
昆明56个桂花品种I SSR分析*
2015-01-17 05:42张俊杰马旭廖源王澍林开文
西部林业科学 2015年3期
张俊杰,马旭,廖源,王澍,林开文
(1.西南林业大学园林学院,云南 昆明650224;2.福建农林大学园艺学院,福建 福州350002)
桂花(Osmanthus fragrans)属木犀科(Oleaceae)木犀属(Osmanthus)植物,乃常绿灌木或小乔木,为中国十大传统名花之一,有两千多年的栽培历史。我国是木犀属植物和桂花品种资源的世界分布中心[1]。桂花按照花期和花色,可分为银桂品种群、金桂品种群、丹桂品种群和四季桂品种群,各品种群下再划分品种。与梅花、牡丹等其他传统名花相比,不论在品种分类、品种起源和演化规律的研究都远远不够。据《中国植物志》记载,桂花原产我国西南部,四川、云南、广西等地均有野生。据不完全统计,云南百年以上树龄的桂花有21 株[2]。2009 年翟丽丽[3]调查出昆明市三环内桂花有30个品种。而其他云南省桂花的相关研究很少,没有体现其资源优势。昆明作为云南省省会,属亚热带高原季风气候,夏无酷暑,冬无严寒,素以“春城”而享誉中外,独特的环境气候造就昆明独有的桂花品种。因此,发掘并研究昆明潜在的桂花品种对我国桂花种质资源的拓展尤为重要。
近年来越来越多的学者运用分子生物学技术研究桂花的遗传多样性和亲缘关系,如同工酶分析、RAPD标记、ISSR-PCR标记、AFLP分析等。ISSR(inter-simple sequence repeat)即简单重复间序列标记,具有操作简单、DNA用量少、重复性高、成本低等特点[4],在桂花品种鉴定、分子标记辅助选择和品种的进化研究方面有广泛的应用前景[5~6]。本研究利用ISSR分子标记技术,对昆明市区的56个桂花品种 (包括10个待定品种)和4个品种群间的遗传多样性和亲缘关系进行分析,以期为昆明桂花品种资源鉴定和选育提供理论依据,从而培育出更多观赏价值高、抗逆性强的名优桂花新品种。
1 材料和方法
1.1 材料
本研究实验材料选用2012年7月至2013年10月在昆明市五城区各类型绿地调查到的56个桂花品种,木犀属的2个对照种华东木犀 (Osmanthus cooperi,采集于昆明世博园)和云南木犀(O.yunnanensis,采集于西南林业大学)共58份材料。包含了桂花的4个品种群,其中四季桂品种群品种5个,银桂品种群品种18个,金桂品种群品种16个,丹桂品种群品种7个,根据形态和花色暂命名的待定品种10个 (表1)。将资源调查中采集的无病虫害的幼嫩叶片放入自封袋中,立即加入硅胶干燥,于-70℃冰箱保存备用。

表1 58个木犀属植物试验材料Tab.1 58 test material of Osmanthus genus plants
续表1
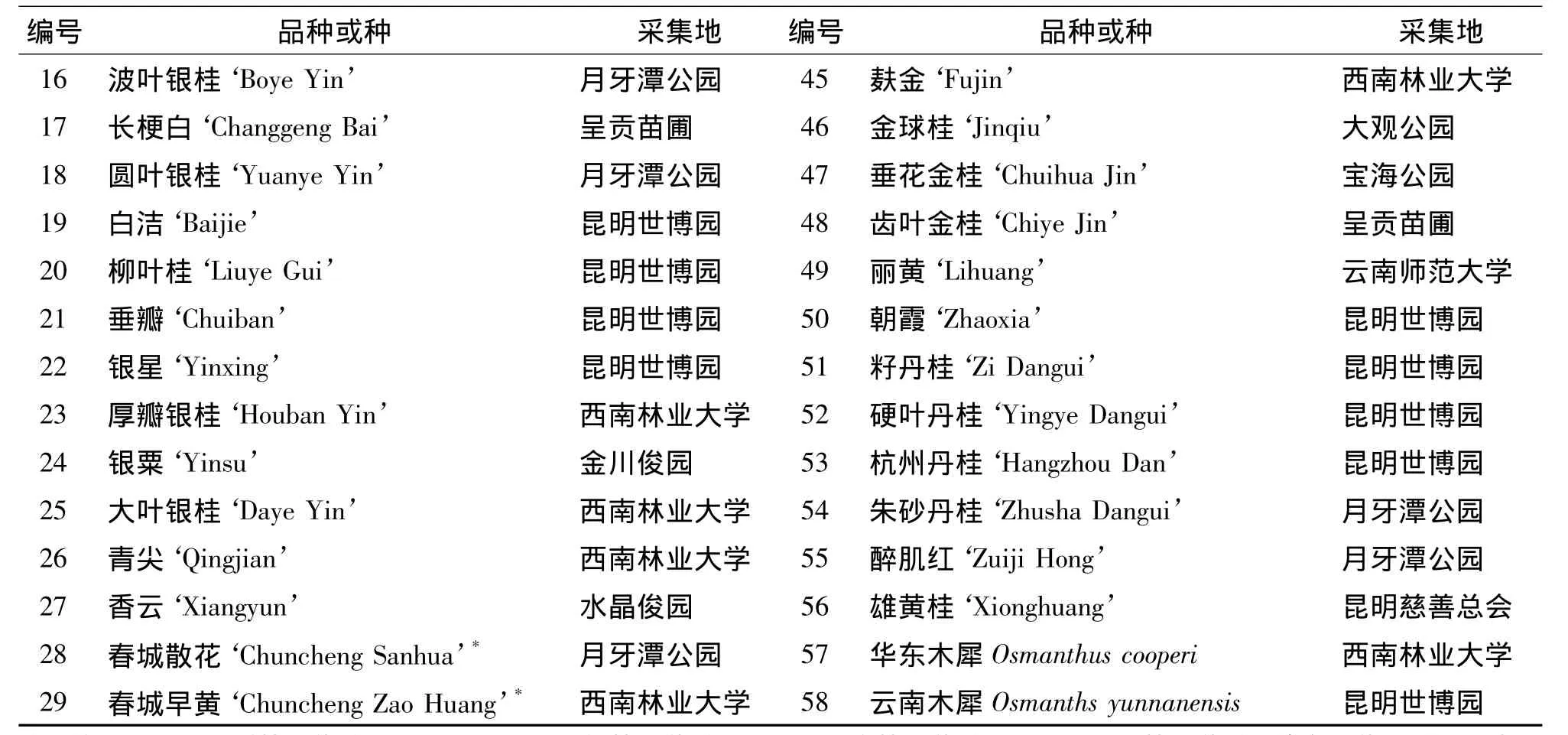
注:编号1~5属四季桂品种群;6~11,16~27属银桂品种群;34~49属金桂品种群;50~56属丹桂品种群;待定品种用“*”表示,12~15为结实待定品种,28~33为不结实待定品种。
1.2 方法
1.2.1 DNA 提取
采用OMEGA HP Plant DNA Kit试剂盒提取样品基因组DNA,核酸蛋白测定仪检测DNA的纯度,用1.0%的琼脂糖在1×TAE液里电泳检测纯度,存放入-20℃冰箱中备用。
1.2.2 引物筛选
引物选用美国哥伦比亚大学公布的100条ISSR引物序列与相关文献里的两条引物[6,8~9]ISSR12、ISSR27共102条引物,由生工生物工程 (上海)股份有限公司合成,从中筛选出扩增条带清晰、丰富、弥散较少的24条引物。
1.2.3 ISSR-PCR 扩增
本实验采用 Takara Mix代替 dNTP、Mg2+、Taq DNA聚合酶与buffer。ISSR-PCR扩增反应采用25 μL 体系,含 Mix 12.5 μL,10 mol/L 的引物 1 μL,50 ng桂花样品DNA 1 μL,ddH2O补充至25 μL。扩增程序为:94℃预变性5 min,94℃变性1 min,50~54℃复性 1 min,72℃延伸 1.5 min,40个循环,72℃延伸10 min。对58个样品进行正式扩增,每个引物扩增两次。PCR产物在以Goldview I为燃料的2.0%的琼脂糖凝胶电泳,用5 μL PCR产物上样,恒压100 v,电泳时间45 min,取出凝胶在凝胶成像系统下观察并拍照。
1.3 数据处理
依据分子标记的有无记录成二元数据,相同位点上有带的位置记为1,无带或带不清晰的位置记为0,录入Excel表格,导入Pop Gene Version 1.32软件进行遗传分析。用NTSYSpc2.1软件对实验数据用UPGMA法聚类分析,绘出58份材料的聚类分析树状图。
2 结果与分析
2.1 ISSR引物扩增多态性
将筛选出的24个引物对供试材料进行ISSR-PCR扩增反应 (表2),发现扩增片段的大小主要集中在300~1 800 bp之间,共扩增出251个位点,多态性位点数达243个,多态性位点比率为96.81%。每个引物扩增出的位点在7至14个之间。平均每个引物扩增位点10.46个,多态性位点10.13个。多态性比率达到 100%的引物有UBC810、UBC811等共17个。引物 UBC841的多态性位点比率最低,为77.78%。引物UBC823、ISSR12的扩增图谱 (图1)。

表2 24条ISSR引物序列及对供试桂花品种的扩增结果Tab.2 24 ISSR primers and amplification results of tested Osmanthus fragrans Cultivars
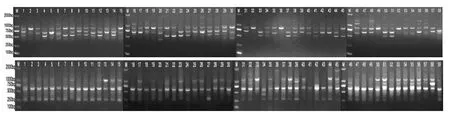
图1 引物UBC823、ISSR12对供试材料的扩增结果Fi g.1 Ampl i f i cat i on r esul t s of t est ed mat er i al s wi t h pr i mer UBC823 and ISSR12
2.2 ISSR遗传距离和聚类分析
58份材料的聚类分析树状图见图2,以遗传相似系数0.726为截值,可将材料分为4类。第Ⅰ类为2个四季桂品种:长梗素花、日香桂。第Ⅱ类包括了四季桂、金桂、银桂和丹桂4个品种群的54个品种,占供试桂花品种的96.43%。第Ⅲ类和第Ⅳ类分别为对照种华东木犀、云南木犀。此时桂花56个品种已与木犀属2个对照种区分开。聚类结果提炼到摘要中去。
以遗传相似系数0.760为截值,可把第Ⅱ类的54个桂花品种分为7个组。第1组为四季桂、银桂、金桂3个品种群共38个品种组成的混合类型。含大叶佛顶珠等3个四季桂品种,波叶银桂等13个银桂品种,潢川金桂等12个金桂品种和10个待定品种。第2组和第3组均含2个不结实的银桂品种:长梗白和银粟,圆叶银桂和白洁。第4组含丽黄等3个金桂品种和醉肌红等4个丹桂品种。第5组为金桂品种金球桂。第6组含朝霞等3个丹桂品种。第7组为银桂品种大叶银桂。
以遗传相似系数0.790为截值,第Ⅱ类第1组中天香台阁等3个四季桂品种与银桂品种长叶碧珠,待定品种厚瓣籽银桂聚类;2个银桂品种垂瓣、银星聚类;待定品种昆明多瓣籽银桂单独聚类;2个待定品种春城早黄和春城散花优先聚类,再与潢川金桂等3个金桂品种聚类;银桂品种早籽银与金桂品种小叶籽金桂聚类;金桂品种大叶黄,2个银桂品种波叶银桂、米花籽银桂和3个待定品种长梗白籽银桂、迷你银桂、柳叶籽银桂聚类;银桂品种籽银桂,2个待定品种小银铃、三瓣早黄和晚金桂等4个金桂品种聚类;镰叶金桂与香云等3个银桂品种聚类;金桂品种垂枝黄与2个银桂品种柳叶桂、青尖聚类;银桂品种厚瓣银桂、待定品种春城齿叶银桂和金桂品种麸金聚类。第4组2个金桂品种球桂、波叶球桂和丹桂品种醉肌红聚类;杭州丹桂等3个丹桂品种聚类。
10个待定品种花色较浅,根据其形态特征和聚类分析结果,可推测均为银桂品种群的品种。据此,以下研究均把10个待定品种归属于银桂品种群。

图2 供试材料的ISSR聚类分析树状图Fig.2 ISSR dendrogram of cluster analysis for 56 cultivars and 2 contrast of Osmanthus
2.3 ISSR遗传多样性
遗传多样性研究对充分认识桂花品种,有效保护桂花品种资源,全面、合理地开发云南的桂花品种资源具有重要的理论和应用意义。根据电泳图谱条带分析得出桂花不同品种群的遗传多样性指数(表3)。在4个品种群中,银桂品种群的观察等位基因数 (1.828 7)、有效等位基因数 (1.301 6)、Nei’s遗传多样性指数 (0.188 5)、Shannon 多样性指数 (0.300 9)和位点多态性比率 (82.87%)皆最高。各品种群的观察等位基因数、有效等位基因数、Nei’s遗传多样性指数、Shannon多样性指数和多态性位点比率排列顺序为,银桂品种群>金桂品种群>四季桂品种群>丹桂品种群。
4个桂花品种群群体总基因多样性指数为0.247 4,种群内遗传多样性指数为0.112 6,种群间遗传多样性比率为54.49%,种群内遗传多样性比率为45.51%,表明桂花品种群之间的变异大于品种群之内的变异。

表3 4个品种群ISSR标记的遗传多样性指数Tab.3 ISSR maker-based Genetic variation index of 4 Osmanthus fragrans groups
2.4 ISSR遗传距离和遗传相似系数
根据电泳图谱条带分析桂花4个桂花品种群间和2个木犀属对照种的ISSR遗传相似系数和遗传距离 (表4)。遗传距离在0.014 0~0.475 6之间。遗传距离最大的在2个对照种华东木犀与云南木犀之间 (0.475 6),2个对照种与桂花4个品种群之间的遗传距离均≥0.217 0。而在桂花4个品种群之间,银桂、金桂、四季桂品种群与丹桂品种群的遗传距离较远,而四季桂品种群与丹桂品种群的遗传距离最远 (0.093 9);银桂品种群与金桂品种群之间的遗传距离最近 (0.014 0)。

表4 4个品种群和2个木犀属对照种的ISSR遗传相似系数和遗传距离矩阵Tab.4 Genetic similar coefficient and genetic distance matrix of 4 Osmanthus fragrans groups and 2 controls
根据ISSR遗传相似系数生成桂花4个品种群和2个对照种的ISSR聚类图见图3,银桂品种群与金桂品种群先聚在一块,再与四季桂品种群聚在一起,花色较浅的3个品种群聚在一块后,再与丹桂品种群聚在一块。2个对照种华东木犀、云南木犀与桂花4个品种群之间亲缘关系较远,与各桂花品种群明显区分开。

图3 基于Nei’s遗传相似系数的桂花4个品种群及2个对照种的ISSR聚类图Fig.3 ISSR clustering dendrogram based on Nei’s genetic similar coefficient of 4 Osmanthus fragrans groups and 2 controls
3 结论与讨论
3.1 结论
本研究中天香台阁等3个四季桂品种与厚瓣籽银桂、长梗素花2个银桂品种聚类,证明了银桂品种与四季桂品种亲缘关系较近。花色浅黄白的银桂品种与花色浅黄的金桂品种聚类,花色金黄的金桂品种与花色最浅的丹桂品种醉肌红聚类,表明同一品种群内的品种,各品种群之间花色相近的品种大多聚类。本研究中大叶银桂、金球桂单独聚类,表明同一品种群内某些花色相近的品种聚类分散,亲缘关系较远,它们的遗传变异程度和遗传分化程度较高。从10个待定品种的聚类方式可知,待定品种均为银桂品种。因此调查出的昆明银桂品种多达28个,占供试桂花品种的50%,说明银桂品种在昆明的演化变异较多,形态和基因多样。
实验结果表明桂花品种之间遗传多样性丰富,桂花品种群之间的变异大于品种群之内的变异,各品种群多态性位点比率依次为:银桂品种群>金桂品种群>四季桂品种群>丹桂品种群。通过4个品种群的ISSR聚类图分析,银桂品种群与金桂品种群优先聚类,再与四季桂品种群聚类,最后与丹桂品种群聚类。
3.2 讨论
银桂品种与四季桂品种亲缘关系较近的观点,与用同工酶、AFLP 技术和胡绍庆的 ISSR-PCR[5,10~11]分析得出的观点一致。花色相近的品种聚类,与上海、湖北、杭州、江苏等地区[5~8,11~13]的研究结论一致。说明花色作为桂花品种形态学上重要的分类依据,得到许多分子水平研究的验证。至于同一品种群内某些花色相近的品种聚类分散,这可能由于跟花色有关的基因在桂花的基因组中所占分量不大,不同桂花品种间相互杂交选育导致其种质资源混杂,以花色为分类标准的传统的形态分类没能确切反映桂花品种间的亲缘关系[12,14]。另外,春城早黄与三瓣早黄亲缘关系较远,而其形态上相似,这可能由于植物体的内部构造和形态建成复杂,DNA水平上的相似在生理或形态上可能差距较远[15],这些具体原因有待进一步研究。
桂花各品种群多态性位点比率与李梅、梁丽丽的研究成果[6,8]不完全相同。可能是由于云南特殊地理环境和长期栽培选育造成与其他地区的品种群多态性位点比率不一致。
4个品种群的ISSR聚类方式不同于李梅[6]的秋桂类3个品种群优先聚类,再与四季桂品种群聚类。从花色角度来看,金桂品种群、银桂品种群先和花色较浅的四季桂品种群聚类有一定形态学支撑,也以基因角度印证了藏德奎比较形态学研究[16~17]提出的秋桂类3个品种群是从银桂进化到金桂再进一步颜色加深进化到丹桂的推论。
10个待定品种若再经一段时间的观察,性状稳定后可扩繁并推广到园林中,成为昆明特有的品种品牌。厚瓣籽银桂和昆明多瓣籽银桂为新发现的结实品种,可与亲缘关系较近的国内常见品种长叶碧珠进行杂交授粉,获得的新品种有望继承长叶碧珠的生态习性,可推广到平原地区栽植。待定品种春城早黄、春城散花与潢川金桂、小花金桂、垂花金桂等3个金桂品种聚类,这5个品种均花量大、花期较长、花色较艳,在昆明生长良好,由于亲缘关系较近导致优良品性类似,值得大量繁殖推广。
[1]李梅.桂花种质资源遗传多样性研究及品种鉴定[D].南京:南京农业大学,2009.
[2]孙卫邦,向其柏.云南古桂的资源现状及保护利用[J].南京林业大学学报(自然科学版),2004,28(增刊):30-34.
[3]翟丽丽.昆明市桂花品种资源调查及园林应用研究[D].昆明:西南林业大学,2009.
[4]Moreno S.Martin J.P,Ortiz J.M.ISSR-PCR for characterization of closely related grape vine germplasm[J].Euphytica,1998,101:117-125.
[5]胡绍庆,邱英雄,吴光洪,等.桂花品种的ISSR-PCR分析[J].南京林业大学学报(自然科学版),2004,28(增刊):71-75.
[6]李梅,侯喜林,单晓政,等.部分桂花品种亲缘关系及特有标记的 ISSR分析[J].西北植物学报,2009,29(4):674-682.
[7]邱英雄,胡绍庆,吴光洪,等.ISSR-PCR技术在桂花品种分类研究中的应用[J].园艺学报,2004,31(4):529-532.
[8]梁丽丽.湖北省及江苏南部桂花品种资源调查及部分品种ISSR分析[D].南京:南京林业大学,2011.
[9]段一凡,王贤荣,梁丽丽,等.古桂遗传多样性与亲缘关系的ISSR分析[J].福建林学院学报,2011,31(4):335-339.
[10]陈建业,宁玉霞,赵翠花,等.河南桂花品种过氧化物酶同工酶研究[J].园艺学报,1995,22(2):176-180.
[11]韩远记,董美芳,袁王俊,等.部分桂花栽培品种的AFLP 分析[J].园艺学报,2008,35(1):137-142.
[12]韩远记,董美芳,袁王俊,等.桂花品种资源的遗传多样性分析[J].植物学通报,2008,25(5):559-564.
[13]赵小兰,姚崇怀.桂花部分品种的RAPD分析[J].华中农业大学学报,1999,18(5):484-487.
[14]尚富德,韩远记,袁王俊,等.木犀属及桂花品种分类研究进展[J].河南大学学报(自然科学版),2012,42(5):608-612.
[15]李书粉.基于ISSR和ITS的木犀属植物亲缘关系及系统分类研究[D].新乡:河南师范大学,2007.
[16]藏德奎,向其柏,刘玉莲.木犀属品种分类研究[J].林业科学,2006,42(5):17-21.
[17]藏德奎,向其柏,刘玉莲.中国桂花品种的起源与演化[C].中国桂花,申报品种国际登录专集(Ⅱ).2004:1-12.
猜你喜欢
莫愁·小作家(2022年4期)2022-04-19
作文小学中年级(2020年9期)2020-09-29
红楼梦学刊(2020年3期)2020-02-06
财讯(2019年33期)2019-08-04
天然产物研究与开发(2018年2期)2018-04-04
西江月(2017年4期)2017-11-22
作文周刊·高二版(2016年32期)2017-04-13
小雪花·成长指南(2016年10期)2016-11-01
中国科技信息(2015年4期)2015-01-02
中医研究(2013年5期)2013-03-11
