
乌金山植物群落物种多样性的垂直分布格局
2014-11-10 10:47:12牛常青牛霞霞
晋中学院学报 2014年3期
牛常青,曲 波,牛霞霞
(1.晋中学院生物科学与技术学院,山西晋中 030600;2.内蒙古大学生命科学学院,内蒙古呼和浩特 010021)
物种多样性沿环境梯度的变化规律一直是生态学家十分关注的问题.物种多样性不仅能反映群落或生境中物种的丰富度、均匀度和时空变化,表征群落和生态系统的特征及其变化演替的规律,也能反映出不同自然地理条件及人为因素与相关群落的相互关系[1].本文调查乌金山的群落类型、结构及其物种多样性垂直分布格局,运用多样性指数、丰富度指数对乌金山森林群落及林下灌木层和草本层及其灌丛草本群落的物种多样性的分析[2~5].研究物种多样性的梯度格局及控制这些格局的生态因子,是保护生物学研究的基础,其主要与气候、群落生产力和其他因子相关[6~9].调查分析乌金山植物群落的多样性对阐明森林生态系统的形成与维持群落的稳定性与演替规律、种群的生态特征和更新具有极为重要的意义,可为乌金山地区森林景观多样性和植物资源保护提供科学依据.
1 研究地区与研究方法
1.1 研究地区概况
研究区位于乌金山自然保护区,属太行山脉,地理坐标112°46′24″E,37°51′36″N,属暖温带半干旱气候,年均气温为9.8℃.最高海拔为1489.2 m,最低海拔为954.1 m.土壤为砂页岩和砂壤或轻壤.有植物330余种.
1.2 研究方法
采用梯度格局法,海拔每升高50m设置一块样地,共11个.乔木层样方大小为10m×10m;在每个乔木层样方中,选取两个5m×5m的小样方作为灌木层样方,进行灌木调查﹔在每个乔木层样方中梅花取样,设置5个1m×1m草本层样方,对草本层进行调查[10].
1.3 调查内容
(1)样地的基本状况:经纬度、海拔、坡度、坡向等;(2)乔木层中,对胸径>4 cm的所有个体,准确测定其胸径和树高;在灌木层和草本层的调查中,记录每个物种的名称、高度和盖度.
1.3.1 物种多样性测度方法[11]
物种丰富度指数=S;S=样方内所有物种数
Gleason丰富度指数(dm):dm=(S-1)/lnA;A为取样面积
1.3.2 α多样性测度方法
Shannon-Wiener指数(H):H=-∑PilnPi
Pielou均匀度指数(J):J=H/lnS
式中:Pi为样方第i种的重要值;
物种重要值(Pi)的计算:
乔木重要值=(相对盖度+相对频度+相对优势度)/300
灌木植物的重要值=(相对高度+相对盖度)/200
草本植物的重要值=(相对高度+相对盖度)/200
(相对高度就是某一物种高度除以其所在层片的总高度)
1.3.4 β多样性测度方法[5]
β多样性可以定义为沿着环境梯度的变化物种替代的程度.不同群落或某环境梯度上不同点之间的共有种越少,β多样性越大.精确地测定β多样性具有重要的意义.
Jaccard指数CJ=j/(a+b-j)
Cody指数βc=[g(H)+l(H)]/2=(a+b-2j)/2
式中:a、b分别为两群落的物种数,j为两群落的共有物种数,g(H)为沿生境梯度H增加的物种数,l(H)为沿生境梯度H失去的物种数.
2 结果与分析
2.1 乌金山森林群落种类组成分析
由表1可以看出,乌金山森林群落物种丰富,层次分明,成层现象突出,具有明显的垂直结构,分为乔木层、灌木层和草本层.
乔木层有乔木13种,其中油松存在于每个海拔梯度层,分布均匀,数量较多,由表1可以看出油松为乔木层的优势种,对乌金山森林群落的结构和环境的形成有明显的控制作用.灌木层约有16种,其中三亚绣线菊、荆条、黄蔷薇在灌木层中占优势,在一定程度上影响着群落的性质及控制着群落的环境,在中海拔出现较多物种.草本层大约有31种.

表1 乌金山植物群落物种组成

表1 乌金山植物群落物种组成(续表)
2.2 物种丰富度与海拔的关系
物种丰富度(species richness)随海拔梯度的变化格局一般有两种:物种丰富度随海拔升高而递减和中海拔地区最丰富.乌金山植物物种丰富度属于在中海拔地区较丰富,中海拔地区降水和温度条件适宜大多数物种的生存,中海拔地区生境较均匀,出现较多种类的草本与灌木,致使植物群落的多样性增加.
从表2,结合图1,2可见,物种丰富度成明显的单峰分布格局,木本植物的丰富度随海拔的升高而呈现递减趋势,而草本植物的丰富度对海拔的升高先增加后下降.物种丰富度在1250m海拔处呈现最高值.中海拔地区降水和温度条件适宜大多数物种的生存,每100m2的样方内一般有17种木本植物以及14种草本植物.
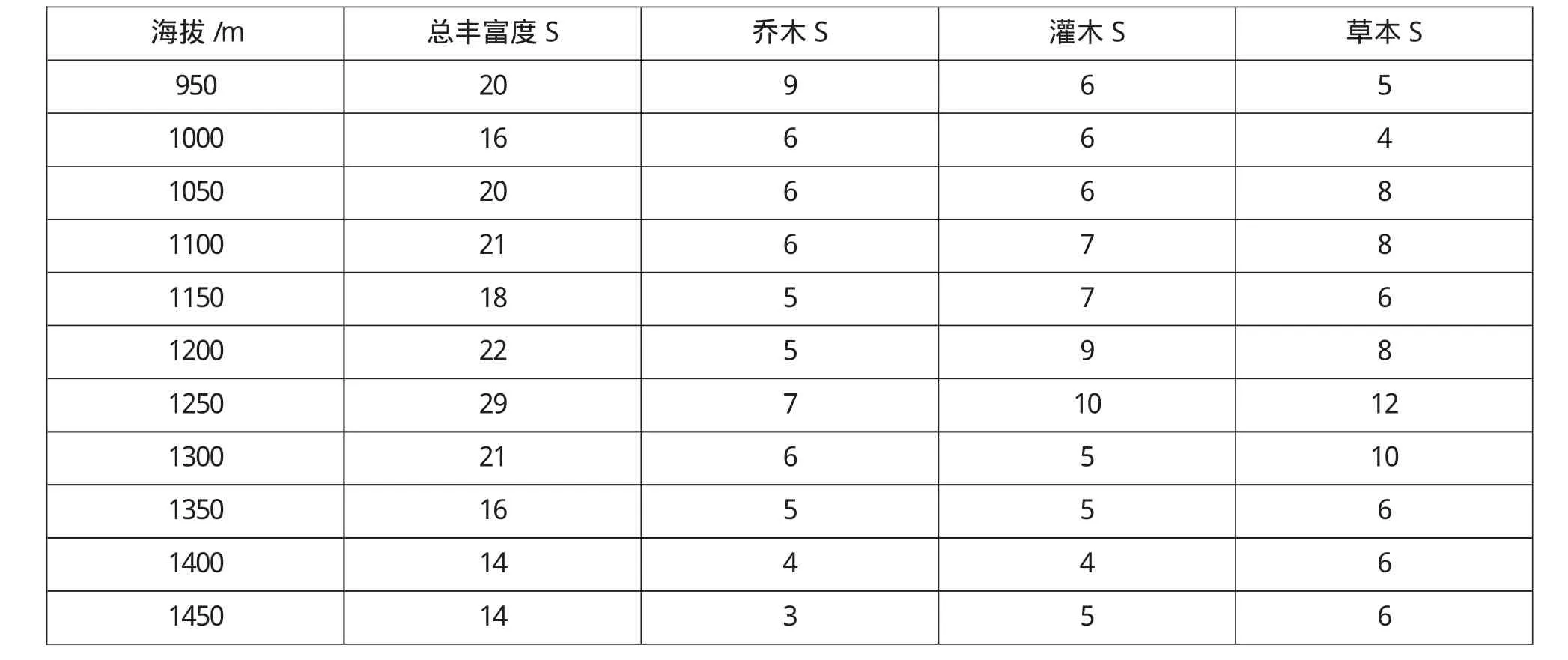
表2 乌金山植物群落物种丰富度随海拔的变化

图1 乌金山植物群落物种丰富度随海拔变化
由于不同生活型的植物存在不同的多样性分布格局,本文对乔灌草的丰富度也做了调查,如图2所示,乔木物种数在低海拔地区丰富度较高,随着海拔的升高出现下降趋势,而灌木物种数先增加后减少,因此出现木本植物的物种多样性随着海拔高度的增加而下降.而草本植物随海拔的升高先增加后减少,在中海拔出现较多的草本植物,这与中海拔地区的降水、生境比较均匀,适宜多种物种生存有关.由图2可以看出,乌金山植物群落物种多样性对海拔的敏感性由大到小的次序依次为草本层、灌木层、乔木层.
2.3 群落结构特征
本文群落的结构特征主要通过最大树高、最大胸径、平均树高、平均胸径等4个指标进行分析[8].由表3,图3中可以看出,随着海拔的升高,最大树高(Hmax)、最大胸径(DBHmax)、平均树高和平均胸径等指标总体上呈单峰分布格局.最大树高和最大胸径均在1200m~1300m呈现最高分布.平均胸径与平均树高单峰分布也比较明显.不同生活型的结构特征反应群落的潜在生产力随海拔而变化.

图2 乔灌草本植物物种丰富度随海拔的变化
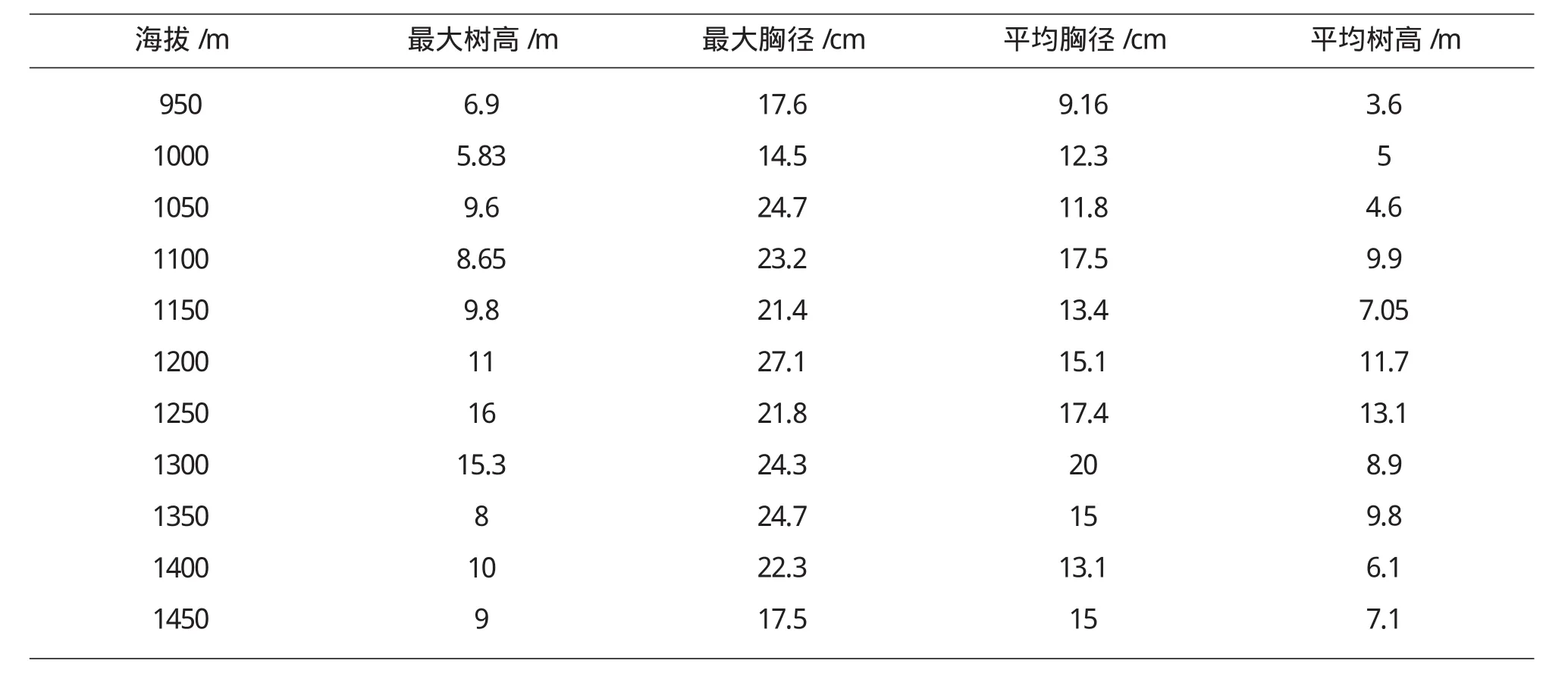
表3 乌金山植物群落结构特征随海拔的变化格局
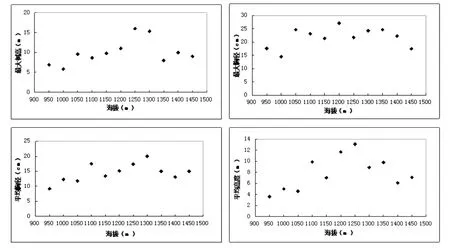
图3 乌金山植物群落结构特征随海拔的变化格局
2.4 α,β多样性随海拔的变化格局
α多样性主要关注局域均质生境下的物种数目,因此被称为生境内的多样性.一般受到小环境以及生物之间的相互作用的影响.本文对α多样性是通过Gleason丰富度指数,Shannon-Wiener指数,Pielou均匀度指数来分析的.
从表4可以看出Shannon-Wiener指数在海拔1250m处达到最大值,结合图1和图4,α多样性随着海拔梯度的变化与物种丰富度相似,随着海拔的升高先增加后降低.这与乌金山的气候条件密切相关,乌金山属于半干旱地区.在低海拔区域,物种多样性受降水的影响,而在高海拔区域受热量的限制,从而形成物种多样性在中海拔出现峰值.
β多样性指沿环境梯度不同生境群落之间物种组成的相异性或物种沿环境梯度的更替速率,即生境间的多样性.一般β多样性随海拔的升高而有所降低,在低海拔处较高,主要原因与干扰及物种之间相互竞争有关.本文是通过Jaccard指数、cody指数来分析β多样性的.Jaccard指数反映群落间物种组成的相似性;Cody指数则反映群落物种组成沿环境梯度的替代速率.
从表5可以看出,Jaccard指数在低海拔地区出现最大值说明群落间物种组成的相似性较大,然后随着海拔的升高而有所降低.结合图5可以看出,在高海拔1350m~1400m出现最大值,此区域属于群落交错区,具有较高的相似性.Cody指数在中海拔区域(1200m~1250m)达到最大值,说明群落物种组成沿环境梯度的替代速率最大.群落相异性表示沿海拔梯度相邻样地间在物种组成上的差异,物种更替则体现物种的替换速率或数量.

表4 乌金山沿海拔梯度Shannon-wiener指数和均匀度指数的变化
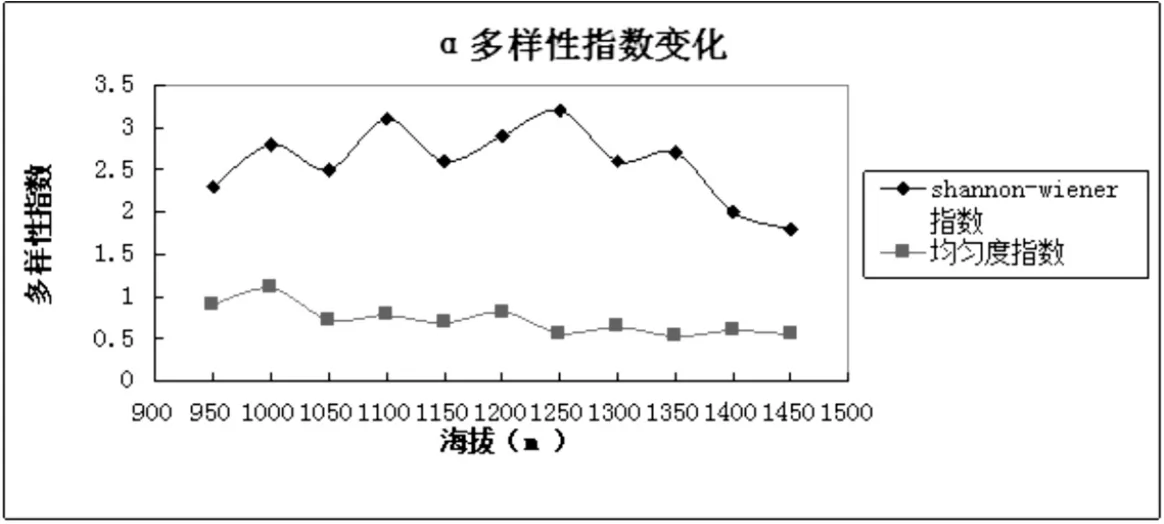
图4 乌金山沿海拔梯度Shannon-w iener指数和均匀度指数的变化
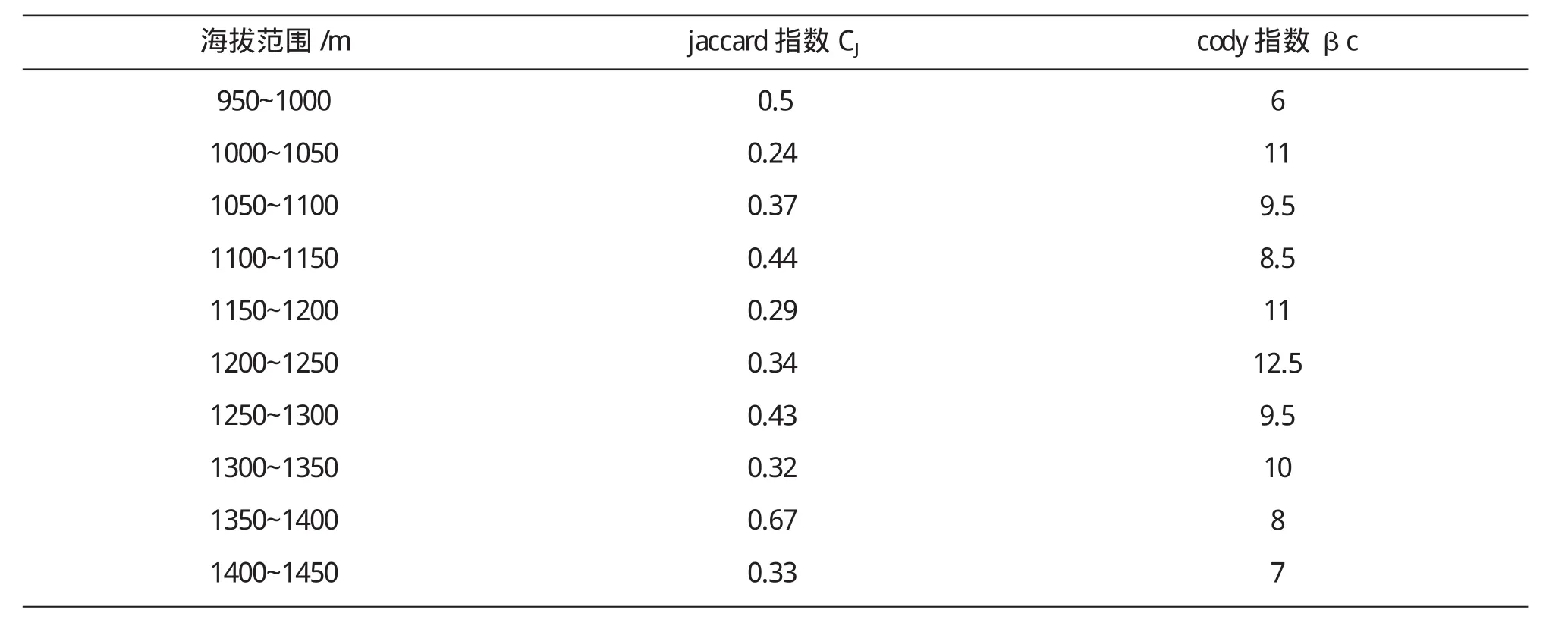
表5 乌金山沿海拔梯度相邻群落间Jaccard指数和Cody指数

图5 乌金山沿海拔梯度相邻群落间Jaccard指数和Cody指数
3 讨论
由调查可以看出乌金山植物群落物种种类丰富,垂直结构明显,共调查到乔木层有13种,灌木层有16种,草本层大约有31种.随着海拔的升高,森林群落依次为油松林、侧柏林、油松侧柏混交林、油松山桃山杏混交林、白皮松林、油松白皮松混交林.水分和能量对植物物种多样性有重要的影响.在中海拔群落(1200m~1250m)物种丰富度出现最大的值,在中海拔群落,水分、能量比较充足,适宜大多数物种生存,出现较多种类的草本与灌木,致使植物群落的多样性增加.另外也与人为因素有关.在低海拔处人为活动频繁,有损生物多样性,在低海拔区域的物种多样性受降水的影响,而在高海拔区域受热量的限制,从而形成物种多样性在中海拔出现峰值.木本植物的物种丰富度随着海拔高度的增加而下降,草本植物的物种丰富度随着海拔高度的增加先增加后下降.乌金山植物群落物种多样性对海拔的敏感性由大到小的次序依次为草本层、灌木层、乔木层.这与张峰、张金屯、上官铁梁对历山自然保护区猪尾沟森林群落植物多样性研究所得结论有所差异,主要原因是乌金山与历山所处地理位置不同、环境因子不同、生态气候不同,从而使不同生活型的物种多样性出现了差异.
α多样性一般受到小环境和生物之间的相互作用的影响.α多样性随着海拔梯度的变化与物种丰富度相似,随着海拔的升高先增加后降低.这与乌金山的气候条件密切相关,乌金山属于半干旱地区.在低海拔区域,物种多样性受降水的影响,而在高海拔区域受热量的限制,从而形成物种多样性在中海拔出现峰值.α多样性主要与水热条件有关,符合唐志尧、方精云的植物物种多样性的垂直分布格局研究的α多样性一般格局.影响β多样性的主要因子有土壤、地质以及干扰等,β多样性随海拔的升高而有所降低,在低海拔较高,主要原因与干扰及物种之间相互竞争有关,低海拔生境异质性较大,在土壤和地形变化频繁的地段,群落含更多植物种.因为海拔不同出现不同的群落,在群落交错的地段,物种多样性较大.
[1]王国宏.祁连山北坡中段植物群落多样性的垂直分布格局[J].生物多性,2002(10):7~14.
[2]高贤明,陈灵芝.北京山区辽东栎群落物种多样性的研究[J].植物生态学报,1998(22):23~32.
[3]唐志尧,方精云.植物物种多样性的垂直分布格局[J].生物多样性,2004,12(1):20~28.
[4]曾从盛.福建典型区生态环境研究[M].北京:中国环境科学出版社,2006.
[5]朱彪,陈安平,刘增力,等.广西猫儿山植物群落物种组成、群落结构及树种多样性的垂直分布格局[J].生物多样性,2004,12(1):44~52.
[6]徐东成,冯建孟,王襄平,等.云南高黎贡山北段植物物种多样性的垂直分布格局[J].生态学杂志,2008,27(3):323~327.
[7]赵淑清,方精云,朴世龙,等.大兴安岭呼中地区白卡鲁山植物群落结构及其多样性研究[J].生物多样性,2004,12(1):182~189.
[8]冯建孟,王襄平,徐成东,等.玉龙雪山植物物种多样性和群落结构沿海拔梯度的分布格局[J].山地学报,2006,24(1):110~116.
[9]唐志尧,柯金虎.秦岭牛背梁植物物种多样性垂直分布格局[J].生物多样性,2004,12(1):108~114.
[10]方精云,王襄平,沈泽昊,等.植物群落清查的主要内容、方法和技术规范[J].生物多样性,2009,17(6):533~548.
[11]李智琦,欧阳志云,曾慧卿.基于物种的大尺度生物多样性热点研究方法[J].生态学报,2010,30(6):1586~1593.
猜你喜欢
中华建设(2024年3期)2024-03-16 08:04:52
中华建设(2023年10期)2023-10-07 05:23:04
建筑与预算(2022年6期)2022-07-08 13:59:18
VOGUE服饰与美容(2022年5期)2022-05-01 10:38:53
小猕猴学习画刊·下半月(2022年2期)2022-04-16 16:06:33
——探访煤炭博物馆
奇妙博物馆(2022年3期)2022-03-23 05:16:20
少儿美术(2020年8期)2020-11-06 05:21:38
中成药(2018年12期)2018-12-29 12:25:34
小说月刊(2014年1期)2014-04-23 08:59:59
建筑与预算(2014年5期)2014-04-11 07:29:28
