
毛竹大孢子发生与雌配子体发育研究
2014-12-14 02:50:42郭起荣周建梅孙立方
江西农业大学学报 2014年5期
郭起荣,周建梅,孙立方,2,廉 超,冯 云,冉 洪,张 莹
(1.国际竹藤中心,国家林业局竹藤科学与技术重点开放实验室,北京 100102;2.南开大学 生命科学院,天津 300071)
毛竹(Phyllostachys edulis)属禾本科(Poaceae/Gramineae)竹亚科(Bambusoideae),其开花周期较长,据相关文献记载可达数十年或者百年以上[1-2]。毛竹一生只开一次花、结一次实,开花结实后通常死亡[3]。本文作者调查了广西桂林境内的开花毛竹林分,发现大部分存在着“花而不实”或者结实率低的现象。根据前人的研究发现,造成这种情况的原因可能是雌雄配子体的发育异常、雌雄异熟、传粉有效性低、病虫害等[4-6]。对于竹类植物来说,有性生殖中的每一个阶段都很重要,其中的任何一个阶段出现障碍,都可能造成生殖失败。毛竹的有性生殖研究,国内最早的是乔士义等[7]所报道的毛竹的胚胎发育研究观察,文章初步揭示了毛竹的胚胎学特征。孙立方等[8]对毛竹的花器官形态与结构进行了解剖观察。本文在前人的研究基础之上,采用石蜡切片技术和显微观察,研究毛竹的大孢子发生与雌配子体发育规律,探讨其结实率低下的原因,以期为毛竹的生殖生物学积累材料。
1 材料与方法
1.1 试验材料
试验材料分别于2010—2012 年,在广西桂北海洋山深山野生毛竹林中采集。该地属中亚热带地区,海拔近千米,年平均气温18~19 ℃,年平均降水量1 750~2 000 mm。
1.2 试验方法
标记3 株有开花征兆的毛竹植株,从小枝开始膨大变粗到果实成熟期间,定期采样(6~9 月份),每隔1~3 d 采样,取不同发育时期的小穗、小花、子房、花药,用70% FAA 固定液(冰醋酸:福尔马林体积分数为90∶5∶5)固定24~48 h 以上,4 ℃冰箱保存。采用常规石蜡切片法连续制片[9],切片厚度为5~10 μm,番红-固绿染色,加拿大树胶封片,OLYMPUS CX31 显微镜下观察、拍照。
2 结果与分析
雌蕊是产生卵细胞(雌配子)的场所。雌蕊由2 个心皮组成的,心皮被认为是叶的变态。雌蕊包括子房、花柱和柱头。子房是雌蕊基部膨大的部分,着生在花托上,外围为子房壁,内有一个子室,子室中生有一个胚珠,胚珠是着生在子房内的卵形小体,是由心皮内侧若干部位的细胞经过快速分裂、生长后出现的突起所形成的。每一胚珠由珠心、珠被和珠柄组成。胚珠着生的心皮壁上形成的肉质突起为胎座。卵细胞经受精作用与一个精子融合成合子,以后发育为种子中的胚。
2.1 子房结构
毛竹的子房为一室的单子房,如图1 所示,胚珠单个,倒生,双珠被,胚珠无柄,基部外延先形成内珠被,而后外珠被才形成,在大孢子母细胞与珠心之间有多层细胞,属厚珠心类型,胎座为侧膜胎座。
2.2 大孢子发生
孢原细胞位于珠孔端珠心表皮下方细胞,在珠被覆盖珠心前即已分化,细胞体积较大,细胞质浓厚,具有显著的细胞核,核内具有核仁(图2-A,B)。孢原细胞不形成囊壁细胞,直接发育成为大孢子母细胞,以后大孢子母细胞进行减数分裂,形成了二分体,沿珠孔下方的珠心处向内形成“一”字型排列的四分体(图2-C)。四分体中靠珠孔端一个大孢子发育为功能大孢子,其余的三个退化。
2.3 雌配子体发育
雌配子体的发育开始于成熟的功能大孢子(图2-D)。当功能大孢子成熟后,细胞壁逐渐解体,周围细胞相继相融,同时大孢子细胞核进行第1 次有丝分裂,形成的2 个细胞分别位于核点端与珠孔端,为二核胚囊(图2-E,F),以后二核胚囊进行第2次分裂形成四核胚囊和第3 次分裂形成成熟胚囊(成熟的雌配子体)。胚囊珠孔端具有3 个细胞组成的卵器,包括一个卵细胞和两个靠得较近的助细胞。卵细胞核大,在靠近卵器的位置,是由两个并排的极核组成的中央细胞。在合点端可以观察到大量的反足细胞。这是由于在形成八核胚囊之后,合点端的3 个反足细胞反复分裂而形成的。至此,成熟胚囊形成,雌配子体产生。成熟胚囊由位于珠孔端的卵器、中间的中央细胞以及靠近合点端的多个反足细胞组成。
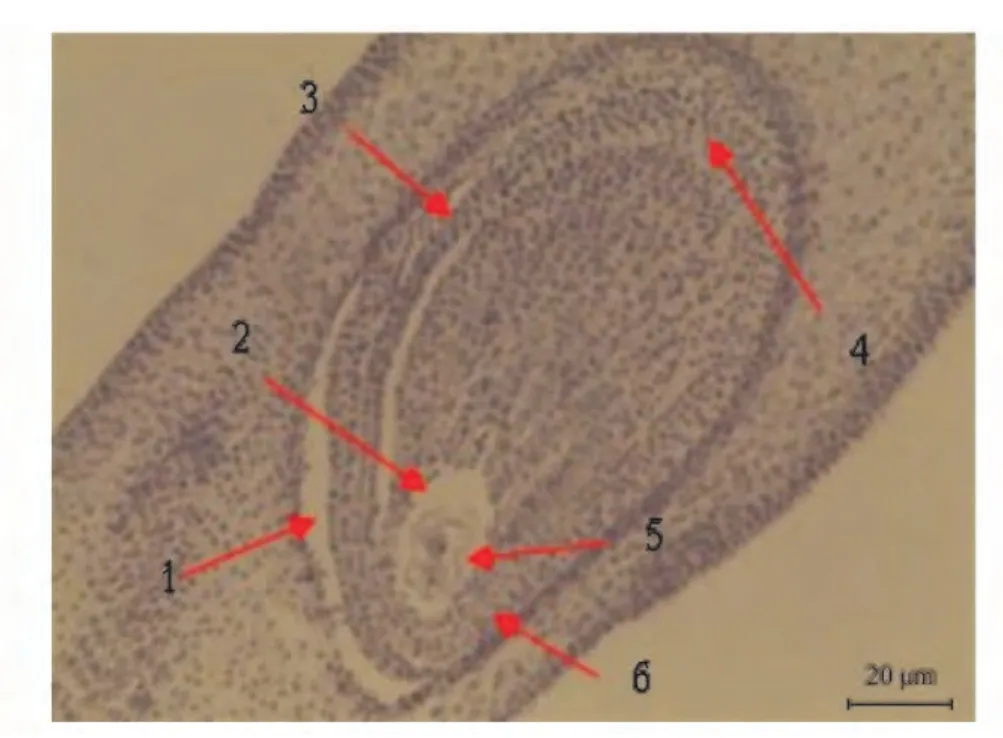
图1 毛竹子房纵切面Fig.1 Section of the ovary of Phyllostachys edulis

图2 毛竹大孢子和胚囊发育Fig.2 The development of megaspore and embryo sac of Phyllostachys edulis
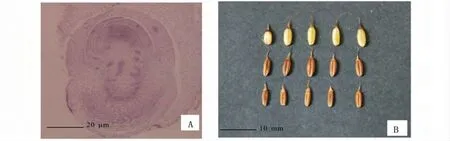
图3 毛竹的成熟胚和颖果Fig.3 Mature embryo and caryopsis of Phyllostachys edulis
2.4 胚和颖果的发育
子房受精后,随着胚和胚乳的发育,珠心、珠被和子房壁发生一系列变化。胚囊不断向珠心深入扩展,珠心逐渐被吸收;同时珠被自外而内、子房壁自内而外被挤毁。配体利用其周围溶解的胚乳组织发育成幼胚,出现盾片、胚根鞘、胚根、胚芽及外胚叶等各器官原基(图3)。当颖果成熟时,珠心几乎全部被胚乳所吸收,果皮仅由栓化的内珠被内层和栓化的子房外壁3~4 层细胞紧密结合而成。
3 结论与讨论
毛竹子房2 心皮1 室,内具1 个倒生胚珠;双珠被,厚珠心。大孢子母细胞由1 个雌性孢原细胞直接发育而成,大孢子四分体呈“一”字型排列;靠珠孔端一个大孢子发育为功能大孢子,由功能大孢子发育为蓼型成熟胚囊,成熟胚囊即成熟的雌配子体包括1 个卵细胞,2 个助细胞,2 个极核组成的中央细胞及大量反足细胞,乔士义和廖光庐在1984 年对毛竹的胚胎发育观察时得出毛竹的胎座为合点胎座,与本文所得结论有所不同。
本文对于毛竹子房的结构特点与之前所报道的雷竹(Phyllostachys violascens)[10]、寒竹(Chimonobambusa marmoreal)[11]、巨龙竹(Dendrocalamus sinicus)[3]及月月竹(Menstruocalamus sichuanensis)[12]的子房构造基本相同,符合禾本科植物子房的特点。
成熟胚囊由位于珠孔端的卵器、中间的中央细胞以及靠近合点端的多个反足细胞组成。助细胞在植物受精过程中起到引导雄配子体进入雌配子体的作用[13],寿命较短,通常在受精作用完成后就破坏。反足细胞具有丰富的细胞质,与珠心相连处具有发达的壁内突,在受精前后逐渐消失,跟胚胎发育的营养有关[14]。
通过对毛竹的大孢子发生及雌配子体的发育研究,发现毛竹大部分胚囊发育正常,不是造成毛竹结实率低的主要原因。林树燕等[12]在月月竹的花药发育研究中发现小孢子收缩变形较严重,可能导致花粉活力变低;庞延军等[4]在爬竹(Drepanostachyum scandeus)花药分化、造孢组织分化、小孢子和雄配子形成时期均发现雄蕊发育异常。因此,雄配子体的发育异常可能是导致竹类植物结实率低的原因之一。在调查过程中,发现开花毛竹林内有病虫害发生,当地农民需要在毛竹开花期内多次喷洒农药,才能保证其正常结实。由此看来,病虫害的发生,也可能是导致毛竹结实率低的原因之一。影响竹类植物结实率的因素是多方面的,除了内部原因,营养或传粉限制等外界环境条件也可能引起结实率低,此问题还需更进一步的研究。
[1]Isagi Y,Shimada K,Kushima H,et al,Watanabe S clonal structure and flowering traits of a bamboo[Phyllostachys pubescens(Mazel)Ohwi]stand grown from a simultaneous flowering as revealed by AFLP analysis[J].Mol Ecol,2004,13:2017-2021.
[2]江泽慧.世界竹藤[M].沈阳:辽宁科技出版社,2002:11-18.
[3]乔士义,廖光庐.毛竹开花生物学特性的观察[J].竹类研究,1984(2):20-24.
[4]庞延军,喻富根,胡成华,等.爬竹雄蕊发育异常的初步观察[J].竹子研究汇刊,1994,13(4):42-46.
[5]何奇江,汪奎宏,华锡奇,等.雷竹花穗和花器的观察研究[J].浙江林业科技,2003,23(2):10-11.
[6]王曙光,普晓兰,丁雨龙,等.巨龙竹生殖器官形态结构及雌雄配子体的发育[J].植物研究,2006,26(3):270-274.
[7]乔士义,廖光庐.毛竹的胚胎发育学观察[J].竹类研究,1984,3(1):15-23.
[8]孙立方,郭起荣,王青,等.毛竹花器官的形态与结构[J].林业科学,2012,48(11):124-129.
[9]李正理.植物组织制片[M].北京:北京大学出版社,1996:129-137.
[10]林新春,袁晓亮,林绕,等.雷竹大孢子发生与雌配子体发育[J].林业科学,2010,46(5):55-57.
[11]胡成华,喻富根,庞延军.寒竹胚胎学观察与研究[J].竹子研究汇刊,1994,13(4):6-11.
[12]林树燕,郝娟娟,辛华,等.月月竹大、小孢子发生和雌、雄配子体发育研究[J].南京林业大学学报:自然科学版,2009(3):9-12.
[13]胡适宜.被子植物生殖生物学[M].北京:高等教育出版社,2005:29-37.
[14]杨弘远.水稻生殖生物学[M].杭州:浙江大学出版社,2005:19-69.
猜你喜欢
果树学报(2023年1期)2023-02-28 19:59:52
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02 01:56:08
西北农林科技大学学报(自然科学版)(2021年5期)2021-05-31 09:34:44
意林·少年版(2020年13期)2020-08-02 11:02:50
中国果树(2020年2期)2020-07-25 02:14:16
草堂(2018年11期)2018-11-13 14:12:16
阅读与作文(小学低年级版)(2016年10期)2016-10-12 09:37:39
中国民族民间医药·下半月(2015年4期)2015-05-22 07:12:43
福建农业科技(2015年1期)2015-02-27 10:20:39
福建农业科技(2015年1期)2015-02-27 10:20:38
