
基于线粒体ATP6基因序列的作物系统发育研究
2015-01-10 02:11胡晓天刘齐元范彦君
湖南农业科学 2015年10期
胡晓天,向 妍,张 柯,刘齐元,范彦君,周 玮
(1. 湖南农业大学植物保护学院,植物病虫害生物学与防控湖南省重点实验室,湖南 长沙 410128; 2. 湖南农业大学,湖南省生物农药与制剂加工工程技术研究中心,湖南 长沙 410128;3. 江西农业大学农学院,作物生理生态与遗传育种教育部重点实验室,江西 南昌 330045)
基于线粒体ATP6基因序列的作物系统发育研究
胡晓天1,2,向 妍1,2,张 柯1,2,刘齐元3,范彦君1,2,周 玮1,2
(1. 湖南农业大学植物保护学院,植物病虫害生物学与防控湖南省重点实验室,湖南 长沙 410128; 2. 湖南农业大学,湖南省生物农药与制剂加工工程技术研究中心,湖南 长沙 410128;3. 江西农业大学农学院,作物生理生态与遗传育种教育部重点实验室,江西 南昌 330045)
ATP6编码ATP合酶a亚基,对酶正常功能的实现作用重要。为探讨线粒体ATP6基因在作物系统发育分析上的应用,采用NJ法的Kimura 2-parameter模型、E距离模型,以及最大似然法对30种作物进行分子系统树构建,并结合植物形态学分类结果进行比较分析。结果表明,基于NJ法和最大似然法的ATP6系统发育树聚类结果一致,契合现有植物形态学分类,在分析作物物种分化起源上有巨大潜力。
ATP合酶;ATP6基因;作物;系统发育
线粒体ATP合酶也叫F1Fo-ATP合酶或复合体Ⅴ,是一个位于线粒体内膜的旋转酶,与呼吸链复合体Ⅰ-Ⅳ在一起[1]。线粒体ATP合酶由两个功能结构域组成:F1处于线粒体的基质中,Fo结合在线粒体内膜上[2]。F1是是水溶性的,突出于线粒体内膜内侧,由3个α和β亚基,以及1个γ、δ和ε亚基构成[2-3]。Fo包含一个c环亚基,a、b、d、F6亚基和一个寡霉素敏感相关蛋白(the oligomycinsensitivity-conferring protein,OSCP)。随着质子流经Fo部分的a亚基进入c环亚基,质子浓度梯度驱动两个分子发动机转动:c环亚基绕着与之相连的中央柱状结构转动,其又带动被α3β3六聚体包着一端的γ亚基转动,为ATP合成提供能量[2]。在整个催化过程中,由于外围柱状结构的作用,α3β3六聚体相对于a亚基总是保持固定。因此ATP合酶可以被机械地划分为“转动子”(c环、γ、δ、ε亚基)和“固定子”(α3β3、a、b、d、F6、OSCP亚基)[2]。
ATP合酶并不是以“孤岛”形式结合在线粒体内膜上的,而是有规律地以二聚体或更高形式的寡聚体组织在一起。a亚基由于可能存在大量的跨膜螺旋而被认为对ATP合酶的二聚作用有着重要的影响[4]。两个Fo部分的亚基:a亚基和A6L亚基分别由线粒体ATP6(mitochondrialDNA encoded ATP6,MT-ATP6)和ATP8(mitochondrial DNA encoded ATP8,MT-ATP8)编码。在人体中,缺失了线粒体DNA(即a亚基和A6L亚基不能形成),余下的ATP合酶亚基会装配成一个550 kD的复合体,略小于完整的ATP合酶(597kD)[4]。MT-ATP6和MT-ATP8基因的突变会引起ATP合酶的变异,MT-ATP6突变是最早报导[5]且目前报导最频繁的ATP合酶基因缺陷;在人体中MTATP6的点突变会导致人的一系列临床疾病[6]。a亚基尚有另两个功能:引导质子从线粒体内膜进入线粒体基质中;另一功能致使在膜内接触a亚基的c环亚基转动,由此产生的机械能诱导F1外膜结构域催化位点的构象变化,帮助ATP合成[2]。
在基于序列分析的真核生物分类上,由于26SrDNA含有能区分相同和不同物种的保守区和可变区,被广泛地应用于酵母分类[7]。也有将其应用于植物分类并讨论选取序列中的不同区段如保守核心、扩展片段或整个序列等对构树结果的影响[8]。MT-ATP6所编码的蛋白质产物在ATP合酶中起着重要的作用[2,4,9-10],其序列中某些位点的突变可能会引发一系列生物学问题[6,11-12],但也可能用于物种分类分析。文中拟以不同方法模型对多种作物ATP6序列进行系统发育树构建,以探讨ATP6序列在作物分类上的应用。
1 材料与方法
1.1 实验数据
从美国国立生物技术信息中心(NCBI)同时下载了表1中30种作物线粒体的ATP6序列。在表1作物的分类上,使用经典的以植物形态学特征为依据的克朗奎斯特分类法[13]。
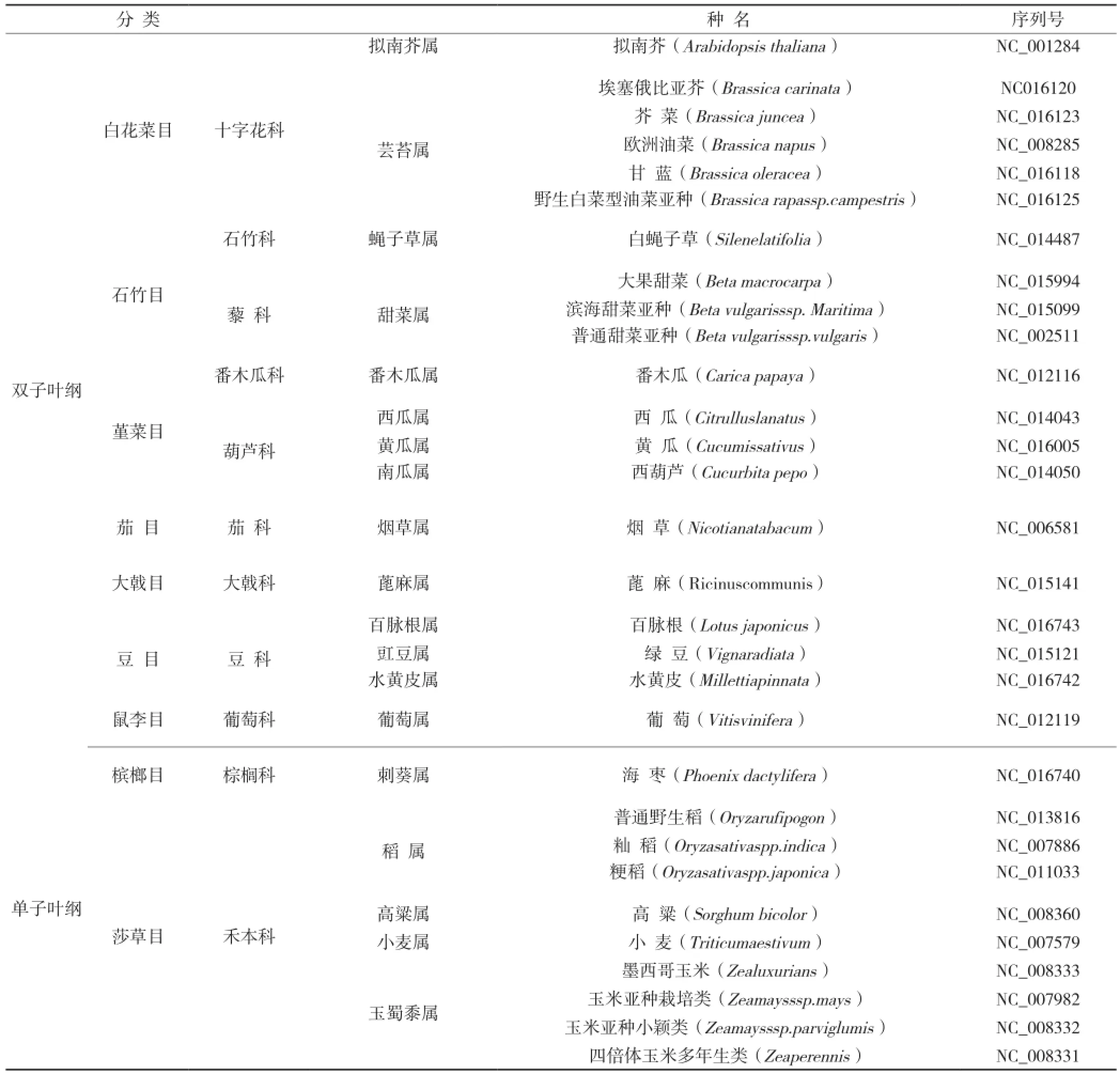
表1 作物线粒体基因组
1.2 系统发育树的构建
将下载得到的数据分别用ClustalX软件进行匹配对齐,用MEGA6.06软件中的邻接(Neighbor-joining,NJ)距离法进行构树[14]。NJ法有着许多模型参数选项,常用的Kimura 2-parameter模型对碱基的转换和颠换率差异做了校正,但是假定了不同位点替换速率相等以及核苷酸频率相等,这或许会导致一定的偏差。由π电子共振能来计算物种的系统发育距离在一定程度上考虑了核苷酸的替换频率和速率;这种距离被定义为E距离,E=1-2c/(a+b),(a、b分别为被替换序列之间的π电子共振能之和,c为按位比较被替换序列之间相同碱基的π电子共振能之和,各碱基π电子共振能比值为A∶G∶C∶T= 0.32∶0.27∶0.23∶0.17),它对应群落间按碱基能量计算的群落相异性指数[15]。通过使用NJ法中的Kimura 2-parameter模型对表1中植物的ATP相关基因进行系统发育树构建,并进行1 000次Bootstraps检验;同时亦使用E距离模型和最大似然(Maximum Likelihood,ML)法建树作为对比参照。
2 结果与分析
用NJ法分别构建表1中30种作物线粒体基因ATP6的Kimura 2-parameter模型系统发育树(图1)、E距离模型系统发育树(图2)及最大似然法系统发育树(图3)。由图1系统树可以看出:与经典形态学分类结果一致,差别最明显的单子叶作物和双子叶作物被分为最大的两类;所有相同目的作物被归为一个分支,其中所有相同科的作物聚在一起后再与同目不同科的作物聚在一起,所有的6种十字花科物、3种藜科作物、3种葫芦科作物、3种豆科作物、9种禾本科作物被聚为一个单系;但是堇菜目的番木瓜未和在其他堇菜目植物(葫芦科)聚在一起,而是和鼠李目的葡萄聚在一起。图2中ATP6的E距离系统发育树与Kimura 2-parameter系统树类似,都有着与经典形态学分类一致的较好的分类结果;在对堇菜目番木瓜和堇菜目葫芦科植物的归类上,同样支持将两者分为不同类。以NJ法的Kimura 2-parameter模型以及最大似然法对ATP6作图,都显示堇菜目番木瓜和堇菜目葫芦科植物为不同类目,而目前较新的APGII[16]分类法已将番木瓜由堇菜目改为十字花目,与之前葫芦科植物所属堇菜目(现改为葫芦目葫芦科)区分开。
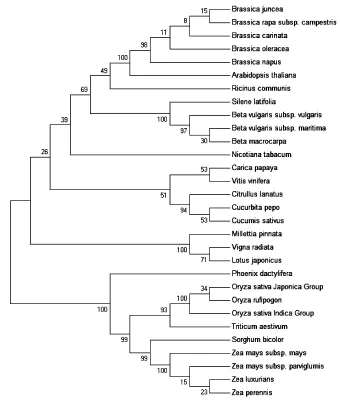
图1 Kimura 2-parameter 模型构建的ATP6系统发育树

图2 E距离模型构建的ATP6系统发育树

图3 最大似然法构建的ATP6系统发育树
3 讨 论
由此看来,具有重要生物功能的ATP6能够将既定作物正确分类,而且聚类结果较于传统的形态学分类更符合较新的植物分类规则。线粒体ATP6编码ATP合酶的a亚基,对ATP合酶生物学功能的正常运转起着重大的作用,有着巨大的生物学分析潜力。现已有学者运用MT-ATP6来对大西洋鳗鱼进行物种形成分析[17]。
一直以来,对于植物物种的分类在不断地发生改变,越来越多的科学家从一开始使用克朗奎斯特形态学分类法到现在逐渐倾向于较新的APGⅡ分类法,而APG也从第一版改编到第三版。基于线粒体ATP6基因的系统发育分析结果符合现有植物学分类,或能给以后作物的分类乃至系统发育和物种起源等分析提供参考。
[1] Wittig I,Schägger H. Structural organization of mitochondrial ATP synthase[J]. Biochimica et Biophysica Acta,2008,(4):592-598.
[2] Jonckheere A I,Smeitink J A M,Rodenburg R J T. Mitochondrial ATP synthase: architecture,function and pathology[J]. Journal of Inherited Metabolic Disease,2012,35(2):211-236.
[3] 张吉斌,刘月平,方美英. 线粒体 ATP 合酶基因组成及生化机制研究进展[J]. 中国畜牧杂志,2010,(7): 64-68.
[4] Wittig I,Meyer B,Heide H,et al. Assembly and oligomerization of human ATP synthase lacking mitochondrial subunits a and A6L[J]. Biochimica et Biophysica Acta,2010,1797(7):1004-1105.
[5] HoltI J,Harding AE,Petty RK,et al. A new mitochondrial disease associated with mitochondrial DNA heteroplasmy[J]. American Journal of Human Genetics,1990,46(3):428-433.
[6] Sgarbi G,Baracca A,Lenaz G,et al. Inefficient coupling between proton transport and ATP synthesis may be the pathogenic mechanism for NARP and Leigh syndrome resulting from the T8993G mutation in mtDNA[J]. Biochemical Journal,2006,395(3):493-500.
[7] Kurtzman C P,Robnett C J. Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S)ribosomal DNA partial sequences[J]. Antonie Van Leeuwenhoek,1998,73(4):331-371.
[8] Kuzoff R K,Sweere J A,Soltis D E,et al. The phylogenetic potential of entire 26S rDNA sequences in plants[J]. Molecular Biology & Evolution, 1998, 15(3):251-263.
[9] Manfredi G,Fu J,Ojaimi J,et al. Rescue of a deficiency in ATP synthesis by transfer of MTATP6,a mitochondrial DNA-encoded gene to the nucleus[J]. Nature Genetics,2002,30(4):394-399.
[10] 魏 磊,费震江,丁 毅. 紫稻线粒体ATP合成酶atp6基因转录本RNA编辑[J]. 植物科学学报,2010,28(3):251-256.
[11] Thyagarajan D,Shanske S,Vazquez-Memije M,et al. A novel mitochondrial ATPase 6 point mutation in familial bilateral striatal necrosis[J]. Annals of Neurology,1995,38(3):468-472.
[12] Akagi H,Sakamoto M,Shinjyo C,et al. A unique sequence located downstream from the rice mitochondrial atp6 may cause male sterility[J]. Current Genetics,1994,25(1):52-8.
[13] Cronquist A. An integrated system of classification of flowering plants[M]. New York:Columbia University Press,1981.
[14] 吕宝忠. 分子进化树的构建[J]. 动物学研究,1993,14(2):186-193.
[15] 袁哲明,欧阳芳,吉洪湖,等. 基于碱基分子质量和于碱电子共振能的几种新距离在系统发育重建中的应用[J]. 生命科学研究,2005,9(1): 77-83.
[16] Bremer B,Bremer K,Chase MW,et al. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II[J]. Botanical Journal of the Linnean Society,2003,141(4):399-436.
[17] Jacobsen M W,Pujolar J M,Gilbert M T,et al. Speciation and demographic history of Atlantic eels (Anguilla anguilla and A. rostrata)revealed by mitogenome sequencing[J]. Heredity,2014,113:432-442.
(责任编辑:高国赋)
Phylogenetic Analysis of Crops with Mitochondrial ATP6 Gene Sequence
HU Xiao-tian1,2,XIANG Yan1,2,ZHANG Ke1,2,LIU Qi-yuan3,FAN Yan-jun1,2,ZHOU Wei1,2
(1. Hunan Key Laboratory for Biology and Control of Plant Diseases and Insect Pests, College of Plant Protection, Hunan AgriculturalUniversity,Changsha 410128,PRC; 2. Hunan Engineering & Technology Research Center for Bio-Agrochemical and Pharmaceutic Processing, Hunan Agricultural University, Changsha 410128, PRC; 3. Ministry of Education's Key Laboratory of Crop Physiology, Ecology and Genetic Breeding, College of Agronomy, Jiangxi Agricultural University, Nanchang 330045, PRC)
ATP6 encodes the subunit a of ATP synthase and plays an important role in the normal function of enzyme. The neighborjoining Kimura 2-parameter method, neighbor-joining E method and maximum likelihood method were used to explore the phylogenetic relationships of 30 various crops base on mitochondrial gene ATP6 in this study. The results showed that the phylogenetic relationships base on mitochondrial ATP6 among species of the given crops were consistent with present morphological classifcation. The mitochondrial genes ATP6 have a great potential in speciation history of crops.
crop; ATP synthase; ATP6 gene; phylogeny
Q949
:A
:1006-060X(2015)10-0001-04
10.16498/j.cnki.hnnykx.2015.10.001
2015-08-17
国家自然科学基金(31301388);国家自然科学基金(31260350);中国博士后特别资助项目(2015T80870);中国博士后面上项目(2014M562109);湖南省自然科学基金(14JJ3092);湖南省科学技术厅科技计划项目(2014GK3046);江西省教育厅科技计划项目(GJJ13275);湖南农业大学大学生创新性实验计划项目(XCX14071)
胡晓天(1993-),男,浙江平湖市人,本科生,主要从事生物信息学研究。
周 玮
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
海洋通报(2021年1期)2021-07-23
生物学通报(2021年4期)2021-03-16
世界最新医学信息文摘(2020年68期)2020-12-25
延安大学学报(医学科学版)(2019年1期)2019-03-29
天然产物研究与开发(2018年10期)2018-11-06
癌变·畸变·突变(2014年1期)2014-03-01
现代检验医学杂志(2014年3期)2014-02-02
中国应用生理学杂志(2013年2期)2013-03-25
中国医学科学院学报(2013年6期)2013-03-11
