不到明显的适应后效。而在进一步测试过程中,短时程机制造成的负的效应会随着时间消退,长时程机制的正的作用仍然存在,因此后测试中适应后效可以从无到有逐渐地显现出来,被称为“自然恢复”现象。多重机制理论可以预测这一自然恢复现象,而单一机制理论则不能预测这种现象。实验结果与多重机制理论的预测一致,从而证实了多重机制的存在(图 1)。
运用类似的实验逻辑和去适应范式,在更短的秒、分钟时长级别的对比度适应上,研究者也观测到了自然恢复现象(Bao,Fast,Mesik,&Engel,2013)。这就证明在几秒到几小时这样的很大的时间跨度上都是具有不同的控制机制影响适应的效果,这些机制对不同环境有各自偏好的速率,即具有不同的时间尺度。比较快速的那些机制可能应对的是环境中的短暂变化,它们满足了神经元对不同场景敏感性范围需要快速切换的需求,而那些慢速的机制应对的可能是环境中持久的改变。在运动适应和面孔适应的研究中,他们也发现了自然恢复现象(Mesik,Bao,&Engel,2013),这说明多重机制在许多不同类型的适应中都存在。因此,具有不同时间尺度的多重控制机制可能是视觉适应背后的一种普遍规律。
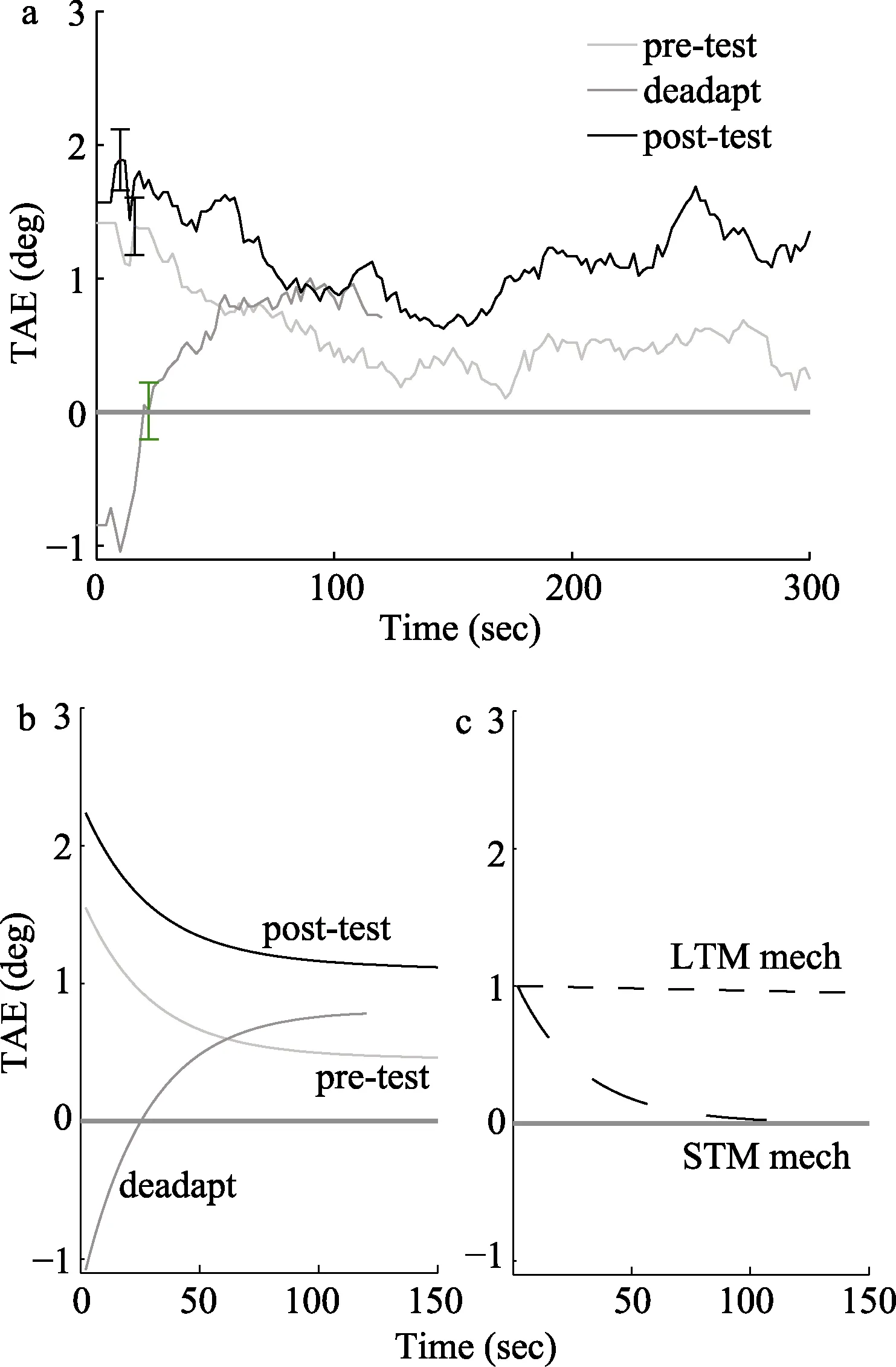
图1 多重机制控制对比度适应的结果图(Bao&Engel,2012)。(A)倾斜后效随时间变化的曲线,蓝色曲线代表出适应后的倾斜后效,绿色曲线代表去适应后的倾斜后效,红色曲线代表再适应后的倾斜后效(B)用双重机制模型来拟合数据可解释超过80%的变异。(C)拟合结果显示的两个机制的消退方程。
4 小结与展望
总体来说,视觉系统能够对输入视觉刺激的颜色、朝向、运动方向等基本刺激属性产生适应,也能对面孔等具有复杂特征的高级刺激产生适应,这表明视觉适应可能是视觉系统的内在功能。从视觉加工的低级脑区到高级脑区都存在视觉适应,且同一种类型的适应可能在不止一个脑区得到加工。针对不同的刺激类型,视觉适应的疲劳假说和再标准化假说都可以解释部分适应效果。然而越来越多的证据支持了再标准化假说,使得疲劳假说逐渐淡出了视觉适应的理论解释行列。
视觉适应可以由快速和慢速不同时间尺度的机制控制,这种由多重时间机制控制的适应在动作学习中是普遍存在的(Smith,Ghazizadeh,&Shadmehr,2006)。自然环境中有些变化持续时间长,有些变化则十分短暂,更为经济的是视觉系统在应对短暂变化的同时保留长时程环境下的行为特征(Kording,Tenenbaum,&Shadmehr,2007),这就需要大脑在应对短暂变化的同时保持对长时程环境中特征的记忆,对于先前学习的记忆能促进适应(Kojima,Iwamoto,&Yoshida,2004)。视觉适应的不同时间尺度背后所基于的神经机制将是未来该方向的研究重点,不同时间尺度的适应控制机制是发生在细胞层面,还是在不同的神经元群,亦或是视觉处理脑区之间,仍然是亟待回答的科学问题。
视觉系统在不同时间尺度上的适应反映了大脑可塑性的复杂性。作为大脑可塑性不同研究方面的视觉适应与知觉学习的关系有待探究。已有研究表明视觉适应和知觉学习存在着交互作用(Harris,Gliksberg,&Sagi,2012;Yehezkel,Sagi,Sterkin,Belkin,&Polat,2010;McGovern et al,2012)。但是这些研究的结果存在互相矛盾的地方。知觉学习和视觉适应之间具体存在哪些交叉,这些交叉与控制适应的不同时间尺度的机制之间又是何种关系,仍然不清楚。探究两者的交互作用对探究视觉可塑性的发生阶段以及神经机制有着重要的意义,这是未来研究的一个方向。
Baccus,S.A.,&Meister,M.(2002).Fast and slow contrast adaptation in retinal circuitry.Neuron,36,909–919.
Baker,D.H.,&Meese,T.S.(2012).Interocular transfer of spatial adaptation isweak at low spatial frequencies.Vision Research,63,81–87.
Bao,M.,&Engel,S.A.(2012).Distinct mechanism for long-term contrast adaptation.Proceedings of the National Academy of Sciences of the United States of America,109,5898–5903.
Bao,M.,Fast,E.,Mesik,J.,&Engel,S.A.(2013).Distinct mechanisms controlcontrastadaptation overdifferent timescales.Journal of Vision,13(10),1–11.
Boynton,G.M.,&Finney,E.M.(2003).Orientation-specific adaptation in human visual cortex.TheJournalof Neuroscience,23,8781–8787.
Brown,S.P.,&Masland,R.H.(2001).Spatial scale and cellular substrate of contrast adaptation by retinal ganglion cells.Nature Neuroscience,4,44–51.
Carandini,M.,Movshon,J.A.,&Ferster,D.(1998).Pattern adaptation and cross-orientation interactions in the primary visual cortex.Neuropharmacology,37,501–511.
Clifford,C.W.,Webster,M.A.,Stanley,G.B.,Stocker,A.A.,Kohn,A.,Sharpee,T.O.,&Schwartz,O.(2007).Visual adaptation:Neural,psychological and computational aspects.Vision Research,47,3125–3131.
Conway,B.R.(2001).Spatial structure of cone inputs to color cells in alert macaque primary visual cortex(V-1).The Journal of Neuroscience,21,2768–2783.
Daelli,V.,van Rijsbergen,N.J.,&Treves,A.(2010).How recent experience affects the perception of ambiguous objects.Brain Research,1322,81–91.
Delahunt,P.B.,Webster,M.A.,Ma,L.,&Werner,J.S.(2004).Long-term renormalization of chromatic mechanisms following cataract surgery.Visual Neuroscience,21,301–307.
Demb,J.B.(2008).Functional circuitry of visual adaptation in the retina.The Journal of Physiology,586,4377–4384.
Derrington,A.M.,Krauskopf,J.,& Lennie,P.(1984).Chromatic mechanisms in lateral geniculate nucleus of macaque.The Journal of Physiology,357,241–265.
Desimone,R.,&Ungerleider,L.G.(1989).Neural mechanisms ofvisualprocessing in monkeys.In F.Boller& J.Grafman(Eds.),Handbook of neuropsychology(Vol.2,pp.267–299).New York:Elsevier Science.
Dickinson,J.E.& Badcock,D.R.(2013).On the hierarchical inheritance of aftereffects in the visual system.Frontiers in Psychology,4,472.
Dragoi,V.,Sharma,J.,& Sur,M.(2000).Adaptationinduced plasticity of orientation tuning in adult visual cortex.Neuron,28,287–298.
Dreher,B.,Fukada,Y.,&Rodieck,R.W.(1976).Identification,classification and anatomical segregation of cells with X-like and Y-like propertiesin the lateralgeniculate nucleus of old-world primates.The Journal of Physiology,258,433–452.
Duong,T.,&Freeman,R.D.(2007).Spatial frequency-specific contrast adaptation originates in the primary visual cortex.Journal of Neurophysiology,98,187–195.
Eger,E.,Schyns,P.G.,&Kleinschmidt.A.(2004).Scale invariant adaptation in fusiform face-responsive regions.Neuroimage,22,232–242.
Elliott,S.L.,Georgeson,M.A.,&Webster,M.A.(2011).Response normalization and blur adaptation:Data and multi-scale model.Journal of Vision,11(2),pii:7.
Engel,S.A.,&Furmanski,C.S.(2001).Selective adaptation to color contrastin human primary visualcortex.The Journal of Neuroscience,21,3949–3954.
Fang,F.,Murray,S.O.,Kersten,D.,& He,S.(2005).Orientation-tuned fMRI adaptation in human visual cortex.Journal of Neurophysiology,94,4188–4195.
Foster,D.H.(2011).Color constancy.Vision Research,51,674–700.
Fox,C.J.,&Barton,J.J.S.(2007).What is adapted in face adaptation?The neural representations of expression in the human visual system.Brain Research,1127,80–89.
Glasser,D.M.,Tsui,J.M.G.,Pack,C.C.,&Tadin,D.(2011).Perceptualand neuralconsequences ofrapid motion adaptation.Proceedings of the National Academy of Sciences of the United States of America,108,E1080–E1088.
Graham,N.V.S.(1989).Visual Pattern Analyzers.Oxford,UK:Oxford University Press.
Grill-Spector,K.,Kushnir,T.,Edelman,S.,Avidan,G.,Itzchak,Y.,&Malach,R.(1999).Differential processing of objects under various viewing conditions in the human lateral occipital complex.Neuron,24,187–203.
Harris,H.,Gliksberg,M.,&Sagi,D.(2012).Generalized perceptual learning in the absence of sensory adaptation.Current Biology,22,1813–1817.
Heinrich,T.S.,&Bach,M.(2001).Contrast adaptation in human retina and cortex.Investigative Ophthalmology&Visual Science,42,2721–2727.
Kohn,A.(2007).Visual adaptation:Physiology,mechanisms,and functional benefits.Journal of Neurophysiology,97,3155–3164.
Kohn,A.,&Movshon,J.A.(2003).Neuronal adaptation to visual motion in area MT of the macaque.Neuron,39,681–691.
Kojima,Y.,Iwamoto,Y.,&Yoshida,K.(2004).Memory of learning facilitates saccadic adaptation in the monkey.The Journal of Neuroscience,24,7531–7539.
Kording,K.P.,Tenenbaum,J.B.,&Shadmehr,R.(2007).The dynamics of memory as a consequence of optimal adaptation to a changing body.Nature Neuroscience,10,779–786.
Kovács,G.,Zimmer,M.,Harza,I.,Antal,A.,&Vidnyánszky,Z. (2005). Position-specificity of facial adaptation.Neuroreport,16,1945–1949.
Krauskopf,J.,Williams,D.R.,&Heeley,D.W.(1982).Cardinal directions of color space.Vision Research,22,1123–1131.
Leopold,D.A.,O’Toole,A.J.,Vetter,T.,&Blanz,V.(2001).Prototype-referenced shape encoding revealed by high-level aftereffects.Nature Neuroscience,4,89–94.
Levinson,E.,&Sekuler,R.(1976).Adaptation alters perceived direction of motion.Vision Research,16,779–781.
Li,G.R.,Ye,X.,Song,T.,Yang,Y.P.,&Zhou,Y.F.(2011).Contrast adaptation in cat lateral geniculate nucleus and influence of corticothalamic feedback.European Journal of Neuroscience,34,622–631.
Maffei.L.,Fiorentini,A.,&Bisti,S.(1973).Neural correlate of perceptual adaptation to gratings.Science,182,1036–1038.
Mather,G.,Pavan,A.,Campana,G.,&Casco,C.(2008).The motion aftereffectreloaded.TrendsinCognitive Science,12,481–487.
McCollough,C.(1965).Color adaptation of edge-detectors in the human visual system.Science,149,1115–1116.
McGovern,D.P.,Roach,N.W.,&Webb,B.S.(2012).Perceptuallearning reconfigures the effectsofvisual adaptation.The Journal of Neuroscience,32,13621–13629.
Meng,X.,Mazzoni,P.,&Qian,N.(2006).Cross-fixation transfer of motion aftereffects with expansion motion.Vision Research,46,3681–3689.
Mesik,J.,Bao,M.,Engel,S.A.(2013).Spontaneous recovery of motion and face aftereffects.Vision Research,89,72–78.
Morgan,M.J.(2012).Motion adaptation does not depend on attention to the adaptor.Vision Research,55,47–51.
Moulden,B.(1980).After-effects and the integration of patterns of neural activity within a channel.Philosophical Transactions of the Royal Society of London Series B:Biological Sciences,290,39–55.
Nathans,J.,Thomas,D.,&Hogness,D.S.(1986).Molecular genetics of human color vision:The genes encoding blue,green,and red pigments.Science,232,193–202.
Neitz,J.,Carroll,J.,Yamauchi,Y.,Neitz,M.,&Williams,D.R.(2002).Color perception is mediated by a plastic neural mechanism that is adjustable in adults.Neuron,35,783–792.
Nishida,S.,Ashida,H.,& Sato,T.(1994).Complete interocular transfer of motion aftereffect with flickering test.Vision Research,34,2707–2716.
Nishida,S.,& Sato,T.(1995).Motion aftereffect with flickering test patterns reveals higher stages of motion processing.Vision Research,35,477–490.
Nishida,S.,Sasaki,Y.,Murakami,I.,Watanabe,T.,&Tootell,R.B.H.(2003).Neuroimaging ofdirection-selective mechanisms for second-order motion.Journalof Neurophysiology,90,3242–3254.
O’Neil,S.F.,&Webster,M.A.(2011).Adaptation and the perception of facial age.Visual Cognition,19,534–550.
Oruç,I.,&Barton,J.J.S.(2011).Adaptation improves discrimination of face identity.Proceedings of the Royal Society B:Biological Sciences,278,2591–2597.
Pavan,A.,Marotti,R.B.,&Campana,G.(2012).The temporal course of recovery from brief(sub-second)adaptations to spatial contrast.Vision Research,62,116–124.
Petersen,S.E.,Baker,J.F.,&Allman,J.M.(1985).Directionspecific adaptation in area MT of the owl monkey.Brain Research,346,146–150.
Phinney,R.E.,Bowd,C.,&Patterson,R.(1997).Directionselective coding of stereoscopic(Cyclopean)motion.Vision Research,37,865–869.
Rieke,F.,&Rudd,M.E.(2009).The challenges natural images pose for visual adaptation.Neuron,64,605–616.
Sanchez-Vives,M.V.,Nowak,L.G.,&McCormick,D.A.(2000).Membrane mechanisms underlying contrast adaptation in cat area 17 in vivo.The Journal of Neuroscience,20,4267–4285.
Schiller,P.H.,& Malpeli,J.G.(1978).Functional specificity of lateral geniculate nucleus laminae of the rhesus monkey.Journal of Neurophysiology,41,788–797.
Seiffert,A.E.,Somers,D.C.,Dale,A.M.,&Tootell,R.B.H.(2003).Functional MRI studies of human visual motion perception:texture,luminance,attention and after-effects.Cerebral Cortex,13,340–349.
Sekuler,R.,&Pantle,A.(1967).A model for after-effects of seen movement.Vision Research,7,427–439.
Sherman,S.M.,Wilson,J.R.,Kaas,J.H.,&Webb,S.V.(1976).X-and Y-cells in the dorsal lateral geniculate nucleus of the owl monkey(Aotus trivirgatus).Science,192,475–477.Smith,M.A.,Ghazizadeh,A.,& Shadmehr,R.(2006).Interacting adaptive processes with different timescales underlie short-term motor learning.PLoS Biology,4(6),e179.
Solomon,S.G.,Peirce,J.W.,Dhruv,N.T.,&Lennie,P.(2004).Profound contrast adaptation early in the visual pathway.Neuron,42,155–162.
Strobach,T.,& Carbon,C.-C.(2013).Face adaptation effects:Reviewing the impact of adapting information,time,and transfer.Frontiers in Psychology,4,318.
Valentine,T.(2001).Face-space models of face recognition.In M.J.Wenger,&J.T.Townsend,(Eds.).Computational,geometric,and process perspectives on facial cognition:Contexts and challenges.Scientific Psychology Series(pp.83–113).Mahwah,NJ:Erlbaum.
van Wezel,R.J.A.,& Britten,K.H.(2002).Motion adaptation in area MT.Journal of Neurophysiology,88,3469–3476.
Vul,E.,Krizay,E.,&MacLeod,D.I.(2008).The McCollough effect reflects permanent and transient adaptation in early visual cortex.Journal of Vision,8(4),4.1–4.12.
Watson,T.L.,&Clifford,C.W.G.(2003).Pulling faces:An investigation of the face-distortion aftereffect.Perception,32,1109–1116.
Webster,M.A.(1996).Human colour perception and its adaptation.Network:Computation in Neural Systems,7,587–634.
Webster,M.A.,Kaping,D.,Mizokami,Y.,&Duhamel,P.(2004).Adaptation to natural facial categories.Nature,428,557–561.
Webster,M.A.(2011).Adaptation and visualcoding.Journal of Vision,11(5),1–23.
Webster,M.A.,& MacLeod,D.I.A.(2011).Visual adaptation and face perception.Philosophical Transactions of the Royal Society B:Biological Sciences,366,1702–1725.Wright,M.J.,&Ikeda,H.(1974).Processing of spatial and temporalinformation in the visualsystem.In F.O.Schmitt&F.G.Worden(Eds.),The neurosciences(Third Study Program)(pp.115–122).Cambridge:MIT Press.
Wuerger,S.(2013).Colour constancy across the life span:Evidence for compensatory mechanisms.PLoS One,8(5),e63921.
Xu,H.,Dayan,P.,Lipkin,R.M.,& Qian,N.(2008).Adaptation across the cortical hierarchy:Low-level curve adaptation affects high-level facial-expression judgments.The Journal of Neuroscience,28,3374–3383.
Yehezkel,O.,Sagi,D.,Sterkin,A.,Belkin,M.,&Polat,U.(2010).Learning to adapt:dynamics of readaptation to geometrical distortions.Vision Research,50,1550–1558.
Zhang,P.,Bao,M.,Kwon,M.,He,S.,&Engel,S.A.(2009).Effects of orientation-specific visual deprivation induced with altered reality.Current Biology,19,1956–1960.
Zhao,L.,&Chubb,C.(2001).The size-tuning of the facedistortion after-effect.Vision Research,41,2979–2994.
猜你喜欢
辽河(2022年4期)2022-06-09
辽河(2022年1期)2022-02-14
辽河(2022年1期)2022-02-14
科学技术创新(2021年9期)2021-04-24
智慧少年·故事叮当(2020年2期)2020-03-08
人与自然(2019年4期)2019-07-26
长江大学学报(自科版)(2019年7期)2019-07-22
读者·校园版(2018年24期)2018-12-04
天然气勘探与开发(2015年2期)2015-12-10
上海制造(2013年11期)2014-03-26
