
烟台潮间带大型底栖动物群落组成和结构研究
2014-12-02 03:09:46韩庆喜袁泽轶陈丙见王玉珏石雅君刘东艳
海洋科学 2014年9期
韩庆喜, 袁泽轶, 陈丙见, 王玉珏, 石雅君, 刘东艳
(1. 中国科学院 烟台海岸带研究所 海岸带环境过程与生态修复重点实验室, 山东省海岸带环境过程重点实验室, 山东 烟台 264003; 2. 国家海洋信息中心, 天津 300171; 3. 上海海洋大学 海洋科学学院, 上海201306)
潮间带作为海洋湿地生态系统一个重要类型,地处陆地与海洋生态系统作用较强烈的地带, 陆海理化因子交替, 环境复杂多变[1], 通常具有丰富的底栖群落, 为海鸟、底栖鱼类和甲壳类等提供的充足的食物来源和产卵育雏地[2-3], 另外人类生产开发活动频繁, 对潮间带生态系统的影响和扰动也与日俱增,因此潮间带生态学的研究越来越受关注。大型底栖动物作为海岸带生态系统的重要组成部分, 在潮间带生态系统碎屑分解、物质循环、能量流动以及生态系统平衡与稳定中起着重要作用[4-5]。大型底栖动物具有相对稳定的生活环境和较差的稳定性, 群落组成和结构对物理环境和人类扰动敏感而深刻[6-7], 因此,是监测海岸带环境的良好的指示生物[8-9], 是研究环境压力对生态系统综合影响的最佳选择[10-12]。
随着烟台人口的急剧增加以及经济的迅速发展,烟台潮间带自20世纪30年代有记录以来发生了很多变化,更深刻的变化来自 20世纪 80年代至今, 现如今沿烟台海岸遍布的大量海草床已消亡殆尽, 四十里湾潮间带海草床仅在烟台渔人码头附近有斑块分布。烟台潮间带从四十里湾北部芝罘岛向南一直到养马岛, 软泥质潮间带、泥沙质潮间带及沙质潮间带均有分布。为使本研究全面而有代表性, 选取了 3个不同类型的潮间带进行取样, 分析烟台滨海潮间带底栖生物的变化。选取的这3种受不同扰动或影响的潮间带类型, 通过比较长期的历史资料,发现其群落演化和生态变迁, 为合理持续开发潮间带生物资源以及保护海岸带健康的生态环境提供依据。
1 材料与方法
1.1 研究区域简介
烟台地处胶东半岛北部。三面环山, 北面濒临黄海, 隔海与辽东半岛相望。烟台海岸曲折蜿蜒构成许多内湾, 沿岸属“基岩海岸”类型, 包括岩岸、砂岸、泥砂岸、砾石滩、海港(泥沙、岩石底质)5种生境, 丰富的生境是海洋动物栖息、产卵和繁殖的良好场所。本次调查选择了 3种最常见的基质类型, 包括辛安河口的沙质底质, 辛安河口的泥沙底质以及养马岛南部的泥质滩涂, 这3处采样点的具体位置如下: 辛安河 口 37°26′25.71′N, 121°32′51.74′E; 渔 人 码 头37°30′52.41′N, 121°26′38.46′E; 养 马 岛 37°27′3.82′N,121°35′49.17′E。调查站位地理位置如图 1 所示。

图1 2009~2010年烟台潮间带调查采样站位分布图Fig.1 Position of sampling site in Yantai intertidal zone during year 2009~2010
1.2 采样方法
本次调查于2009年11月, 2010年2月、5月和8月4个季度月(秋、冬、春、夏)在烟台海域辛安河口和渔人码头近岸海域潮间带进行采集。在辛安河口与渔人码头潮间带取一垂直于海岸的断面, 从高潮的基准面下退15 m至低潮区附近等距离设 3个站点 (高、中、低潮区) , 用 GPS定位仪定位以便 4个季度重复取样, 对大型底栖生物进行调查取样。
调查区域范围是烟台滨海潮间带, 调查选取具有代表性的3个断面, 每个断面在高、中、低潮带各设 1个站, 必要的断面将增加潮下带和潮上带站位以增强对潮间带底栖生物群落特征的理解, 先期进行 1年的调查取样, 以期能获得烟台滨海潮间带底栖生物的群落特征和受人类影响干扰情况, 并为未来进行潮间带底栖生物群落演替作一个本底的参考。调查时间定为2009年11月和2010年2月、5月、8月, 每季度野外调查一次, 采获的底栖生物样品拿回实验室进行室内研究分析, 包括鉴定、计数、称质量, 调查规范依据《海洋监测规范》第七部分。数据分析主要利用各种生物统计软件, 包括PRIMER, SPSS AMBI软件包和EXCEL进行数据的处理和作分布图。
采集的生物样品为大型底栖动物, 分为定量样品和定性样品。定量样品采集根据采用0.5 m×0.5m的取样框, 对 0.25 m2底质内的样品进行采集, 用0.5 mm 网目的筛子进行冲洗, 获得大型底栖生物,所得样品用 75%的酒精进行固定后带回实验室。在实验室用 1%的虎红溶液对筛选所得的大型底栖动物标本和残渣进行染色挑选, 最后进行鉴定、计数及称质量。易断的纽虫和多毛类按头部计数, 管栖动物剥去管子, 软体动物的死壳不计数, 软体动物不去贝壳, 群体标本不计算个数, 寄居蟹去螺壳称质量。分类计数后的标本用精度为1 mg的电子天平称量。并换算求得各种群的优势度、栖息密度(个/m2)和生物量(g/m2)。样品的处理、鉴定、称质量等均按《海洋调查规范》(GB 12763.6-91)进行。同时对底质的粒径组成、有机质含量进行分析。
1.3 数据处理
底栖动物群落数据在标准化后采用单因子和多因子统计分析进行群落结构分析。底栖群落的生物特征包括丰度(A), 物种数目 (S), 香农威纳指数(H′;log10), 多样性指数 (d)和均匀度指数 (J′) 。
优势种的计算: 优势种的定义在不同资料、文献中有所区别, 在应用研究中, 对优势种主要通过一系列量化标准进行划分, 本文采用优势度 Y[13]作为划分优势种的标准。大型底栖生物优势度计算公式为:
Y=(ni/N)×fi
式中, N为采泥样品中所有种类的总个体数目, ni为第i种的个体数; fi为该种在各站位出现的频率; 当物种优势度Y > 0.02时, 该种即为优势种。
依据 Gaudêncio 等[14]的粒径分析标准, 按粒径大小将底质分为砾石(≥2.000 mm, 粗砂(0.500~2.000 mm),细砂(0.250~0.063)和泥(<0.063 mm)。粒径采用 Mastersizer 2000型(马尔文仪器有限公司, 英国)激光粒度仪测定, 粒径测量范围为0.02~2000µm, 相对误差小于1%。
运用普利茅斯海洋实验室开发的PRIMERTM6.0软件包, 进行群落组成及结构的统计分析, 以研究分析物种丰度的时空变化。底栖丰度数据经过平方根转化之后, 采用Bray-Curtis相似性矩阵进行转换,采用组平均排序和非度量多维测度进行等级聚类。采用 AMBI 5.0软件包(下载于 www.azti.es)中的AZTI’s Marine Biotic Index (AMBI)和 multivariate-AMBI (m-AMBI) 分析底栖生态状况[15]。AMBI阈值如下: “正常”, 0.0~1.2; “轻微污染”, 1.2~3.3; “扰动”,3.3~5.0; “剧烈扰动”, 5.0~6.0 和“极端扰动”以及“无生命”[16]。m-AMBI的质量阈值如下: “高”>0.77; “良好”0.53~0.77; “中等”0.38~0.53; “差”0.20~0.38; 和“恶劣” <0.20[17]。
各采样区域的底质粒径分布见表1。

表1 烟台潮间带各采样区域的底质粒径分布Tab.1 Grain size of different intertidal zones in Yantai
2 结果
2.1 种类组成及季节变化
根据在烟台潮间带3个典型潮间带4个季节的大型底栖动物采集, 共获得 9155号标本, 分属 111种, 包括多毛类动物40种, 甲壳动物37种, 软体动物29种, 棘皮动物1种, 鱼类2种, 其他动物2种(包括纽虫和腔肠动物各1种)。多毛类动物和甲壳动物占据了研究区域底栖群落的主要成分, 两者分别占据了群落总种数的36.04%和33.33%, 软体动物同样也是该区域潮间带底栖群落的重要组成部分, 它占据了群落总种数的26.13%。泥沙砾石的渔人码头潮间带总种数为70种, 远高于沙质的辛安河口34种和泥质的养马岛泥质潮间带36种。在各季节的类群分布中, 甲壳动物在冬季种类数量要少于其他 3个季节, 在3个潮间带区域的种数分别为1种(渔人码头),0种(辛安河口)和 1种(养马岛); 并且冬季的总种数在4个季节中也是最低的(表2)。

表2 烟台潮间带各季节大型底栖动物物种数分布Tab.2 Seasonal distribution of macrobenthos in intertidal zones in Yantai
2.2 大型底栖动物的丰度与生物量
烟台潮间带 3个典型潮间带大型底栖动物平均丰度为(2034.44±2042.58)个/m2, 平均生物量为(198.86±191.82)g/m2。不同动物类群的丰度的时空差异都很显著, 泥沙砾石性质的渔人码头潮间带丰度最大, 占据了 60.6%, 尤其以日本大螯蜚丰度最大,冬季的丰度达到 1768.67个/m2。沙质的辛安河口则以多毛类为主, 而泥质的养马岛则以软体动物为主。丰度的季节变化中, 春、夏和秋季3个季节差异较小,而冬季则变化剧烈。渔人码头和养马岛冬季的丰度要远高于其他 3个季节, 而辛安河口的冬季丰度则明显小于其他 3个季节。不同动物类群生物量的空间差异显著, 平均生物量以养马岛最高(103.87g/m2),渔人码头次之(32.66 g/m2), 辛安河口最低(12.62 g/m2)。各个动物类群在不同季节的生物量略有不同, 渔人码头和辛安河口均以秋季的生物量为最高, 而养马岛潮间带的生物量在冬季达到最高, 远高于其他3个季节。3个典型潮间带中, 软体动物均是生物量的主要贡献者, 除渔人码头秋季(40%)和辛安河口夏季(33%)外, 软体动物在总生物量的比重均超过59%(详见表 3)。
2.3 优势种的时空分布
在本次调查研究中, 烟台潮间带出现的优势种共15种, 包括6种多毛类, 5种软体动物, 4种甲壳动物, 但多数优势种仅在某个潮间带的单个季节出现,优势种的季节变化明显, 包括多毛类动物的长双须虫和日本刺沙蚕, 软体动物的等边浅蛤、西施舌和异白樱蛤, 甲壳动物的伍氏厚蟹、大蝼蛄虾以及长趾股窗蟹, 而在多个季节或潮间带出现的优势种仅有 6种, 包括多毛类动物的中型阿曼吉虫、膜质伪才女虫和小头虫, 软体动物的菲律宾蛤仔和古氏滩栖螺,甲壳动物的日本大螯蜚。在渔人码头大型底栖动物群落中, 日本大螯蜚(优势度为 0.29~0.82)是全年的优势种, 是该群落的重要组成部分; 其次为小头虫,是除夏季之外其他 3个季节的优势种。中型阿曼吉虫则是辛安河口大型底栖动物群落的主要优势物种,它在春、夏、秋季均为该研究区域的优势种。养马岛的底栖群落优势种则为菲律宾蛤仔, 在春、夏、冬季的优势度为0.07~0.48(表4)。
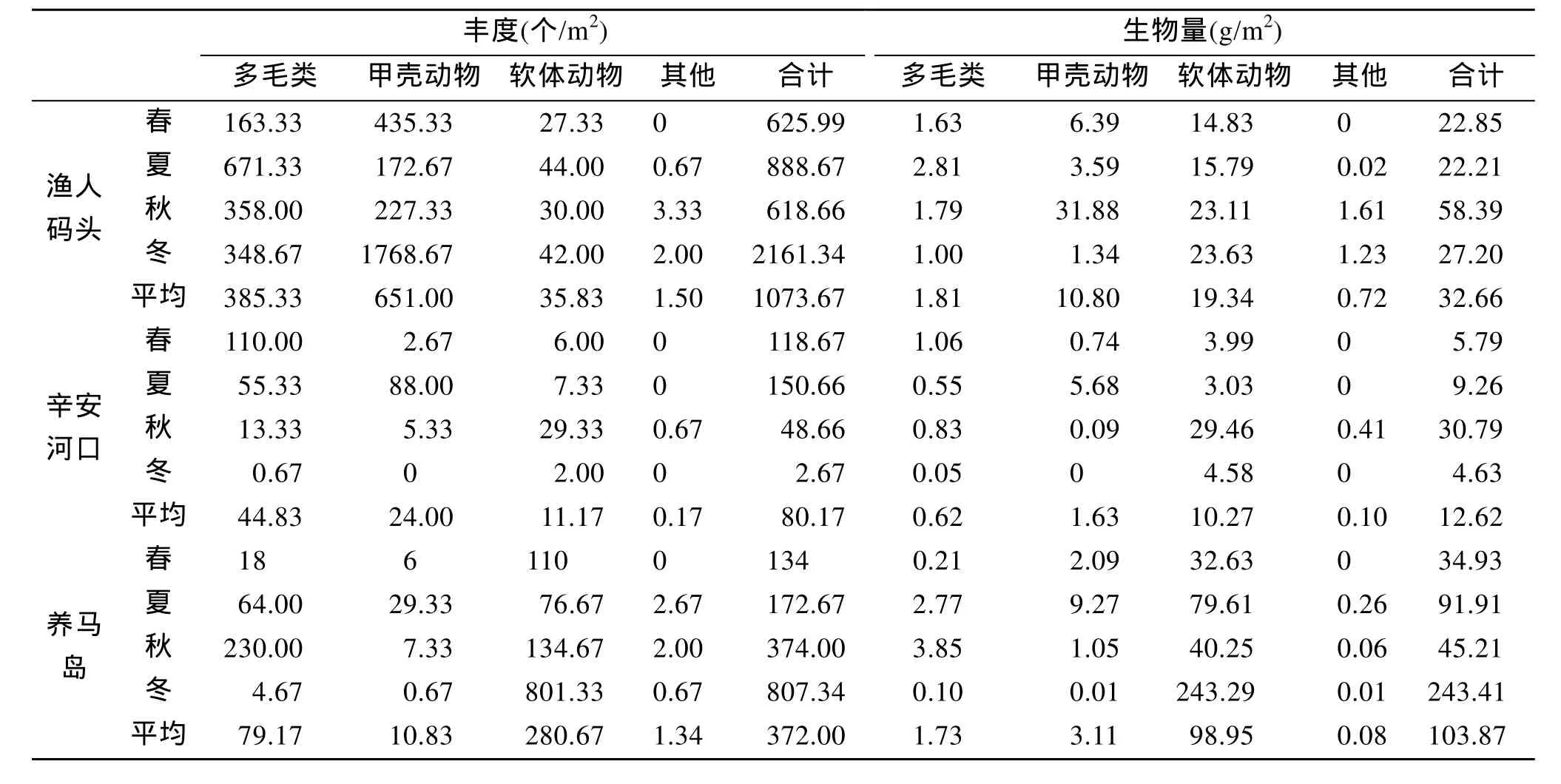
表3 烟台潮间带大型底栖动物生物量和栖息密度分布Tab.3 Seasonal distribution of macrobenthic abundance and biomass in intertidal zones in Yantai
2.4 群落结构的时空变化
为了避免稀有种的相对丰度对可靠性的影响,以提高非参数检验的强壮性, 将所有占总丰度0.05%以下的种剔除[18]。在后面的这些分析中, 这些种的丰度占到了所采样品总丰度的99.21%。在对群落数据进行了二次方根转化之后, 转化为 Bray-Curtis矩阵, 进行Cluster聚类, 结果(图2)表明, 在3种不同的底质类型下, 沙质的辛安河口底栖群落受的季节变迁异常剧烈; 而渔人码头和养马岛底栖生物类群的季节变化维持在一个相对稳定的水平上,两者不同的底质类型决定了底栖群落的结构和组成,季节因素对其施加的影响在底质之下。在渔人码头,春季和夏季的群落结构相类似, 而养马岛底栖群落则在秋冬季较为相近。
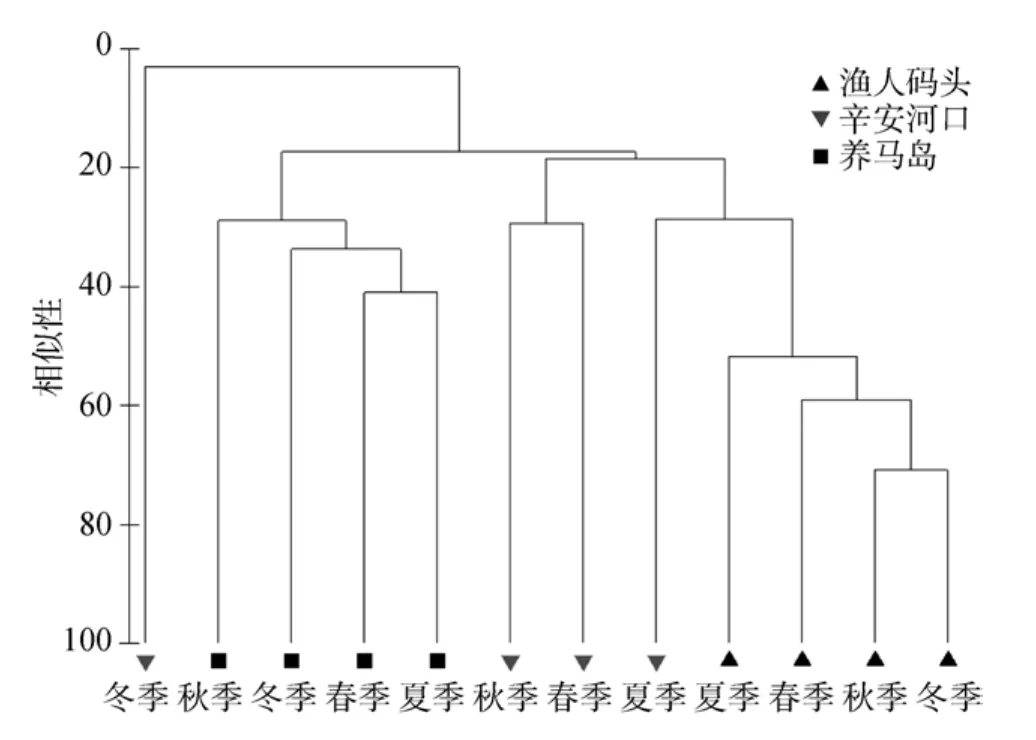
图2 烟台潮间带大型底栖动物群落的聚类分析Fig.2 Cluster analysis of macrobenthic community in different intertidal zones in Yantai
作者对各采样区域和各季节的底栖群落进行SIMPER分析, 结果也与以上结果相符。对各采样区域的 SIMPER分析显示, 渔人码头不同季节的群落的平均相似性为57.45, 远大于养马岛的32.60和辛安河口的15.72。渔人码头的平均相似性的主要贡献者是Grandidierella japonica Stephensen, 1938, Capitella capitata (Fabricius, 1780)和Armandia intermedia Fauvel, 1902, 它们对平均相似性的贡献率均超过10%。养马岛底栖群落平均相似性的主要贡献者是Ruditapes philippinarum (Adams & Reeve, 1850),Macoma incongrua (Martens, 1865), Batillaria cumingi (Crosse, 1862), Lumbrineris latreilli Audouin et Edwards, 1834, 辛安河口平均相似性的主要贡献者为Armandia intermedia Fauvel, 1902, Ruditapes philippinarum (Adams & Reeve, 1850), Gomphina aequilatera (Sowerby, 1825)和Pseudopolydora kempi(Southern, 1921), 这些种也是各采样区域群落丰度的主要贡献者, 除渔人码头的Pseudopolydora kempi(Southern, 1921)似外, 它的丰度贡献值超过10.28%,但对平均相似性的贡献率只有 6.05%, 它的季节变化异常剧烈, 夏季的丰度值占到了全年总丰度的93.89%。作者对不同季节的底栖群落的 SIMPER分析显示, 各季节在不同采样区域的平均相似性均比较低, 春季最高, 仅为22.62, 冬季最低, 仅4.96, 夏季和秋季分别是20.89和22.28。
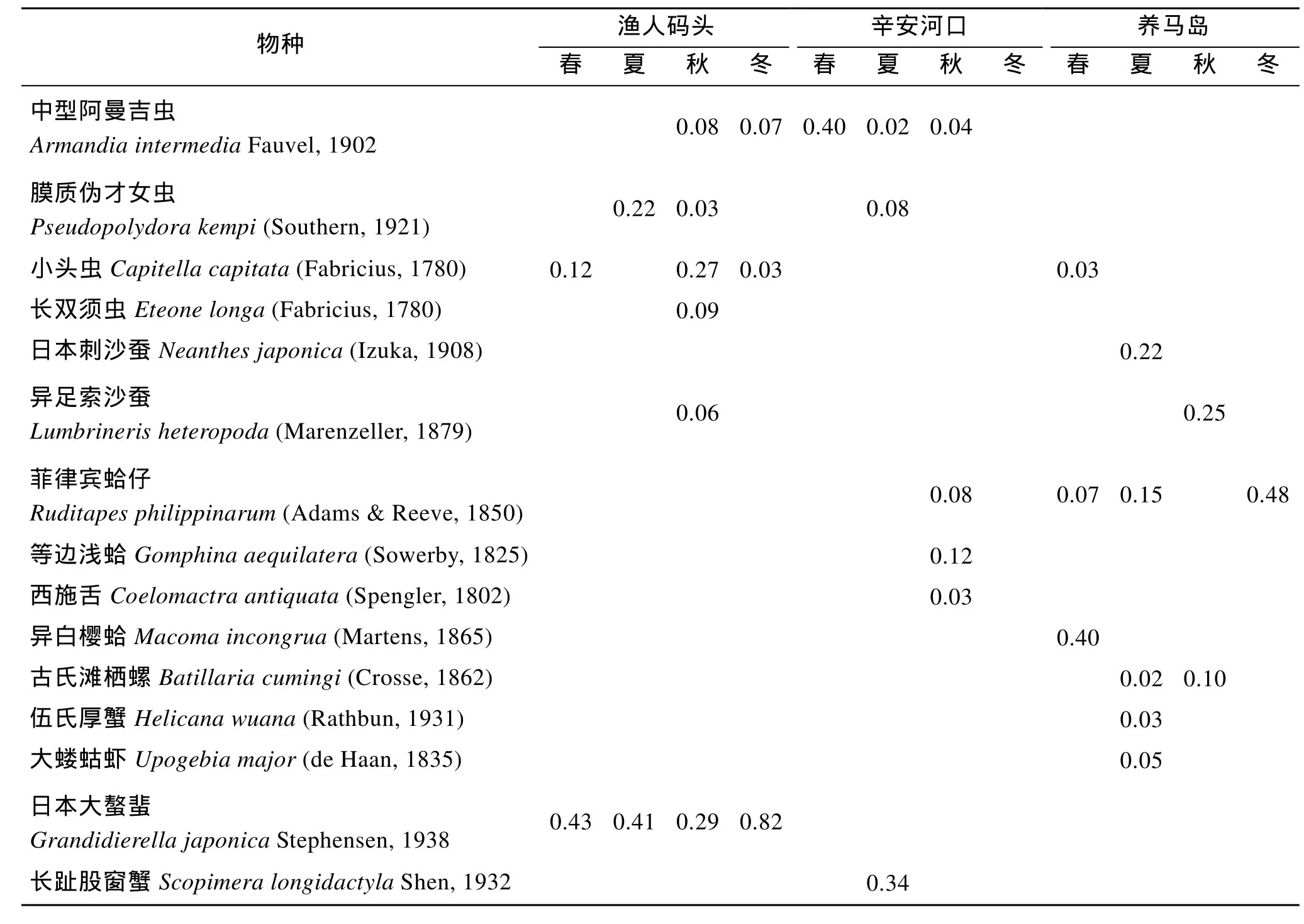
表4 烟台潮间带大型底栖动物主要优势种及其优势度Tab.4 Dominant species and its seasonal dominance in different intertidal zones in Yantai
2.5 底栖群落与环境扰动
由物种丰度数据进行了多样性、AMBI和 m-AMBI的计算, 结果(表 5)显示, 除辛安河口由于物种数的季节变化较大(比如冬季仅获得3种底栖动物)使得评估结果的可信度存在一定问题之外, 渔人码头泥沙质海滩和养马岛泥滩均受到了人类活动的轻微扰动, 同时生态质量状况处于相对良好的水平。AMBI指数建立在某一站位各底栖动物栖息密度所占比例的基础之上, 评估结果受优势物种的影响较大, 在物种数和生物多样性指数较低的辛安河口(冬季), 当某一站位的底栖动物种类数和个体数较少时,AMBI的敏感性会明显降低[19], 而综合了AMBI、物种多样性指数和物种数的m-AMBI对生态环境质量的评估结果较之更为准确[20]。
3 讨论
烟台底栖生物从张玺[21]和张修吉[22]的种类记述开始, 在他们的早期工作中, 对烟台潮间带及浅海的底栖动物种类进行了种类图示描绘及优势种的记述, 但其在采样过程中并不严格区分潮间带与浅海,张玺[21]对烟台各类型浅海的代表性底栖生物进行了种类记述, 某些种类并附有精美的外形图。张修吉[22]统计了烟台海滨标本计有112科, 131属, 并对一些稀有物种进行了形态和生境描述。但在渔人码头附近仅记录4种, 包括沙蚕科沙蚕属Nereis, 鳞沙蚕科Polynoë pontogema 及矶沙蚕 Eunicidae Arabella。杨宗岱和吴宝铃[23]记录了渔人码头附近多毛类 14种,明显少于本次调查所获得的28种。丛建国[24]根据每年1次连续7年的调查, 获得无脊椎动物128种, 与本次研究相比, 他的报道也涉及了岩石滩和海港码头, 通过比较香农威纳指数显示, 无脊椎动物群落的多样性、均匀度和种类丰富度及个体数量在急剧下降, 沙质海滩上的美人虾Callianassa sp.及海棒槌Paracaudina chilensis ransonnetii已绝迹, 在本次调查中亦未有发现, 亦说明人类对该区域潮间带的影响深刻, 短时间内并无好转的迹象。在可预见的未来,烟台的潮间带生态系统仍将受大量人类活动的影响,比如海参养殖、贝类滩涂等。
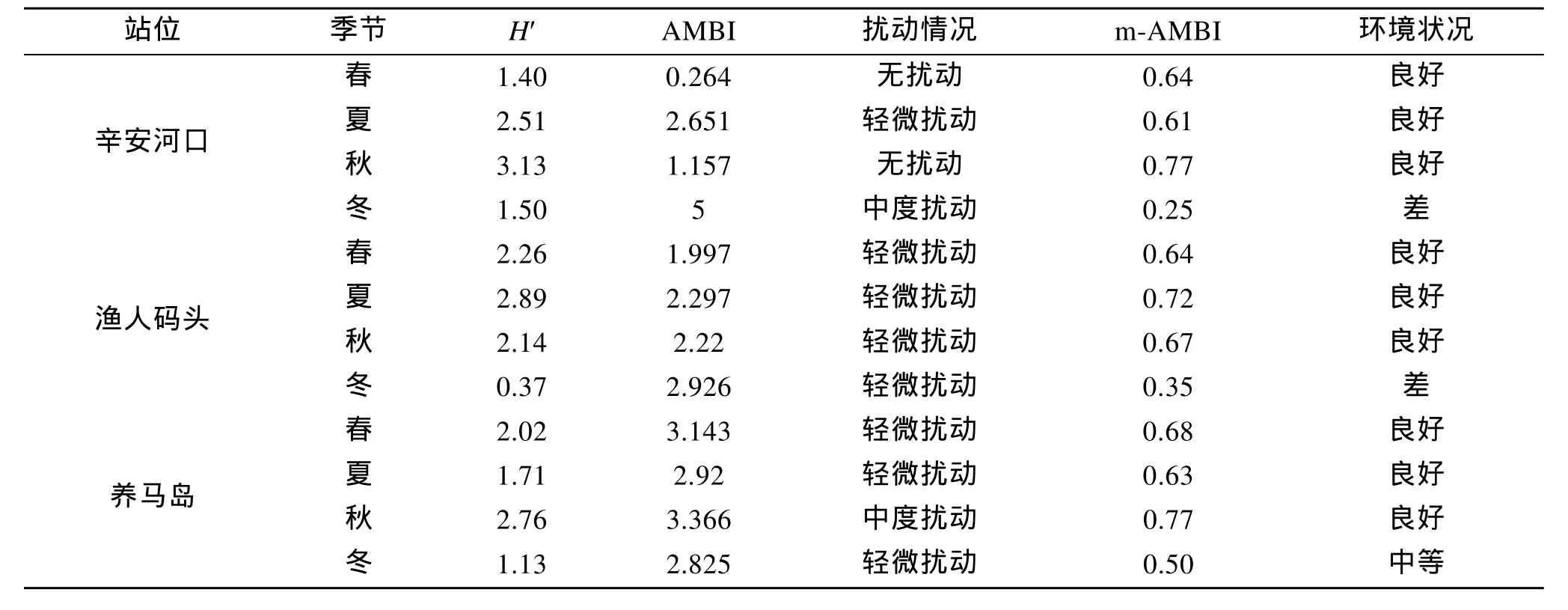
表5 烟台大型底栖动物群落的多样性、AMBI和m-AMBI生态环境评估Tab.5 Evaluation of ecological status of intertidal zones in Yantai by diversity, AMBI and m-AMBI
养马岛南岸泥沙滩在 20世纪30年代的代表动物[21]包括海仙人掌 Cavernularia habereri (Moroff.),叶须虫 Phyllodoce, 吻沙蚕 Glycera, 海豆芽Lingula anatine (Bruguiére), 钩虾 Gammarus, 虾蛄 Squilla,脊腹褐虾 Crangon affinis (de Haan)(现为日本褐虾Crangon hakodatei Rathbun), 圆球股窗蟹Scopimera globosa (de Haan), 关公蟹 Dorippe, 软体动物的Tapes varietatus (Hanley) [现为 Venerupis aspera(Quoy & Gaimard, 1835)], 扇贝Pecten laetus (Gould)[应为踵扇贝Talochlamys zelandiae (Gray, 1843)], 海螂 Mya, 蚶 Arca, 壳蛞蝓 Philine japonica Doridium(现为东方壳蛞蝓Philine orientalis A. Adams, 1854),金氏真蛇尾 Ophiura kinbergi (Ljungman), 紫蛇尾Ophiopholis mirabilis (Duncan), 哈氏刻肋海胆Temnopleurus hardwickii (Gray), 细雕刻肋海胆 Opleurus toreumaticus (Klein)。
渔人码头的泥沙岸在 20世纪30年代的代表动物[21]包括燐沙蚕Chaetopterus variopedatus (Renier),沙 蠋Arenicola, 扇贝Pectinaria, 吻沙蚕Glycera, 沙蚕Nereis, 螠虫Echiurus unicinctus (von Drasche), 裸体方格星虫 Sipunculus nudus (Linné), 海豆芽 Lingula anatina (Bruguiére), Talitrus, 跳钩虾 Orchestia,短脊鼓虾Alpheus brevicristatus (de Haan), 大蝼蛄虾Gebia major (de Haan)[大蝼蛄虾Upogebia major (De Haan, 1841)], 美人虾Callianassa harmandi (Bouvier)[应为 Nihonotrypaea harmandi (Bouvier, 1901)], 管须蟹Albunea, 圆球股窗蟹Scopimera globosa (de Haan),肉球近方蟹Hemigrapsus sanguineus (de Haan), 蓝氏三强蟹 Tritodynamia rathbuni (Shen)(现为蓝氏三强蟹Tritodynamia rathbunae Shen, 1932), 三疣梭子蟹Portunus trituberculatus (Miers), 宽身大眼蟹 Macrophthalmus dilatatus (de Haan), Uca; 圆滨蛤Hiatula olivacea (Jay)(现为圆滨蛤Nuttallia ezonis Kuroda &Habe in Habe, 1955), 日本镜蛤 Dosinia japonica(Reeve), 樱蛤 Tellina, 青蛤 Cyclina chinensis (Chemnitz), 蛤蜊Trigonella(现为Mactra Linnaeus, 1767),砂海螂Mya acuta (Say)(应为Mya arenaria Linnaeus,1758), 长竹蛏 Solen gouldi (Conrad.)(现为 Solen strictus Gould, 1861), 蛤 仔 Tapes variegates(Hanley)[现为 Venerupis aspera (Quoy & Gaimard,1835)], 织纹螺Nassa(现为织纹螺Nassarius Duméril,1805), 笋螺Terebra, 彩虹虫昌螺Umbonium vestiarium (L.), 壳蛞蝓Philine japonica (Lische)(现为东方壳蛞蝓 Philine orientalis A. Adams, 1854), 扁玉螺Natica didyma var. bicolor (Phil.) [现为Neverita didyma(Röding, 1798)], 尻参 Caudina chilensis (Müller) [现为海棒槌 Paracaudina chilensis (J. Müller, 1850)],细锚参 Leptosynapta, 心形海胆 Echinocardium cordatum (Pennant)。
以上两个潮间带区域在20世纪30年代的这些代表性种类之中, 很多体型较大的甲壳动物和软体动物均具有一定的经济价值, 如甲壳动物的虾蛄、日本褐虾、短脊鼓虾、三疣梭子蟹、大蝼蛄虾、短脊鼓虾; 软体动物的扇贝、蚶、长竹蛏, 蛤仔, 蛤蜊, 青蛤, 扇贝等等。但在2009~2010年调查中所获得的典型代表动物及优势动物中, 大型的种类退化严重,经济种类很少; 并且养马岛区泥质潮间带的经济动物菲律宾蛤仔亦与附近贝类滩涂养殖的养殖生物逃逸有关。另外, 由于在张玺[21]和张修吉[22]所记录的很多底栖种类包括了相当一部分的浅海底栖生物,绝大部分并非潮间带区域所生存, 而是生活于潮下带近岸水域, 这与本次调查的范围有所区别。
在群落丰度组成中, 日本大螯蜚是渔人码头泥沙滩的全年优势种, 作为该区域底栖群落的重要组成部分, 它的出现应与渔人码头潮间带残存的一片海草床(Zostera marina, 约 4 m2)有关, 它为这些小型端足类的出现提供了一定的食物来源及庇护场所;中型阿曼吉虫是辛安河口大型底栖动物群落的主要优势物种, 它与辛安河带至河口区域的大量有机质堆积污染有关; 而养马岛菲律宾蛤仔则与附近的滩涂养殖有关, 这些养殖贝类的逃逸对周围环境的大型底栖群落组成和结构也产生的一定的影响。这些场所均存在一定的环境污染和人为扰动, 但相对较好地保存了各自潮间带的原有特征。
国内学者广泛将大型底栖群落的多样性指数(包括香农指数、丰富度指数、均匀度指数等)与环境扰动或污染相联系, 分析环境扰动或污染对生态系统和生物群落的影响。在大型底栖群落分析方法上, 最常采用的是ABC曲线法[25-27], 以及对ABC方法进行发展改进的MPI指数法[28]。ABC曲线法在用于评价大型底栖群落和环境质量中存在一些局限性, 比如它只适用于软底质的有机污染, 并且非多毛类的小个体种和喜有机质的大个体种的存在会严重影响分析结果[29]。而采用底栖生物指数(如 AMBI和m-AMBI), 能够简化测量工作, 评估生态风险, 具有较大的尺度和环境状况上的通用性, 较为准确地预测环境变化的方向, 甚至能够确定需要采取措施的阈值所对应的环境胁迫压力。生态指数的应用较为成熟, 但生态指数的适应性与底质环境密切相关,生态指数中的丰度和均匀度等易受高运动性常见物种的影响[30], 单纯使用生态指数分析底栖群落将不够全面, 因此, 在进行群落结构和组成的分析上, 将以生态指数为主, 以主要优势物种的扰动敏感程度作为辅助参考。
在欧盟水框架计划 WFD(Water Frame Directive的推动下, 底栖生物指数得到了迅速发展, 并广泛应用于河口和海岸带水域的水环境质量检测和生态研究中, 应用比较广泛的底栖生物指数包括Borja等[31]创立的 AMBI指数 (AZTI Marine Biotic Index)、Simboura[32]的 Bentix 指数、Rosenberg 等[33]的BQI(Benthic Quality Index)、Dauvin 等[34]的 BOPA(Benthic pportunistic Polychaetes Amphipods)和Muxika等[35]以AMBI指数为基础, 整合种类丰富度和香农指数, 发展出和m-AMBI指数。AMBI指数和m-AMBI指数以Pearson-Rosenberg的有机质富集模型为理论基础[6], 将底栖物种按污染(主要是有机质污染)耐受程度, 分为 5个生态群组[31], 通过将所得的底栖种类通过分组之后, 计算生物指数[34]。AMBI和 m-AMBI指数可以反应软底质底栖群落对自然或人为扰动的响应, 被用于评价欧洲海岸的生态质量,被成功的用于包括欧洲周边各海域[36]、印度洋[37]、南美和香港[15]的水域。需要注意的是, AMBI等生物指数尽管在欧洲海域有很好的适应性[36], 但在我国北方海域使用时, 仍需要结合底栖动物区系组成、对环境改变和污染的响应特征等进行必要的修订[16]。在本文 ABC曲线和生物指数的联合应用中, AMBI和 m-AMBI指数显示出了较强的适应性, 是替代ABC曲线的较好选择。已有的研究也证实, AMBI和m-AMBI指数均能有效地检测生态系统所承受人为或环境扰动影响, 并已应用于渤海湾潮间带, 长江口, 香港和深圳等地的近海海域的底栖生态环境质量评价, 并且对于有机污染物、重金属污染、富营养化等环境压力梯度表现出较好的适应性[20,38]。
[1] 庄树宏, 陈礼学, 孙力. 南长山岛岩岸潮间带底栖藻类群落结构的季节变化格局 [J]. 海洋科学进展,2003, 21(2): 194-202.
[2] Reise K. Tidal flat ecology – an experimental approach to species interactions [M]. Berlin: Springer Verlag,1985.
[3] Boehs G, Absher T M, Cruz-Kaled A D. Composition and distribution of benthic molluscs on intertidal flats of Paranaguá Bay (Paraná, Brazil) [J]. Scientia Marina,2004, 68(4): 537-543.
[4] Herman P M J, Middelburg J J, van de Koppel J, et al.Ecology of estuarine macrobenthos [M]. Advances in Ecological Research, 1999, 29: 195-240.
[5] 沈国英, 施并章. 海洋生态学(第二版) [M]. 北京:科学出版社, 2002: 1-446.
[6] Pearson T H, Rosenberg R. Macrobenthic succession in relation to organic enrichment and pollution of the marine environment [J]. Oceanographic Marine Biology Annual Review, 1978, 16: 229-311.
[7] Ysebaert T, Herman P M J. Spatial and temporal variation in benthic macrofauna and relationships with environmental variables in an estuarine, intertidal soft-sediment environment [J]. Marine Ecology Progress Series, 2002, 244: 105-124.
[8] Elias R, Bremec C S. Biomonitoring of water quality using benthic communities in Blanca Bay (Argentina)[J]. Science of Total Environment, 1994, 158: 45-49.
[9] Gesteira J L G, Dauvin J C, Fraga M S. Taxonomic level for assessing oil spill effects on soft-bottom sublittoral benthic communities [J]. Marine Pollution Bulletin, 2003, 46: 562-572.
[10] Lim H S, Hong J S. An environmental impact assessment based on benthic macrofauna in Chinhae Bay, Korea [J]. Bulletin of Korean Fisheries Society,1994, 27: 659-674.
[11] Nipper M. Current approaches and future directions for contaminant-related impact assessments in coastal environments: Brazilian perspective [J]. Aquatic Ecosystem Health and Management, 2000, 3: 433-447.
[12] Shin P K S, Huang Z G, Wu R S S. An updated baseline of subtropical macrobenthic communities in Hong Kong [J]. Marine Pollution Bulletin, 2004, 49(1-2):128-135.
[13] 徐兆礼, 陈亚瞿. 东黄海秋季浮游动物优势种聚集强度与鲐 鲹渔场的关系 [J]. 生态学杂志, 1989, 8(4):13-15.
[14] Gaudêncio M J, Guerra M T, Glémarec M. Recherches biosédimentaires sur la zone maritime de l’estuaire du Tage, Portugal: données sédimentaires preliminaries[C]//Elliot M, Ducrotoy J C. Estuaries and Coasts:Spatial and Temporal Intercomparisons. Olsen and Olsen, Fredensborg, Denmark, 1991: 11-16.
[15] Borja Á, Muxika I. Guidelines for the use of AMBI(AZTI's Marine Biotic Index) in the assessment of the benthic ecological quality[J]. Marine Pollution Bulletin,2005, 50: 787-789.
[16] Pinto R, Patrício J, Baeta A, et al. Review and evaluation of estuarine biotic indices to assess benthic condition [J]. Ecological Indicators, 2009, 9(1): 1-25.
[17] Borja Á, Tunberg B G. Assessing benthic health in stressed subtropical estuaries, eastern Florida, USA using AMBI and M-AMBI [J]. Ecological Indicators,2011, 11(2): 295-303.
[18] Silva G, Costa J S, de Almeida P R, et al. Structure and dynamics of a benthic invertebrate community in an intertidal area of the Tagus estuary, western Portugal: a six year data series [J]. Hydrobiologia, 2006, 555:115-128.
[19] Borja Á, Muxika I. Guidelines for the use of AMBI(AZTI's marine biotic index) in the assessment of the benthic ecological quality [J].Marine Pollution Bulletin, 2005, 50(7): 787-789.
[20] 蔡文倩, 孟伟, 刘录三, 等. 长江口海域底栖生态环境质量评价—AMBI和 M-AMBI法 [J]. 环境科学,2013, 34(5): 1725-1734.
[21] 张玺. 烟台海滨动物之分布 [J]. 国立北平研究院院务汇报, 1934, 5(5): 93-158.
[22] 张修吉. 渤海海洋生物研究室第二次年报 [J]. 国立北平研究院动物学研究所中文报告汇刊, 1937, 19:1-74.
[23] 杨宗岱, 吴宝铃. 烟台潮间带底栖生物生态学的初步研究 [J]. 黄渤海海洋, 1988, 6(1): 31-38.
[24] 丛建国. 烟台潮间带底栖无脊椎动物群落和多样性研究 [J]. 生态学报, 1998, 18(1): 56-62.
[25] Warwick R M, Pearson T H, Ruswah y. Detection of pollution effects on marine macrobenthos: further evaluation of the species abundance/biomass method[J]. Marine Biology, 1987, 95(2): 193-200.
[26] 田胜艳, 于子山, 刘晓收, 等. 丰度/生物量比较曲线法监测大型底栖动物群落受污染扰动的研究 [J]. 海洋通报, 2006, 25(1): 92-96.
[27] 张敬怀, 高阳, 方宏达, 等. 珠江口大型底栖生物群落生态特征[J]. 生态学报, 2009, 29(6): 2989-2999.
[28] 蔡立哲. 大型底栖动物污染指数(MPI) [J]. 环境科学学报, 2003, 23(5): 625-629.
[29] 曲方圆, 于子山, 隋吉星, 等. 丰度生物量比较法应用局限性 [J]. 海洋科学, 2009, 33(6): 118-121.
[30] Patrício J, Neto J M, Teixeira H, et al. The robustness of ecological indicators to detect long-term changes in the macrobenthos of estuarine systems[J]. Marine Environmental Research, 2009, 68(1): 25-36.
[31] Borja Á, Franco J, Pérez V. A Marine Biotic Index to Establish the ecological quality of soft-bottom benthos within european estuarine and coastal environments[J].Marine Pollution Bulletin, 2000, 40(12): 1100-1114.
[32] Simboura N, Zenetos A. Benthic indicators to use in ecological quality classification of mediterranean soft bottom marine ecosystems, including a new biotic index [J]. Mediterranean Marine Science, 2002, 3(2):77-111.
[33] Rosenberg R, Blomqvist M, Nilsson H C, et al. Marine quality assessment by use of benthic species-abundance distributions: a proposed new protocol within the European Water Framework Directive[J]. Marine Pollution Bulletin, 2004, 49: 728-739.
[34] Dauvin J C, Ruellet T. Polychaete/amphipod ratio revisited[J]. Marine Pollution Bulletin, 2007, 55(1-6):215-224.
[35] Muxika I, Borja Á, Bald J. Using historical data, expert judgement and multivariate analysis in assessing reference conditions and benthic ecological status,according to the European Water Framework Directive[J]. Marine Pollution Bulletin, 2007, 55(1-6): 16-29.
[36] Muxika I, Borja Á, Bonne W. The suitability of the marine biotic index (AMBI) to new impact sources along European coasts [J]. Ecological Indicators, 2005,5(1): 19-31.
[37] Bigot L, Gréare A, Amouroux J M, et al. Assessment of the ecological quality status of soft-bottoms in Reunion Island (tropical Southwest Indian Ocean) using AZTI marine biotic indices [J]. Marine Pollution Bulletin,2008, 56(4): 704-722.
[38] 蔡文倩, 刘录三, 孟伟, 等. AMBI方法评价环渤海潮间带底栖生态质量的适用性[J].环境科学学报, 2012,32(4): 992-1000.
猜你喜欢
百科探秘·海底世界(2024年2期)2024-02-27 14:37:15
烟台果树(2021年2期)2021-07-21 07:18:22
邯郸职业技术学院学报(2021年4期)2021-07-20 00:40:04
海洋信息技术与应用(2020年4期)2021-01-18 06:21:36
海洋通报(2020年3期)2020-10-20 09:26:36
艺术品鉴(2020年6期)2020-08-11 09:36:24
汽车维修与保养(2019年4期)2019-09-11 05:48:48
小学阅读指南·低年级版(2019年6期)2019-09-09 00:48:23
走向世界(2019年18期)2019-08-16 13:06:02
小星星·阅读100分(低年级)(2018年7期)2018-09-04 03:16:36
