
山药连作对土壤微生物群落及土壤酶活性的影响
2015-01-03 06:30孙凯宁王克安吕晓惠柴秀乾
水土保持研究 2015年6期
孙凯宁,杨 宁,王克安,吕晓惠,柴秀乾
(山东省农业科学院 蔬菜花卉研究所/国家蔬菜改良中心山东分中心/山东省设施蔬菜生物学重点实验室,济南250100)
山药属薯蓣科,为多年蔓生草本植物薯蓣(Dioscoreaopposita)的根茎,营养价值高,具有免疫调节、抗氧化、降血脂、降血糖、抗肿瘤以及调节脾胃等功效[1]。但山药若在同一地块连续种植,易造成病虫害多发且减产严重,表现出明显的连作障碍[2]。土壤微生物是土壤的重要组成部分,它几乎参与了土壤中的所有生化反应,其主要生理菌群则直接参与到土壤碳、氮等营养元素的循环和能量流动中,其数量和活性直接影响到土壤生态系统的质量和维持[3-4]。同时,土壤微生物可作为植物病原菌或生长促进因子直接影响到作物生长,也可通过与其他微生物的相互作用间接对作物起到促进和抑制作用[5-6]。土壤酶活性可以反映土壤中进行的各种生物化学反应过程的动向和强度,尽管数量较少,但作用颇大[7],因为它们能够参与各种元素的生物循环、有机质的转化、腐殖质和有机无机胶体的形成等。土壤酶活性可作为衡量土壤生物学活性和土壤生产力的指标[8]。
我国蔬菜种植面积广大,但由于经济利益的驱使,长期以来人们过度追求产量,大量施用农药和化肥,且复种指数高,连作现象普遍,使土壤环境日益恶化。目前,针对蔬菜连作土壤中养分变化、微生物及酶类变动情况已有较多报道[9-10],但关于山药不同连作年限及轮作之后土壤微生物及酶活性的变化特点却较为鲜见。本文以此为切入点,研究不同连作年限和轮作(山药—小麦)模式条件下山药种植土壤耕层主要微生物数量和土壤酶活性的变化特征,以期为更加深入探讨山药连作障碍机理与控制技术提供有效支撑。
1 材料与方法
1.1 试验设计
试验于2014年11月21日在山东省桓台县新城细毛山药农民专业合作社的试验地进行取样,分别对山药连作(0a:CK;1a:T1;2a:T2)和轮作(山药—小麦:T3)土壤耕层(0—30cm)进行多点取样,每类样地按“S形”选取5个样点,每个样点按随机多点混合取样法将土样混合均匀。混匀的土样一部分风干保存,测定主要土壤酶活性,一部分在4℃下保存鲜样,测定主要微生物数量。
1.2 测定项目及方法
1.2.1 土壤微生物数量的测定 细菌、真菌、放线菌计数采用稀释涂抹平板法。细菌培养采用葡萄糖牛肉膏蛋白胨培养基,真菌采用马丁孟加拉红培养基,放线菌采用淀粉硝酸钾培养基(高氏一号)。微生物数量以每克样品的菌数表示。每克样品的菌数=同一个稀释度几次重复的菌落平均数×5×稀释倍数。
1.2.2 土壤酶活性测定 不同酶活性参照严昶升[11]的方法。脲酶采用苯酚—次氯酸钠比色法;过氧化氢酶采用高锰酸钾滴定法;蔗糖酶活性用3,5-二硝基水杨酸比色法测定,磷酸酶用磷酸苯二钠(4-氨基安替比林)比色法。
1.3 数据分析
原始数据的整理采用Excel软件完成;差异显著性测验采用SPSS软件完成。
2 结果与分析
2.1 山药连作和轮作模式对土壤主要理化性状的影响
由表1可知,山药连作与轮作对耕层土壤的pH影响不大,各处理间无差异;EC值随着连作年限的增加呈上升趋势,T1,T2分别比CK增加了14.4%和30.7%,T3较T2降低了8.17%,说明连作增加了土壤含盐量,而轮作可以起到降低土壤含盐量的效果。就对土壤肥力的影响而言,各指标均表现出了相似的规律,即随着连作年限的增加,各肥力指标逐渐降低,而轮作之后的土壤耕层肥力则有所增加。T2的有机质、全氮、有效磷和速效钾含量分别比T1降低了18.5%,5.88%,18.2%和8.79%,而T3则分别比T2增加了75.6%,37.5%,54.4%和11.8%,说明轮作对恢复土壤肥力有一定效果。

表1 山药不同种植模式土壤(0-30cm)主要理化指标
2.2 山药不同种植年限土壤微生物数量的变化
由图1可知,在三种菌类中,以细菌所占比例最大,其次为放线菌,真菌最少。与对照相比,细菌数量随着山药连作年限的增加呈递减趋势,表现为CK>T3>T1>T2,其中T1和T2分别较CK降低了8.26%和79.5%;而轮作模式(T3)则促进了土壤中细菌数量的增加,较T2增加了110.5%。放线菌与细菌表现出的趋势相类似,即随着种植年限的增加呈递减趋势,与CK相比,T1,T2分别减少了41.6%,87.9%,其降低幅度较细菌更大,表现出对山药连作更强的敏感性;T3较T2增加了570.5%,差异达极显著水平(p<0.01),说明轮作可以极大地增加连作土壤中放线菌的数量。真菌随连作年限呈先增后降的趋势,与CK相比,T1增加了16.3%;而连作2a后,真菌数量则急剧下降,较T1降低了54.3%,差异达显著水平(p<0.05);T3与T2差异不显著。
2.3 山药不同种植年限土壤酶活性的变化
由图2可知,各处理脲酶活性大小顺序为:T3>CK>T1>T2,表现为随连作年限增加而递减的趋势,而轮作模式可以增加其活性,T3与T2差异显著(p<0.05)。蔗糖酶与脲酶的规律近似,连作2a较1a处理大幅降低,差异达极显著水平(p<0.01),说明其活性受连作年限的影响更为明显;而与连作2a处理相比,轮作可以极显著提高蔗糖酶活性(p<0.01)。碱性磷酸酶随种植年限的增加呈先升后降的趋势,轮作模式较T2有所增加。过氧化氢酶随着种植时间的增加而逐渐增加,轮作模式与T2相差不大。
3 讨论与结论
作物连作对土壤养分状况有较大影响,如刘建国等[12]研究发现,新疆绿洲棉花长期连作后土壤pH、有效钾含量下降,盐分则逐渐增加;吴凤芝等[13]发现设施土壤养分随黄瓜连作年限的增加而呈递增的趋势。前人关于连作对土壤养分影响的研究较多,但作物多样,结论不一,特别是关于山药连作及轮作对土壤养分的影响报道很少。本研究发现山药连作增加了土壤含盐量,而山药—小麦轮作则可以起到降低土壤含盐量的效果;同时山药连作2a较1a明显降低了土壤有机质、全氮、有效磷和速效钾含量,而轮作模式则有增加土壤肥力的作用。
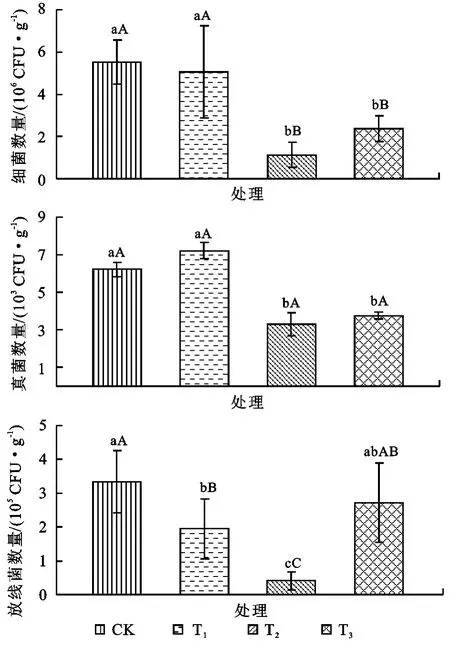
图1 山药连作和轮作模式对土壤主要微生物数量的影响

图2 山药连作和轮作模式对土壤酶活性的影响
前人的研究表明,连作会使土壤从“细菌型”向“真菌型”转化,导致地力衰竭,真菌数量越多土壤肥力越差[14]。就本试验细菌来看,连作减少了细菌数量,这与前人的研究相似;但真菌却表现为随连作年限增加呈先增后降的趋势,这与前人关于连作促使真菌逐年增加的结论不同[15-16]。有研究表明,黄瓜根际细菌和放线菌随连作年限的增加呈“倒马鞍”形变化[17],而温室番茄根际细菌、真菌数量和种类随连作年限的增加而大量减少[18]。由此可见,土壤微生物可能与种植作物、连作年限、日常管理等均有关,从而造成了其变化规律较为复杂。另有研究发现,根系分泌物中的一些含硫化感物质会促进放线菌生长,酚酸等化感物质会严重地抑制真菌生长[19]。山药连作2a后的土壤真菌数量明显降低,这可能与山药根系分泌物累积有关。
土壤酶活性高低可以反映土壤中各种生物化学过程的强 度 和 方 向[20]。前 人 研 究 表 明[21-23],轮 作 对土壤质量具有一定的修复作用,可以改善土壤的理化状况。本研究发现,轮作模式可以激发土壤脲酶、碱性磷酸酶和蔗糖酶的活性,而连作时该三种酶活性明显降低,其中连作2a的土壤脲酶和蔗糖酶活性低于轮作处理,差异分别达显著水平(p<0.05)和极显著水平(p<0.01),这与前人的研究结论相似。而连作则提高了过氧化氢酶活性,虽然与吴凤芝等的研究结论相近[13],但却与许多研究结论相反[17,21],具体原因还待进一步研究。影响土壤酶活性的因素很多,除栽培方式和种植年限外,土壤类型、管理方式等均影响土壤的酶活性。所以本研究认为,山药土壤酶活性受多种因素的控制,因此其只间接地表明某些营养物质的转化情况。
本研究结果表明,山药连作年限对土壤养分、土壤微生物种类和数量产生了显著的影响,这可能是导致山药连作障碍的主要原因之一,而轮作则可以在一定程度上提高脲酶、碱性磷酸酶、蔗糖酶的活性,该研究结论为山药连作障碍的研究提供了一定的数据基础。但有研究表明[24],微生物与根系分泌物的种类和数量存在着密切关系,比如植物根系分泌的酚酸化合物,可直接破坏作物根系细胞膜,或者通过改变土壤微生物类群等,对作物产生抑制作用。因此为了更深入地揭示山药连作障碍,在以后的工作中应重点探讨根系分泌物的种类、数量等同土壤微生物及酶类之间的关系等问题。
[1] 袁书林.山药的化学成分和生物活性作用研究进展[J].食品研究与开发,2008,29(3):176-179.
[2] 王文庆.平遥长山药连作障碍机理研究及其防治对策[D].太原:山西大学,2011.
[3] 刘玮琦,茆振川,杨宇红,等.应用16SrRNA基因文库技术分析土壤细菌群落的多样性[J].微生物学报,2008,48(10):1344-1350.
[4] Yao H,He Z,Wilson M J,et al.Microbial biomass and community structure in a sequence of soils with increasing fertility and changing land use[J].Microbial Ecol.,2000,40(3):223-237.
[5] Benizri E,Baudin E,Guckert A.Root colonization by plant growth promoting Rhizobacteria[J].Biocontrol Sci.Technol.,2001,5:557-574.
[6] Compant S,Duffy B,Nowak J,et al.Use of plant growthpromoting bacteria for biocontrol of plant diseases:principles,mechanisms of action,and future prospects[J].Appl Environ.Microbiol,2005,71(9):4951-4959.
[7] 关松荫.土壤酶与土壤肥力[J].土壤通报,1980,1(6):41-44.
[8] 邱莉萍,刘军,王益权,等.土壤酶活性与土壤肥力的关系研究[J].植物营养与肥料学报,2004,10(3):277-280.
[9] 闵红,张丹,绳金房,等.大棚蔬菜轮/连作系统土壤肥力与微生物因子综合评价[J].微生物学报,2011,38(11):1673-1678.
[10] 刘新晶,许艳丽,李春杰,等.大豆轮作系统对土壤细菌生理菌群的影响[J].大豆科学,2007,26(5):723-727.
[11] 严昶升.土壤肥力研究法[M].北京:农业出版社,1988.
[12] 刘建国,张伟,李彦斌,等.新疆绿洲棉花长期连作对土壤理化性状与土壤酶活性的影响[J].中国农业科学,2009,42(2):725-733.
[13] 吴凤芝,刘德,王东凯,等.大棚蔬菜连作年限对土壤主要理化性状的影响[J].中国蔬菜,1998(4):7-10.
[14] 李春格,李晓鸣,王敬国.大豆连作对土体和根际微生物群落功能的影响[J].生态学报,2006,26(4):1144-1150.
[15] 李琼芳.不同连作年限麦冬根际微生物区系动态研究[J].土壤通报,2006,37(3):563-565.
[16] 邹莉,袁晓颖,李玲,等.连作对大豆根部土壤微生物的影响研究[J].微生物学杂志,2005,25(2):27-30.
[17] 马云华,魏珉,王秀峰.日光温室连作土壤酚类物质变化及其对黄瓜根系抗病性相关酶的影响[J].应用生态学报,2005,16(1):79-82.
[18] 吴凤芝,赵凤艳,谷思玉.保护地黄瓜连作对土壤生物化学性质的影响[J].农业系统科学与综合研究,2002,18(1):20-22.
[19] 胡开辉,罗庆国,汪世华,等.化感水稻根际微生物类群及酶活性变化[J].应用生态学报,2006,17(6):1060-1064.
[20] 姜勇,梁文举,闻大中.免耕对农田土壤生物学特性的影响[J].土壤通报,2004,35(3):347-351.
[21] 吴凤芝,孟立君,王学征.设施蔬菜轮作和连作土壤酶活性的研究[J].植物营养与肥料学报,2006,12(4):554-558.
[22] 赵索,周传余,周超,等.蔬菜不同轮作方式对温室甜瓜产量及根区土壤酶活性的影响[J].黑龙江农业科学,2014(1):73-76.
[23] 李威,程智慧,孟焕文,等.轮作不同蔬菜对大棚番茄连作基质中微生物与酶及后茬番茄的影响[J].园艺学报,2012,39(1):73-80.
[24] 乔卿梅,程茂高,王新民,等.怀山药根际土壤微生物、酶活性和酚酸物质变化及其关系研究[J].中国农学通报,2009,25(24):151-154.
猜你喜欢
今日农业(2021年11期)2021-11-27
现代畜牧科技(2021年4期)2021-07-21
基层中医药(2020年9期)2020-11-27
文苑(2020年6期)2020-06-22
农家科技中旬版(2020年1期)2020-03-12
创新作文(小学版)(2019年22期)2019-12-07
中国化肥信息(2018年12期)2018-03-01
新农业(2017年24期)2018-01-30
中成药(2017年6期)2017-06-13
动物营养学报(2015年3期)2016-01-07
