
黄花败酱不同居群的形态多样性分析
2014-12-05 05:31:00张舒娜潘晓曦曲正义刘继勇张志东王英平
吉林中医药 2014年7期
张舒娜,邵 财,张 浩,潘晓曦,马 琳,曲正义,刘继勇,张志东,王英平*
(1.中国农业科学院特产研究所,长春130112;2.康美新开河药业有限公司,吉林集安134200)
种质资源遗传多样性研究,是收集、保存、评价和利用资源的依据,是当今种质资源研究的热点[1]。对于任何一个物种来说,其拥有的遗传多样性越丰富,对环境变化的适应能力就越强。正如Meffe所说,一个群体中遗传多样性的下降意味着其适应环境能力的下降。遗传多样性的丢失可能会导致有机体对环境变化的适应能力降低,同时也使人类丧失了具有潜在应用价值的信息和资源。通过检测了解药用植物的遗传多样性之后,洞悉植物的遗传多样性在居群内与居群间的分布情况,有助于评估特定居群的保护价值[2],及对药用植物制定具体的保护措施。因此,药用植物种质资源遗传多样性的研究对药用植物保护及开发具有十分重要的意义。物质的遗传多样性可以从形态、细胞、生理、DNA序列等不同方面来体现。形态性状是指物种在生长发育过程中,表现出的可观察到的形态特征、特性,如株高、茎粗等,其表现直观,易于识别,无须借助精密仪器,人们对它的应用远远早于其他标记。如利用形态学性状叶长、叶宽对稀有种B homblei和广布种B obovatum研究发现,广布种的遗传变异程度明显大于稀有种。物种的野外调查收集主要是以形态特征为依据的,因此,以形态学为依据建立合理的取样策略及其多样性研究具有现实的意义。但是,药用植物资源取样策略在形态学方面的研究罕见报道。因此,本研究选取黄花败酱(patrinia scabiosaefolia fisch)作为研究对象。黄花败酱是败酱草的正品之一,主产于辽宁、吉林、黑龙江、内蒙古、河北、山东、江西、河南、湖南及云南。其性微寒,味苦、辛,归肺、大肠、肝经。具有清热解毒、破瘀排脓的功效,主治肠痈、肺痈、痢疾、带下、产后瘀滞腹痛、热毒痈肿[3]。现代研究证明其具有镇静、抗菌、抗病毒、抗肿瘤、保肝利胆等作用,临床上用于治疗流行性腮腺炎、鼻窦炎、慢性盆腔炎及阑尾脓肿和结肠炎等[4]。通过对2个黄花败酱居群2个分类学形态性状的多样性分析,试图建立黄花败酱的野外调查、收集取样策略。
1 材料与方法
1.1 材料 黄花败酱由中国农业科学院特产研究所于2013年从吉林省磐石市采集,2个居群原采集地地形均为路边,其具体的地理位置和生态环境见表1。在同一个居群内采取混合取样策略,至少采集50~100个单株,以尽量保证包含不同的基因型[5]。

表1 供试材料
1.2 方法
1.2.1 取样梯度研究 以居群B2为代表,从中随机抽取6、10、14、18、22、26、30 个单株组成7 个梯度抽样群体,对各个抽样群体进行形态学性状调查统计和遗传多样性分析[5]。对2个黄花败酱居群每个居群随机采取30个单株进行形态学性状遗传多样性分析。
1.2.2 形态学性状观测 每个居群以单株为单位,参考形态学分类标准,选取株高、茎粗2个数量性状在成熟期调查。
1.2.3 统计分析 农艺性状数据统计方法以单株为单位,统计农艺性状的最大值(max)、最小值(min)、平均值(mean)、标准差(σ)、变异系数(cv)均由Microsoft Excel软件处理完成。9项农艺性状不同保存群体间差异性比较分析通过SAS软件ANOVA程序完成。利用 Shannon-Weiner index(H')[6]衡量群体遗传多样性大小。具体方法如下:根据总体平均值(mean)和标准差(σ),将数据划分为10级,从第1级[;<(-2σ)]到第10 级[;>(+2σ)],每0.5σ 为一级,计算每一级的相对频率Pi;(即某一性状第i级别内材料份数占总份数的百分比)然后计算各个性状在每个群体中的遗传多样性指数,计算公式为:

其中,Pi指某性状第i级别内材料份数占总份数的百分比,Ln为自然对数。
差异显著性分析采用SAS软件ANOVA程序完成。
2 结果与分析
2.1 不同取样梯度下遗传多样性指数比较 对居群B2的7个梯度抽样群体的2个形态性状进行观测并作遗传多样性分析,见表2。结果表明,不同取样梯度下,各个性状的遗传多样性指数均随单株取样数目的增加呈现增大趋势,总体上来看,当取样数目达到22株时,平均遗传多样性指数最高,为2.007,见图1。

表2 居群B2不同取样梯度下2个形态性状遗传多样性指数
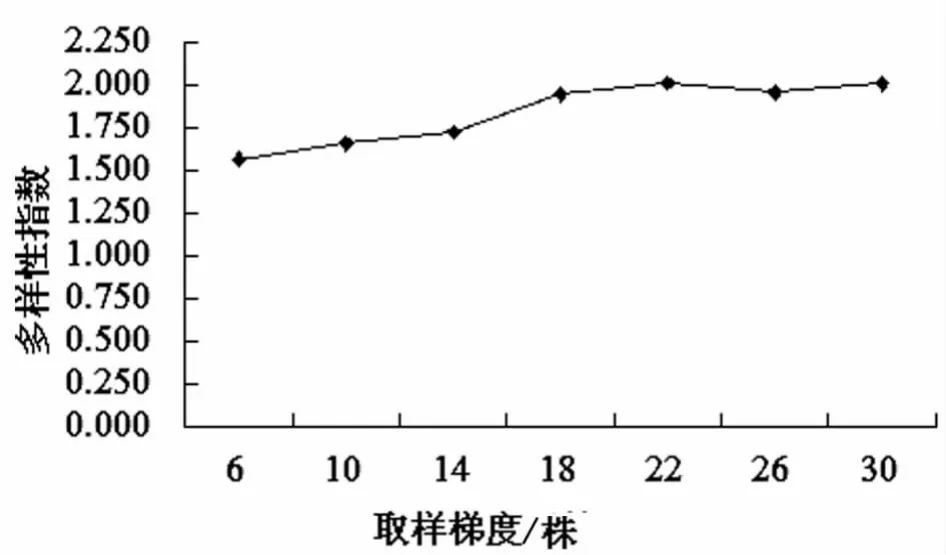
图1 居群B2在8个取样梯度下2个形态性状平均遗传多样性指数的变化
由表2可以看到,株高、茎粗2个性状均在取样量达到22株时遗传多样性指数最高。将2个形态性状综合看,见图1。当取样量为6株时,2个形态性状的平均遗传多样性指数最低,为1.561;当取样量由6株增至30株时,遗传多样性指数随着取样个数的增多呈增大趋势,并且增幅速度很快;当取样量为22株时,平均遗传多样性指数最高,为2.007;当取样量高于22株时,平均遗传多样性指数不再增大,而是在1.953~2.005之间波动。因此,在对黄花败酱居群进行野外考察、收集、保护及多样性研究过程中,每个居群至少要随机选取22个单株才能代表和反映一个居群的整体遗传特性[5]。
2.2 遗传多样性分析
2.2.1 总遗传多样性分析 对2个黄花败酱居群各30个单株2个形态性状数据进行统计分析,见表3。结果表明,不同居群和各性状均表现出丰富的遗传多样性和较高的遗传变异。由表3可见,2个黄花败酱居群总的遗传多样性指数(HT)为2.022,各居群平均遗传多样性指数(Hs)为1.950,占总遗传多样性指数的96.44%;而居群间遗传多样性指数为0.072,只占总遗传多样性指数的3.56%。因此,黄花败酱的遗传多样性主要集中在居群内。
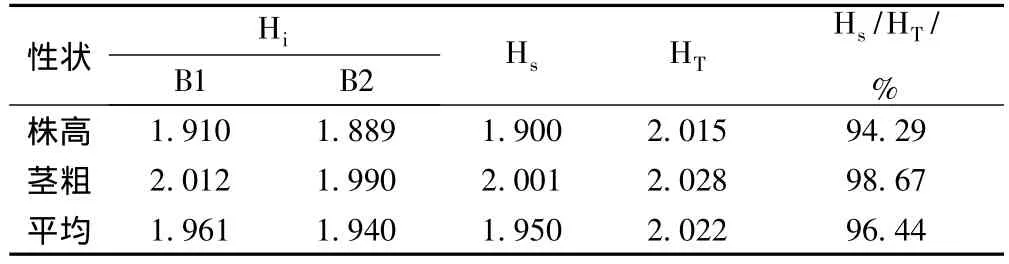
表3 黄花败酱各居群2个形态性状的遗传多样性指数
2.2.2 居群间遗传多样性比较 各居群的遗传多样性分析结果表明,2个居群B1、B2的遗传多样性指数都很高,分别为1.961、1.940。其中居群B1遗传多样性指数最高,表明居群B1具有更大的遗传变异。居群 B1、B2在茎粗上的遗传多样性指数2.012、1.990均高于株高的遗传多样性指数1.910、1.889,表明茎粗具有更大的遗传变异。
2.2.3 居群内个体间的遗传多样性分析 以单株为单位对2个黄花败酱居群进行形态性状的调查统计,结果表明,居群内个体间在两个形态学性状上均存在一定差异。在2个居群中,B1的遗传多样性表现最为丰富,该居群2个性状株高、茎粗的遗传多样性指数分别为1.910、2.012,其中茎粗的遗传多样性指数是所有形态性状中最高的,其平均值为5.09 mm,变异在3.07~8.61 mm之间,变异幅度达5.54 mm,变异系数达23.93%;大的变异幅度和高的变异系数说明,在居群B1内不同的个体间存在差异,见表4。
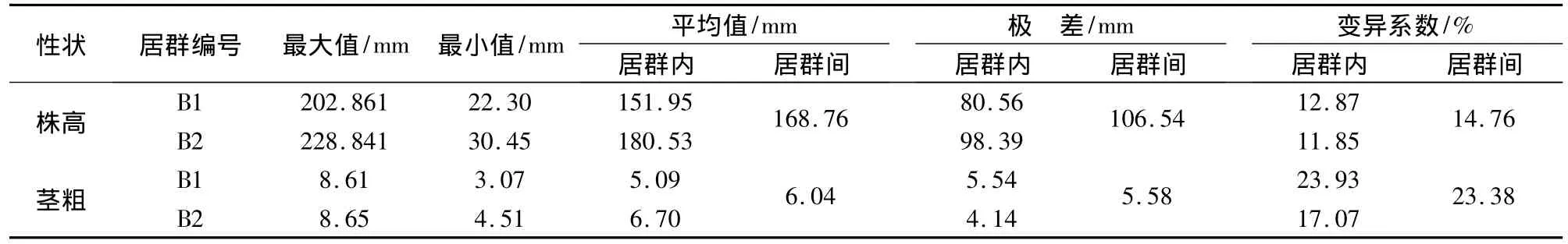
表4 黄花败酱居群2个形态性状表现
2.2.4 2个形态性状的遗传多样性比较 将2个居群全部个体的2个形态性状的遗传多样性进行分析,结果表明,2个形态学性状的遗传变异均主要存在于居群内个体间,不同形态性状的遗传多样性表现不同。不同性状的居群平均遗传多样性指数占该性状总遗传多样性指数的比率(Hs/HT)在94.29% ~98.67%之间。茎粗的居群平均遗传多样性指数(2.001)占总遗传多样性指数(2.028)的比例达到了98.67%,是居群平均遗传多样性指数最高的。
3 结论
遗传多样性的取样策略是指对一定地理分布范围内的生物个体取样时,使样本具有代表性和包含尽可能多的遗传变异的最佳取样方法,包括取样数目(一个给定区域的居群数和一个居群的个体数)以及取样方式[7]。不同类型植物的取样策略不同,在不同采样方法对细距堇菜遗传估测研究表明,每群体取30株时有较充分的代表性,小于20个个体的取样可能引起遗传多样性估测出现大的偏差[8]。史冀伟等[5]对小麦族中间鹅观草居群的形态多样性分析表明,对于小麦族自花授粉植物野外调查、收集时,应以居群为单位,而且每一居群至少应调查、收集18个单株,才能代表居群的遗传多样性。Marshall等[9]认为在不知道物种生物学特性和遗传背景的情况下,对一个居群进行资源保护的随机取样数目可以确定在30个左右。本文针对黄花败酱居群2个形态性状进行了取样策略研究,结果表明,不同取样梯度下的遗传多样性指数随单株取样数目的增加呈现增大趋势,当取样量为22株时,平均遗传多样性指数最高(2.007),当取样量为22~30株时,平均遗传多样性指数在1.953~2.005之间波动。因此,建议在进行黄花败酱居群形态学多样性研究中,单个居群要至少保证22株作为分析单位。这为黄花败酱物种以居群为单位的遗传多样性研究提供了取样策略的依据。
黄花败酱形态变异的丰富性,不仅体现在不同的居群间和不同的性状上,还表现在居群内个体间的差异上,不同个体间存在丰富的变异。比如株高变异在130.45~228.84 mm之间,变异幅度达98.39 mm。如此大的变异范围,要求在野外调查、收集和研究以及种质资源的利用中以居群为单位、以个体取样是非常有必要的[10]。
[1]董玉琛.生物多样性及作物遗传多样性检测[J].作物品种资源,1995(3):1.
[2]唐炎林,苏智先,张军.遗传多样性的检测途径及其对濒危植物保护的意义[J].内江师范学院学报,2004,10(2):38-41.
[3]南京中医药大学.中药大辞典[M].上海:上海科学技术出版社,2006:1914.
[4]王盈.黄花败酱的化学成分及药理作用研究进展[J].齐鲁药事,2009,28(4):222-225.
[5]史冀伟,高爱农,刘津刚,等.小麦族中间鹅观草不同居群的形态多样性分析[J].植物遗传资源学报,2009,10(4):547-552.
[6]Shannon C E,Weaver W.The mathematical theory of communication[M].Urbana:University of Illinois Press,1949:3-14.
[7]金燕,卢宝荣.遗传多样性的取样策略[J].生物多样性,2003,11(2):155-161.
[8]李昂,王可青,葛颂.不同采样策略对细距堇菜遗传多样性估算的影响[J].植物学报,2000,42(10):1069-1074
[9]Marshall D R,Brown A H D.Optimum samp ling strategies in genetic conservation[M]//Frankel O H,Hawles JG.Crop genetic resource for today and tomorrow.London:Cambridge University Press,1975:53-80.
[10]史冀伟.小麦野生近缘植物中间鹅观草的遗传多样性与基因组构成分析[D].北京:中国农业科学院,2009.
猜你喜欢
Journal of Traditional Chinese Medicine(2022年1期)2022-11-17 06:52:50
今日农业(2022年14期)2022-09-15 01:44:50
河北科技师范学院学报(2022年2期)2022-08-26 08:55:24
今日农业(2022年3期)2022-06-05 07:12:00
浙江中医药大学学报(2021年6期)2021-07-12 03:05:14
亚太传统医药(2020年12期)2021-01-11 07:28:00
黄河之声(2020年16期)2020-11-05 01:57:46
今日农业(2020年16期)2020-09-25 03:04:32
草地学报(2018年5期)2018-11-07 02:25:00
中老年健康(2017年9期)2017-12-13 15:24:20
