
长沙市不同森林类型土壤养分含量与土壤酶活性
2014-12-28 06:39黄志宏
中南林业科技大学学报 2014年6期
戴 凌 ,黄志宏 ,2,文 丽
(1. 中南林业科技大学 生命科学与技术学院, 湖南 长沙 410004;2. 南方林业生态应用技术国家工程实验室,湖南 长沙 410004)
长沙市不同森林类型土壤养分含量与土壤酶活性
戴 凌1,黄志宏1,2,文 丽1
(1. 中南林业科技大学 生命科学与技术学院, 湖南 长沙 410004;2. 南方林业生态应用技术国家工程实验室,湖南 长沙 410004)
以湖南省森林植物园内2种林龄的3种典型森林类型为研究对象,研究了有机质、全N、全P、水解N、有效P等土壤化学性质,同时测定了过氧化氢酶、脲酶、磷酸酶等3种土壤酶活性。结果表明: 0~30 cm土层土壤养分各指标含量在不同林型间均存在显著差异(p<0.05),且从高到低的顺序为:樟树+马尾松混交林>樟树林>马尾松林;在不同林龄阶段,全N、水解N、有效P含量存在显著差异(p<0.05),有机质、全P含量不存在显著差异(p>0.05),且从中龄林(24 a生)到成熟林(45 a生),除有机质外,其他指标含量均有下降的趋势;3种土壤酶活性在不同林型(或林龄)间的表征特性与土壤养分各指标含量类似。土壤酶活性与土壤养分因素间密切相关,3种酶能指示土壤养分状况。
森林土壤养分;土壤酶活性;土壤肥力;林型;林龄
森林土壤是维持林木健康生长的基质,其肥力特征影响并控制着林木的健康状态[1]。森林土壤肥力状况,与构成林分的树种及树种组成、植被年龄等林分因子有密切关系。反过来,林分生长状况也能反映土壤肥力高低[2]。土壤酶活性是作为土壤肥力的有效生物学指标之一,可反映土壤中养分转化能力的强弱[3],Puglisi等[4]认为土壤酶活性可作为土壤肥力评价和各种条件下土壤状况的检测指标。同时,有研究表明,土壤酶活性与土壤养分因素密切相关[5]。
不同森林植被下的土壤具有不同的化学性质,研究不同森林植被下土壤化学性质的差异对了解森林与土壤之间的关系,如森林更新、森林的恢复与重建等都具有重要意义。目前,针对森林土壤在维护森林健康中的重要性,不少学者从不同角度对土壤类型[6]、土壤理化性质[7]、土壤酶活性[3-8]等方面进行了一些研究。但从不同林型、林龄对土壤化学特性、土壤酶活性影响的系统性研究并不多见[7]。本研究以24 a和45 a生马尾松林(针叶林)、24 a和45 a生樟树+马尾松混交林(针阔混交林)、24 a和45 a生樟树林(常绿阔叶林)为研究对象,旨在探讨不同林型与林龄下土壤酶活性特征和土壤化学特性及其它们之间的相关关系,借以了解长沙市森林土壤肥力的总体状况,可为长沙市森林土壤的利用、维护和改良提供理论依据。
1 研究区概况
研究地位于湖南省森林植物园(113°02′~03′E,28°06′~07′ N)。该地区气候属于典型的亚热带湿润季风气候,年均气温约为17.2 ℃。1月份最冷,平均温度4.7 ℃,极端最低温度-11.3 ℃;7月份最热,平均温度29.4 ℃,极端最高气温40.6 ℃。年均日照时数为1 677.1 h。雨量充沛,多年平均降水量约为1 422 mm。全年无霜期为270~310 d。试验地海拔50~100 m,坡度12°~21°。土壤主要由第四纪更新世的冲积性网纹红土和砂砾发育而来,属于典型红壤丘陵区。
2 研究方法
2.1 样地设置与土样采集
2013年3月份在湖南省森林植物园林地实验区内,分别设置24 a、45 a生的樟树林、马尾松林及樟树+马尾松混交林(混交林)20 m×20 m的标准样地各3块,总计18块样地。然后在标准样地内按品字形选取4个采样点,每个采样点均按0~30 cm土层采样。土壤样品带回实验室后,自然风干,当风干到一定程度后,按各测定指标的要求进行处理,用铝盒装好冷藏备用。试验使用前将供试土壤样品放在恒温22.5 ℃的烘箱内处理48 h。
2.2 样品化学分析
采用重铬酸钾-水合加热法测定土壤有机质,全N采用半微量凯氏定氮法测定,全P采用盐酸-钼锑抗比色法测定,水解N采用碱解蒸馏法测定,有效P用HCl-H2SO4浸提-钼锑抗比色法测定[9]。
过氧化氢酶活性采用0.1 mol·L-1高锰酸钾滴定法测定,酶活性以常温条件培养30 min后1 g土消耗0.1 mol·L-1KMnO4的毫升数表示;脲酶活性采用比色法测定,酶活性以37 ℃下培养24 h后1 g土释放的NH3-N的毫克数表示;磷酸酶活性采用磷酸苯二钠比色法测定,酶活性以37 ℃下培养24 h后1 g土消耗五氧化二磷的毫克数表示[10]。
2.3 数据处理方法
利用SPSS 16.0统计软件对各指标数据进行正态检验和方程齐次分析。对土壤养分及土壤酶活性各指标采用双因素方差分析(Two-way ANOVA),检验森林类型、植被年龄对土壤养分及酶活性特征影响的显著性。对土壤酶活性与土壤养分,以及土壤酶活性之间的关系进行相关分析。
3 结果与分析
3.1 不同林型土壤养分含量
表1是不同林型0~30 cm土层有机质、全N、全P、水解N、有效P等土壤养分测定的平均值。从表1可以看出,3种林型的土壤有机质含量从高到低的顺序为:混交林(19.91±0.90 g·kg-1)>樟树林(16.55±0.77 g·kg-1)>马尾松林(14.25±0.81 g·kg-1)。方差分析表明,3种林型土壤有机质含量存在显著差异(p<0.05)。土壤有机质含量是衡量土壤健康的重要指标之一[11]。土壤有机质含量在不同林型间存在差异,其因素有多种:不同林型的凋落物量不同,导致土壤有机质含量不同。有研究发现,不同类型的人工林地上凋落物量有差异,这种差异与凋落物量的多少和凋落物的分解速率大小有关[12]。不同类型森林的凋落物分解速率也不同,导致土壤有机质含量也不同。有研究发现,阔叶林地凋落物比针叶林地凋落物分解速率要快些[13]。
森林生态系统中的主要限制营养元素N、P,其主要来源于凋落物的分解[11]。全N含量从高到低的顺序为:混交林(1.19±0.04 g·kg-1)>樟树林(1.16±0.04 g·kg-1)>马尾松林(1.03±0.06 g·kg-1)。方差分析表明,樟树林与混交林全N含量不存在显著差异(p>0.05),但二者均与马尾松林间存在显著差异(p<0.05)。水解N含量从高到低的顺序为:混交林(24.92±1.12 mg·kg-1)>樟树林(19.48±1.12 mg·kg-1)>马尾松林(19.11±0.85 mg·kg-1)。方差分析表明,樟树林与马尾松林水解N含量不存在显著差异(p>0.05),但二者均与混交林间存在显著差异(p<0.05)。可见,林下土壤全N(水解N)含量受林型影响。不同林型土壤全N含量的差异与多种因素有关。一是不同林分凋落物本身初始的C/N比值不一样,导致其养分元素释放率不同[14]。如高志勤等[15]研究发现,竹阔混交林凋落物N素贮量显著高于毛竹林。二是不同林分的凋落物分解速率不同,从而导致地力的不同。三是与凋落物分解速率密切相关的土壤微生物群落结构因林分而异,土壤微生物活动力和数量受林型影响[16-17],土壤微生物多样性与植物群落多样性呈高度正相关[18]。
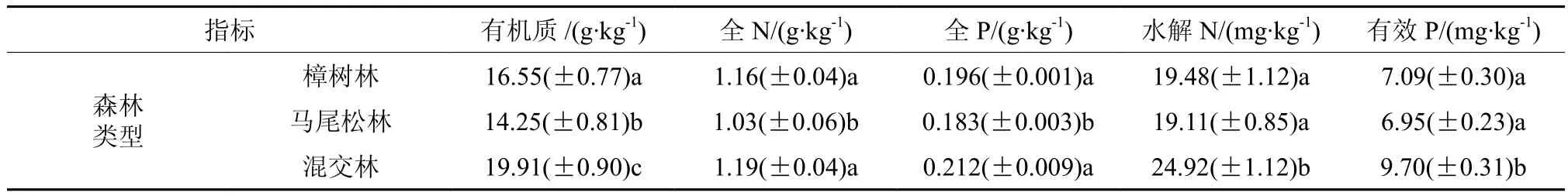
表1 不同森林类型土壤养分含量(平均值±标准误差)†Table 1 Soil nutrients of different type of forests (mean value ± standard error)
全P含量在不同林型间的差异性与全N类似,有效P含量在不同林型间的差异性与水解N类似。可见,林下土壤全P(有效P)含量也受林型影响。不同林型间全P含量高低与多种因素有关。研究表明,阔叶林比针叶林更有利于养分对林地土壤的归还[13]。如南亚热带马尾松混交林林地表层(0~20 cm)土壤全P、速效P的质量分数分别较马尾松纯林增加9.5%、9.9%[19]。本研究中土壤全P(有效P)含量在针阔混交林中最高,阔叶樟树林中次之,马尾松林中最低。这与黄志宏等[20]的阔叶林地土壤肥力高于针阔混交林的研究结果不同。这可能与研究地气候条件的不同有关。因为在区域尺度上凋落物分解主要受气候条件特别是年降雨量和平均气温影响,在林分尺度上则受凋落物化学特性、土壤特性和微生物等因素控制[21]。
3.2 不同林型土壤酶活性
图1是不同林型内0~30 cm土层3种土壤酶活性在3月份测定平均值的比较。从图1可以看出,过氧化氢酶活性在樟树林(1.268±0.025 mL·g-130min-1) 和 马 尾 松 林(1.203±0.017 mL·g-130min-1)间无显著差异,但它们都显著小于混交林(1.401±0.019 mL·g-130min-1)。过氧化氢酶活性与土壤的微生物数量有关[16-17],也与植物根系有关[22],可以用来表征土壤腐殖质化强度大小和有机质的积累程度。混交林地的过氧化氢酶活性高,表明其水解过氧化氢的强度大、碳素循环快和土壤腐殖质合成能力高,从而促进了土壤有机质的积累。

图1 不同林型土壤酶活性比较(A:过氧化氢酶;B:脲酶;C:磷酸酶)Fig.1 Comparison of soil enzymes activity under different type forests (A∶catalase; B∶ urease; C∶ phosphatase)
脲酶能促进有机分子中肽键的水解,其活性可以用于表示N素供应状况[23]。脲酶活性在樟 树 林(0.183±0.009 mg·g-1d-1) 和 马 尾 松 林(0.157±0.009 mg·g-1d-1)间无显著差异,但它们都显著小于混交林(0.225±0.005 mg·g-1d-1)。脲酶活性与有机质和全N含量密切相关[24]。混交林地的有机质和全N含量高,因而脲酶活性也高,这有利于有机分子中肽键的水解,提高土壤中水解N的含量。
磷酸酶活性在3种林型间都有显著性差异(p<0.05),其中,混交林中最高(0.333±0.006 mg·g-1d-1),樟树林(0.306±0.004 mg·g-1d-1)次之,马尾松林最低(0.248±0.064 mg·g-1d-1)。混交林地的磷酸酶活性高,能加速有机磷的循环,提高磷的有效性。樟树林地和马尾松林地磷酸酶活性低,不利于其土壤的有机磷向无机磷转化。
综上可见,土壤酶活性受林型影响。土壤酶活性结果表明,阔叶林地3种土壤酶活性比针叶林地高。这与周庆等[25]的研究结果相同。其可能原因是,一是与凋落物分解程度有关[13],二是与阔叶林地土壤水分条件较好有关,土壤水分状态能够影响土壤酶活性[26]。同时,本研究结果也表明混交林地的3种土壤酶活性比纯林的高。Twanho[27]的研究结果与本研究结果类似。有研究认为,落叶松混交林的土壤养分含量要高于落叶松纯林,导致混交林地的微生物活动力和数量均高于纯林,从而促使混交林地的土壤酶活性高于纯林[16-17]。此外,植物群落的物种多样性和物种组成与土壤酶活性密切相关,林下植物种类多样性高,土壤酶活性相应较高[28-29]。已有研究表明,森林生态系统中植物多样性与过氧化氢酶、转化酶、酸性磷酸酶等酶活性呈正相关[30]。
3.3 不同年龄森林土壤养分含量
表2是不同年龄林分0~30 cm土层土壤养分各指标含量及土壤酶活性测定的平均值。从表2可以看出,土壤有机质含量在24 a生(16.57±0.77 g·kg-1)和 45 a 生(17.23±0.79 g·kg-1)森林内不存在显著差异(p>0.05)。其总体趋势为24 a生<45 a生。这与焦如珍等[31]等的研究结果一致。可能原因是,随着林龄的增长,林分郁闭度增大,光照条件比较差,地被植物少,凋落物多,导致有机质含量增加[32]。
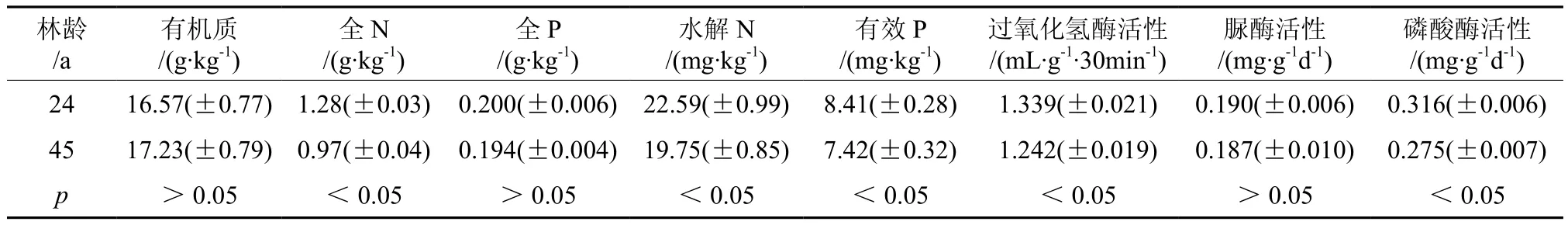
表2 不同年龄森林土壤养分和土壤酶活性(平均值±标准误差)Table 2 Soil nutrients and soil enzymes activity of forests with different stand ages (mean value ± standard error)
土壤全N含量表现出24 a生(1.28 ±0.03 g·kg-1)高于 45 a 生(0.97 ±0.04 g·kg-1),且 2 种林龄间存在显著差异(p<0.05)。水解N含量表现出24 a生(22.59±0.99 mg·kg-1)高于45 a生(19.75±0.85 mg·kg-1),2种林龄间也存在显著差异(p<0.05)。焦如珍等[31]对不同发育阶段杉木林的研究发现,从幼龄林至中龄林至近熟林,土壤速效养分含量呈先下降后上升趋势。这种现象在杨树人工林中也有发现[33]。这些都与本研究结果不一致,可能因为本研究中24 a生的3种森林受人为干扰较多。直接干扰如施加N肥,导致土壤理化性质发生改变;间接干扰如改变了土壤微生物生活坏境。
土壤全P含量表现出24 a生(0.200±0.006 g·kg-1)高于 45 a 生(0.194±0.004 g·kg-1),2 种林龄间不存在显著性差异(p>0.05)。有效P含量表现出 24 a生(8.41±0.28 mg·kg-1)高于 45 a生(7.42±0.32 mg·kg-1),且2种林龄间存在显著性差异(p<0.05)。在落叶松纯林中亦发现类似结果,即随着落叶松林龄的增大,林地土壤全P及有效P含量逐渐降低[34]。
综上可见,从中龄林到成熟林,除有机质外,土壤其他养分含量均有下降的趋势。研究结果与“随着演替的发展土壤养分及酶的活性逐渐升高”这一结论并不一致[35],但与前人[34]的研究结果相似。不同发育阶段林地养分除与林下环境、土壤微生物、及施肥等因素有关外,刘福德等[36]发现,不同发育阶段林地养分还可能与森林起源有关。因此,林龄对土壤养分状况的影响,有待于进一步深入研究。
3.4 不同年龄森林土壤酶活性
从表2可以看出,过氧化氢酶活性表现出24 a生(1.339±0.021 mL·g-130min-1) 森 林 高 于45 a生(1.242±0.019 mL·g-130min-1), 且 2种林龄间存在显著差异(p<0.05)。磷酸酶活性表征特性与过氧化氢酶相同。脲酶活性在24 a生(0.190±0.006 mg·g-1d-1)和 45 a 生 (0.187±0.010 mg·g-1d-1森林内不存在显著差异(p>0.05)。
这表明随着林龄增长,土壤酶活性有下降的趋势。研究表明,根际土壤酸性、中性磷酸酶活性、非根际土壤过氧化氢酶活性、土壤转化酶活性、脲酶等多种土壤酶活性都随着林龄的增加而下降[37]。有研究发现,随着林龄的增加,林地的郁闭度增大,林地光照和水分条件变差,微生物活动减弱,导致近熟林阶段土壤酶活性会有所降低[36]。森林植被可通过改变土壤理化性质、土壤水热状况和土壤生物区系而间接影响到土壤酶活性特征[30]。
3.5 土壤酶活性与土壤养分的相关分析
3种土壤酶活性与土壤各养分指标的相关分析见表3。有研究表明,土壤酶活性与土壤养分间有密切的相关性,并认为土壤酶活性可作为土壤肥力的指标[3-5]。由表3可以看到:过氧化氢酶及磷酸酶活性均与土壤有机质,全N、全P、水解N、有效P含量均极显著正相关。表明过氧化氢酶和磷酸酶对土壤中主要营养物质的转化有一定作用。脲酶活性与土壤有机质、水解N、有效P含量极显著正相关,与全N、全P含量有一定正相关关系,但不显著(r=0.113、0.121,p>0.05)。而秦国宣等[8]发现杉木林中土壤脲酶活性与全N含量有显著正相关关系。这与本研究结果稍有出入,结果的不同可能与本研究样地中人为施加了N肥有关。Ajwa[39]的研究也证实了施加N肥会使土壤脲酶活性有所下降。综上可见,3种土壤酶活性都与有机质、水解N、有效P含量极显著正相关(p<0.001)。说明有机质、水解N、有效P在土壤养分体系和酶体系之间有极为重要的纽带作用。
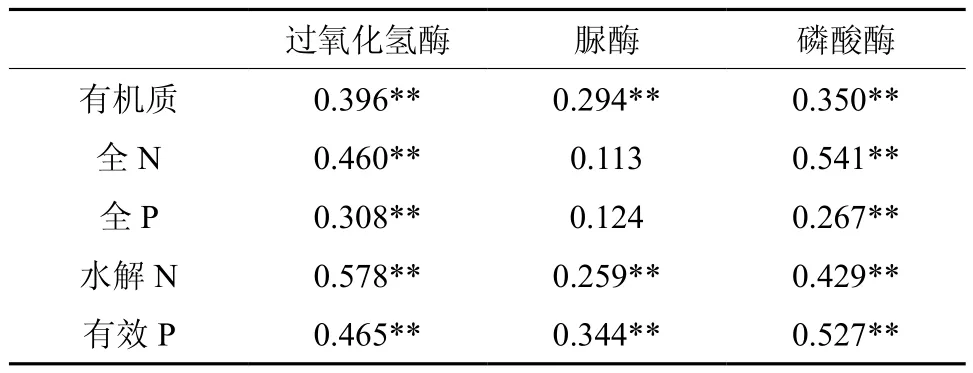
表3 森林土壤酶活性与土壤养分之间的相关性†Table 3 Correlation between soil enzyme activity and soil nutrients
3.6 土壤酶活性相互之间的相关分析
如表4所示,土壤过氧化氢酶活性与土壤脲酶活性极显著相关(r=0.443,p<0.001);土壤过氧化氢酶活性与土壤磷酸酶活性极显著相关(r=0.676,p<0.001);土壤脲酶活性与土壤磷酸酶活性极显著相关(r=0.525,p<0.001)。这表明,过氧化氢酶(还原酶)和其他2种酶(水解酶)在促进土壤有机质的转化过程中,在参与土壤物质转化和能量交换中,它们不仅显示其专有特性,而且还存在共性关系,共同影响土壤养分状况。这与前人的研究结果基本一致[11,40]。说明林地土壤酶活性之间有密切关系,它们不是孤立存在的,而是共同指示土壤养分状况。
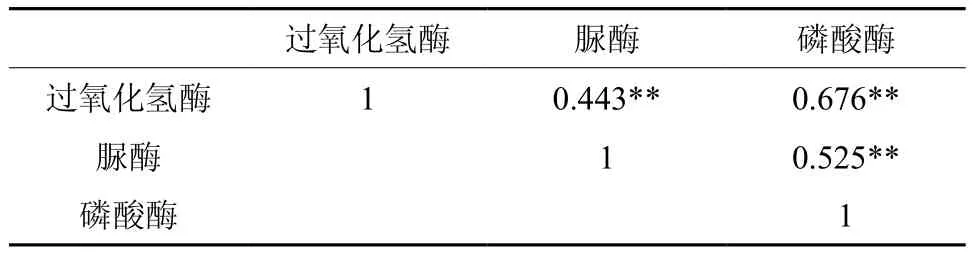
表4 土壤酶活性之间的相关系数矩阵†Table 4 Correlation coefficient matrix between soil enzyme activities
4 结 论
影响森林土壤肥力的因素有多种,如林型、林龄、土壤环境及人为干扰等。不同林型间土壤养分存在显著差异(p<0.05)。3种林型土壤有机质、N(全N、水解N)、P(全P、有效P)含量从高到低的顺序为:樟树+马尾松混交林>樟树林>马尾松林。说明针阔混交林土壤养分各指标含量最高,是当地最佳的林地利用方式。
在森林生态系统中,全N、水解N、有效P含量在不同林龄阶段差异显著(p<0.05),有机质、全P含量差异不显著(p>0.05)。各林型随着林龄的增长,有机质、N(全N、水解N)、P(全P、有效P)等土壤养分指标含量均发生一定变化,除有机质外,其他养分指标含量均表现为24 a生>45 a生。说明从中龄林到成熟林土壤养分有下降的趋势。
不同林型间3种土壤酶活性均存在显著差异(p<0.05);不同林龄间,过氧化氢酶活性与磷酸酶活性存在显著差异,脲酶活性不存在显著差异(p>0.05)。但它们的表征特性与土壤养分类似,这表明土壤酶活性可反映土壤生物化学过程的强度和方向。
土壤化学性质和土壤酶活性之间有着密切联系,它们并不是单独发挥作用的,而是互相作用,彼此共同促进土壤环境的改善。相关分析表明:3种土壤酶活性都与有机质、水解N、有效P含量极显著正相关(p<0.001),3种土壤酶活性间的关系都极显著 (p<0.001)。土壤酶活性能指示土壤养分状况,研究土壤酶作为森林土壤质量综合评价指标具有可行性。
[1] Fisher R F, Binklet D. Ecology and management of forest soils(3rd ed)[J]. New York: John Wiley and Sons, 2000, 282-284.
[2] 王 忠. 植物生理学[M]. 北京: 中国农业出版社, 2000: 121-184.
[3] Bruce A Caldwell. Enzyme activities as a component of soil biodiversity: A review[J]. Pedobiologia, 2005, 49: 637-644.
[4] Puglisi E,et al. Development and validation of numerical indexes integrating enzyme activities of soils[J]. Soil Biology&Biochemistry, 2006, 38: 1673-1681.
[5] 杨万勤, 王开运. 土壤酶研究动态与展望[J]. 应用与环境生物学报, 2002, 8(5): 564-570.
[6] 耿玉清, 余新晓, 岳永杰, 等. 北京山地森林的土壤养分状况[J]. 林业科学,2010, 46 (5):169-175.
[7] 耿玉清, 孙向阳, 等. 长白山林区不同森林类型下土壤肥力状况的研究[J]. 北京林业大学学报, 1991, 21(6):96-103.
[8] 秦国宣, 方 晰, 田大伦, 等. 湖南会同第2代杉木人工林地土壤酶活性[J]. 中南林业科技大学学报, 2008, 28(2):1-7.
[9] 刘光崧. 土壤理化分析与剖面描述[M]. 北京: 中国标准出版社, 1996.
[10] 田大伦. 杉木林生态系统定位研究方法[M]. 北京: 科学出版社, 2004.
[11] Quideau SA, Chadwick OA, Benesi A, et a1. A direct link between forest vegetation type and soil organic matter composition[J]. Geoderma, 2001, 104(1): 41-60.
[12] 邹 碧, 李志安, 丁水祯, 等. 南亚热带4种人工林凋落物动态特征[J]. 生态学报, 2006, 26(3): 715-721.
[13] 李志安, 邹 碧, 丁永祯, 等. 森林凋落物分解速率影响因子及其研究进展[J]. 生态学杂志, 2004, 23(6): 77-83.
[14] 李海涛, 于贵瑞, 李家水, 等. 井冈山森林凋落物分解动态及磷、钾释放速率[J]. 应用生态学报, 2007, 18(2): 233-240.
[15] 高志勤, 傅懋毅. 毛竹林凋落物养分状况的林型变异特征[J].林业科学, 2007, 43(增刊): 95-100.
[16] 薛 立, 陈红跃, 邝立刚. 湿地松林地土壤养分、微生物和酶活性研究[J]. 应用生态学报, 2003, 14(1): 157-159.
[17] Rold·nA, Salinas-Garcl·JR, Alguacil MM, et al. Changes in soil enzyme activity, fertility, aggregation and carbon sequestration mediated by conservation tillage practices and water regime in a maize fi eld[J]. Applied Soil Ecology, 2005, 30: 11-20.
[18] 周 桔, 雷 霆. 土壤微生物多样性影响因素及研究方法的现状与展望[J]. 生物多样性, 2007, 15(3): 306-311.
[19] 刘世忠, 敖惠修, 夏汉平, 等. 赤红壤丘陵水土流失区马尾松纯林改造效益分析[J]. 热带地理, 1999, 9(3): 208-212.
[20] 黄志宏, 田大伦, 周光益, 等. 广东南岭不同林分类型土壤养分状况比较分析[J]. 东北林业大学学报, 2009, 37(9): 63-67.
[21] KÖchy M, Wilson S D. Litter decomposition and nitrogen dynamics in aspen forest and mixed-grass prairie[J]. Ecology,1997, 78(3): 732-739.
[22] Grierson PF, Adams MA. Plant species affect acid phosphatase,ergosterol and mierobial P in a Jarrah (Eucalyptus marginata Donn ex Sm.) forest in southwestern Australia[J]. Soil Biol Biochem, 2000, 32: 1817-1828.
[23] 吴际友, 叶道碧, 王旭军. 长沙市城郊森林土壤酶活性及其与土壤理化性质的相关性[J], 东北林业大学学报, 2010,38(3):97-99.
[24] 刘广深, 徐冬梅, 许中坚, 等. 用通径分析法研究土壤水解酶活性与土壤性质的关系[J]. 土壤学报, 2003, 40(5): 756-762.
[25] 周 庆, 刘有美, 黄锦龙. 桉树林地酶活性研究初报[J]. 华南农业大学学报, 1997, 18(02): 46-50.
[26] 朱同彬, 诸 葛, 玉 平, 等. 不同水肥条件对土壤酶活性的影响[J]. 山东农业科学, 2008, (03): 74-78.
[27] Twanho. Forest Science[M]. 1992, 25(4): 567-568.
[28] 杨万勤, 李瑞智, 韩玉萍. 缙云山天然次生林土壤酶活性的分布特征[G]. 生态学研究论文集(董鸣, M J A Werger编著).重庆: 西南师范大学出版社, 1999a: 171-179.
[29] 杨万勤, 钟章成, 韩玉萍. 缙云山森林土壤酶的分布特征和季节动态及其与四川大头茶的关系[J]. 西南师范大学学报(自然科学版), 1999b, 24(3): 318-324.
[30] 杨万勤, 钟章成, 陶建平,等. 缙云山森林土壤酶活性与植物多样性的关系[J]. 林业科学, 2001, 37(4): 124-128.
[31] 焦如珍, 杨承栋, 屠星南, 等. 杉木人工林不同发育阶段林下植被、土壤微生物、酶活性及养分的变化[J]. 林业科学研究,1997, 10(4): 373-379.
[32] 冯保平, 高润宏, 张秋良, 等. 不同经营方式下兴安落叶松林土壤微生物年季动态研究[J]. 内蒙古农业大学学报, 2009,18(4): 3-4.
[33] 孙翠玲, 朱占学, 王 珍, 等. 杨树人工林退化及维护与提高土壤肥力技术的研究[J]. 林业科学, 1995, 31(6): 506-512.
[34] 陈立新. 陈祥伟, 段文标. 落叶松人工林凋落物与土壤肥力变化的研究[J]. 应用生态学报, 1998, 9(6): 581-586.
[35] Johansson MB. The chemical composition of needle and leaf litter from scots pine, Norway spruce and white bireh in Scandina vian forests[J]. Forests, 1995, 68: 49-62.
[36] 刘福德, 孔令刚, 安树青, 等. 连作杨树人工林不同生长阶段林地内土壤微生态环境特征[J]. 水土保持学报, 2008, 04:121-12.
[37] 耿玉清, 白翠霞, 赵铁蕊, 等. 北京八达岭地区土壤酶活性及其与土壤肥力的关系[J]. 北京林业大学学报, 2006, 28(5):7-11.
[38] 陈立新,杨承栋. 落叶松人工林土壤磷形态、磷酸酶活性演变与林木生长关系的研究[J]. 林业科学,2004, 40(03):12-18.
[39] Ajwa HA. Changes in enzyme activities and microbial biomass of tallgrass prairie soil as related to burning and nitrogen fertilization[J]. Soil Biochemistry, 1999, (31): 769-777.
[40] 陈彩虹, 叶道碧. 4种人工林土壤酶活性与养分的相关性研究[J]. 中南林业科技大学学报, 2010, 30 (6): 64-69.
Soil nutrients and enzyme activity characteristics of different type forests in Changsha city
DAI Ling1, HUANG Zhi-hong1,2, WEN Li1
(1. School of Life Science and Technology, Central South University of Forestry and Technology, Changsha 410004, Hunan China;2. State Engineering Lab. for Applied Forestry Ecological Technology in Southern Regional of China, Changsha 410004, Hunan, China)
∶ By taking the three typical types of forests with two forest-age as the studied objects, the soil chemical properties, including soil pH, organic matter, total N, total P, hydrolytic N and available P were investigated, and three types of soil enzymes activity,such as catalase, urease and phosphatase activity of the three typical type forests in Hunan Botanical Garden with two forest-age in Chanagsha,Hunan province were measured. The results show that there were signif i cant differences of soil fertility content in 0~30 cm soil layer between different forest types in different forest types (p<0.05), and there were a clear consistent trend: Cinnamomum camphora and Pinus massoniana mixed forest>Cinnamomum camphora forest>Pinus massoniana forest; in different forest-age stages, there were signif i cant differences (p<0.05) in the contents of total N, hydrolytic N, available P, while there were no differences in that of organic matter and total P (p>0.05); in addition, from middle age of stand (24-year-old forests) to mature forest (45-year-old forests), except organic matter, other indicators’ contents had dropped trends; the properties of three soil enzymes activity measured here were similar to the soil nutrients indexes content in different forest types (or forest-age); there was a closely related between various soil enzymes and soil nutrients, therefore, the three soil enzymes could be used as the indicators for soil quality assessment.
∶ forest soil nutrients; soil enzyme activity; soil fertility; forest types; forest age
S714.5
A
1673-923X(2014)06-0100-06
2013-12-10
国家自然科学基金(31200346)
戴 凌(1988-),男,湖北荆州人,硕士研究生,主要从事森林生态学方面的研究;E-mail:dailing2933@163.com
黄志宏(1969-),男,湖北天门人,副教授, 博士,主要研究方向为森林生态学;E-mail:huanghugh2013@yahoo.com
[本文编校:文凤鸣]
猜你喜欢
防护林科技(2022年5期)2022-08-22
东南大学学报(自然科学版)(2022年4期)2022-08-12
辽宁林业科技(2021年3期)2021-07-08
食品与发酵工业(2021年9期)2021-05-21
今日中国·西班牙文版(2021年4期)2021-04-09
防护林科技(2020年10期)2020-12-22
腐植酸(2020年1期)2020-11-29
四川林业科技(2020年1期)2020-08-31
浙江理工大学学报(自然科学版)(2018年6期)2018-11-07
现代园艺(2018年2期)2018-03-15
