
白纹伊蚊气味受体基因OR21a的克隆及定量分析*
2014-11-10 02:00李海滔赵明慧苏建新董言德李春晓赵彤言
寄生虫与医学昆虫学报 2014年2期
阎 婷 李海滔 赵明慧 苏建新 董言德 李春晓 赵彤言
(军事医学科学院微生物流行病研究所,病原微生物生物安全国家重点实验室, 北京 100071)
白纹伊蚊Aedesalbopictus,嗜吸人血,是我国登革热和登革出血热的重要传播媒介(安继尧等,2003)。由于白纹伊蚊的孳生特点,环境治理和化学防治常常难以控制蚊虫密度,近年来,研究人员试图从白纹伊蚊的生理行为方面寻找突破口,为发展新的防治方法奠定基础。
蚊虫的很多行为,例如产卵、搜寻定位宿主、吸血等都依赖于嗅觉系统来完成(Takkenetal.,1999)。蚊虫通过嗅觉系统识别环境中的气味分子,将化学信息编码成电信号,传递给脑部,从而引起行为反应(李慧等,2010)。在蚊虫嗅觉系统中,主要包括气味结合蛋白(Odorant binding protein,OBP)、气味受体(Odorant receptor,OR)等多种嗅觉蛋白,其中气味受体蛋白具有结合气味分子的特性,已成为筛选蚊虫驱避剂或引诱剂的重要靶标基因(徐炜,2009;李慧等,2010)。
二氧化碳(CO2)是人和动物宿主呼出的主要气体成分,也是蚊虫搜寻定位宿主的重要信息来源(Guerensteinetal., 2008; Hallemetal., 2008),目前基于该原理的CO2诱蚊灯等设备已普遍应用于蚊虫种群密度调查等试验,有研究证实CO2在一定的释放范围内,随浓度升高,捕获的蚊种及数量均有增加(Klineetal., 1990; Klineetal., 1991)。对黑腹果蝇(Joneetal., 2006)的研究发现,当受体Gr21a和Gr63a共同表达时表现出对CO2的敏感性,二者缺一不可。对冈比亚按蚊Anophelesgambiae(Hilletal., 2002)和致倦库蚊Culexquinquefasciatus(Turneretal., 2009)的研究发现,与果蝇Gr21a和Gr63a同源的气味受体基因,具有相似的表达方式,很可能也具有相同的功能(Hilletal., 2002)。如果有针对地研究与CO2相关的气味受体,探索其分子机制,进而干扰由CO2调节的蚊虫搜寻宿主的行为反应,将会大大降低蚊虫吸血及传播疾病的概率。
本研究通过同源扩增的方法,获得白纹伊蚊OR21a基因片段,应用定量PCR技术比较未吸血雌蚊、打断吸血雌蚊、饱血雌蚊和雄蚊头部中OR21a基因起始表达量的差异,为推断OR21a基因的功能和进一步阐明白纹伊蚊嗅觉感受系统的分子机制奠定基础。
1 材料与方法
1.1 实验用虫
白纹伊蚊敏感株来源于军事医学科学院微生物流行病研究所媒介生物学和防治研究室,蚊虫养殖条件为:温度(26±1)℃、相对湿度65%±5%、光照∶黑暗=14 h∶10 h,成蚊喂食8%糖水,羽化3~5 d的蚊虫用于实验。
1.2 白蚊伊蚊OR21a片段的克隆
1.2.1引物合成: 根据GenBank数据库中埃及伊蚊、致倦库蚊和冈比亚按蚊的OR21a基因序列,使用Primer premier 6.0软件,在相似度高的区域设计简并引物。上游引物OR1:5′- ATGATTCACA(G/C)(C/T/A)CAGATGGA -3′;下游引物OR2:′- GGTCA(A/G)(C/T)TTGAACTGCATCA -3′;引物由北京赛百盛生物技术有限公司合成。
1.2.2白纹伊蚊OR21a基因的扩增: 使用总RNA提取试剂盒(吉百特公司,DR202)提取白纹伊蚊5只雌蚊的总RNA,50 μL DEPC水溶解。取7 μL总RNA溶液,根据全式金公司 TransScript All-in-One First-Strand cDNA Synthesis SuperMix for qPCR(One-Step gDNA Removal)试剂盒说明书进行逆转录,得到cDNA溶液20 μL。PCR反应体系总体积50 μL,包括Premix Taq(loading dye mix) 25 μL,模板cDNA 1.0 μL, 上游引物(20 μmol/L)1.0 μL, 下游引物(20 μmol/L)1.0 μL, ddH2O 22 μL。反应条件为94 ℃, 3 min;94 ℃ 45 s,5 5℃ 45 s,72 ℃ 30 s,40个循环;72 ℃, 7 min;4 ℃保存。取25 μL PCR产物,1.2%琼脂糖凝胶电泳检测(120 V 30 min)后,对PCR产物进行回收及纯化。
1.2.3PCR产物的克隆、鉴定和测序: 将纯化的DNA片段和pMD18-T载体连接,转入感受态大肠杆菌DH5α,蓝白斑筛选后,对阳性克隆测序,将测序结果与NCBI数据库中的已知物种核酸序列进行比对。
1.3 白纹伊蚊OR21a基因的定量分析
1.3.1白纹伊蚊头部cDNA 的获得: 用手术刀切下白纹伊蚊未吸血雌蚊、雄蚊、打断吸血雌蚊和饱血雌蚊的头部,每1.5 mL冰上预冷的离心管中分别放入5只未吸血雌蚊、打断吸血雌蚊、饱血雌蚊和雄蚊头部。提取白纹伊蚊头部总RNA,逆转录后-20℃保存备用。
1.3.2设计引物: 根据简并引物扩增得到的气味受体基因测序结果,使用Primer premier 6.0 软件,设计特异引物,引物序列:上游为:5′-TCTTCACCACAATTATCGCCAC-3′,下游为:5′-GATGGCCACCAGAAACATTTC-3′。管家基因β-actin的引物序列参考文献(阎婷等,2010),引物由赛百盛生物技术有限公司合成。
1.3.3白蚊伊蚊OR21a和β-actin基因的扩增: PCR反应体系同(1.2.2),反应条件为94 ℃, 3 min;94 ℃ 45 s,60 ℃ 30 s,72 ℃ 30 s,40个循环;72 ℃, 7 min;4 ℃保存。
1.3.4PCR 产物的克隆、测序和鉴定: 分别将两条基因PCR扩增产物与pMD 18-T载体连接,蓝白斑筛选出的阳性克隆经检测后送公司测序,测序结果进行BLAST比对鉴定。
1.3.5制备OR21a和β-actin基因标准品,构建标准曲线: 按照天根公司高纯度质粒试剂盒说明书提取重组质粒,测定质粒浓度,计算拷贝数,按10倍梯度逐级稀释制备标准品,范围为103~108copies/μL。反应体系总体积25 μL,包括Power SYBR Green PCR Master 2×Mix 12.5 μL,模板cDNA 1.0 μL, 上游引物(10 μmol/L)1.0 μL, 下游引物(10 μmol/L)1.0 μL, ddH2O 9.5 μL。反应条件为50 ℃ 2 min,95 ℃ 10 min;95 ℃ 15 s,60 ℃ 1 min,进行40个循环,熔解曲线反应条件为:95 ℃ 15 s,60 ℃ 1 min,95 ℃ 15 s。
1.3.6白纹伊蚊头部OR21a和β-actin基因的定量分析: 构建标准曲线后,用β-actin基因的拷贝数对OR21a基因的拷贝数进行校正,比较OR21a基因在白纹伊蚊未吸血雌蚊、打断吸血雌蚊、饱血雌蚊和雄蚊头部表达量的差异倍数。
1.4 白蚊伊蚊OR21a基因在蚊虫不同发育阶段的分布
使用1.3.2所设计的引物,通过扩增检测OR21a基因是否在白纹伊蚊1~2龄幼虫、3~4龄幼虫、蛹和成蚊头部中分布。
2 结果
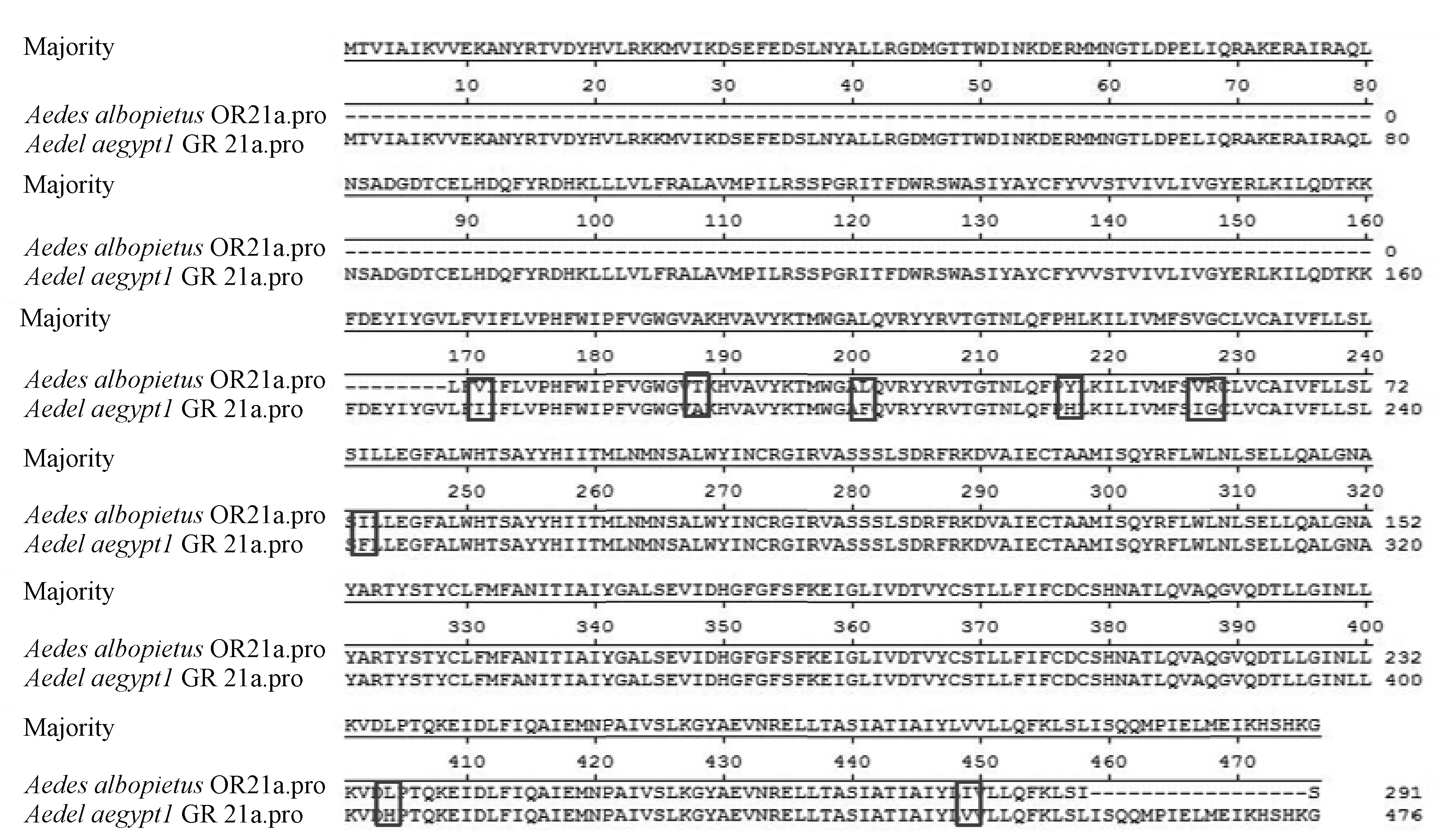
图1 白纹伊蚊OR21a和埃及伊蚊GR21a氨基酸序列及比较Fig. 1 Amino acid sequences of of AealOR21a and AedeGR21a and comparison within them方框:差异的氨基酸;Box: the difference of amino acids.
2.1 白纹伊蚊雌蚊OR21a基因的序列分析
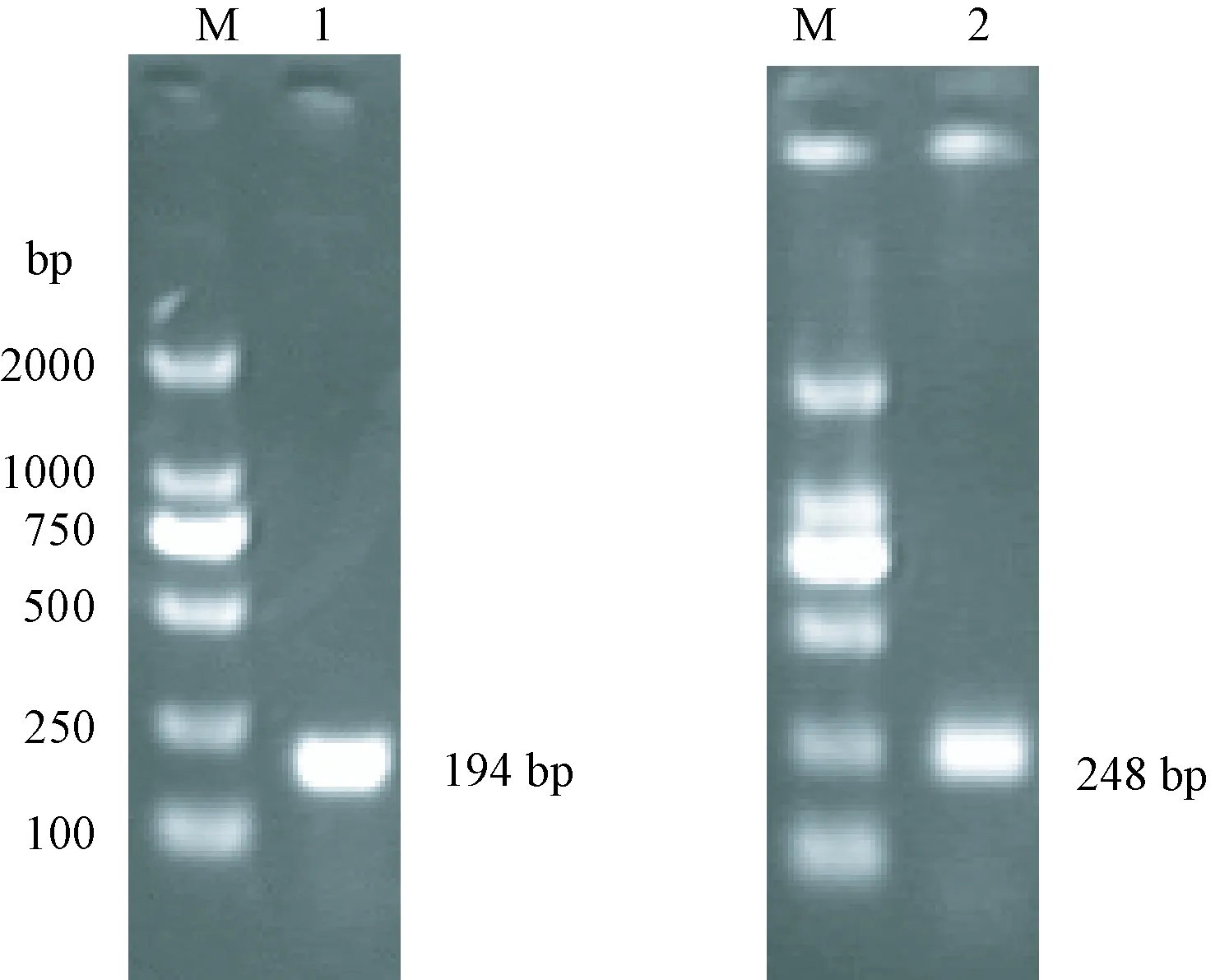
图2 白纹伊蚊基因扩增产物电泳图Fig. 2 Electrophoresis of PCR products of Ae.albopictus genes fragmentM:DL2000 marker;1:β-actin基因扩增条带;2:OR21a基因扩增条带。M: DL2000; 1: β-actin amplification band; 2: OR21a amplification band.
以白蚊伊蚊cDNA为模板,设计简并引物,经PCR扩增克隆测序后,得到白蚊伊蚊OR21a基因测序(GenBank登陆号KM056288),基因片段长度为875 bp,预测编码291个氨基酸片段。将测序结果在NCBI数据库中进行核酸比对,发现该基因与其他蚊种OR基因的相似性大于75%。与埃及伊蚊GR21a氨基酸同源性最高,大于90%(图1),将克隆得到的该基因命名为OR21a。
2.2 白纹伊蚊OR21a基因的差异表达分析
2.2.1白纹伊蚊OR21a基因与管家基因β-actin的扩增结果(图2):以白纹伊蚊未吸血雌蚊cDNA为模板,特异性引物扩增白纹伊蚊OR21a、β-actin片段大小分别为248、194 bp,将PCR产物直接送往测序公司进行测序,测序结果完全与预期结果相符。
2.2.2标准曲线和熔解曲线的制作(图3~6):白纹伊蚊OR21a和β-actin重组质粒的浓度和OD比值分别为:370 μg/mL, A260/280=1.83;350 μg/mL,A260/280=1.80。将质粒浓度换算成拷贝数后, 10倍浓度稀释用于制作标准曲线和熔解曲线,每个反应重复3次。从标准曲线斜率可知OR21a和β-actin基因的扩增效率较高,并且熔解曲线表明OR21a和β-actin基因都不存在非特异性扩增。
图3 β-actin定量PCR的标准曲线Fig.3 Standard curve of β-actin for real-time PCR
2.2.3OR21a基因表达量的定量分析结果:白纹伊蚊未吸血雌蚊、打断吸血雌蚊、饱血雌蚊和雄蚊头部中OR21a基因拷贝数进行定量分析,并比较雌蚊吸血前后和雌雄蚊头部OR基因的表达差异倍数,结果见表1和图7。
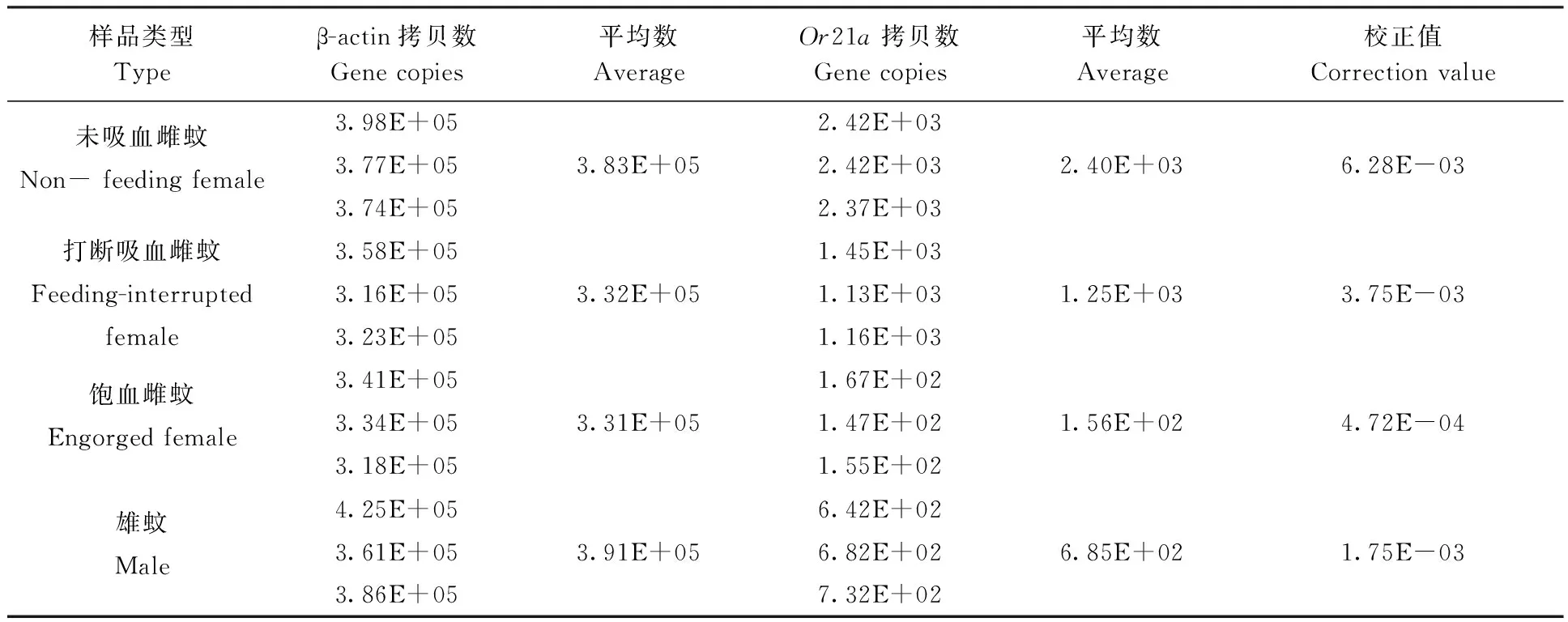
表1 OR21a基因在白蚊伊蚊未吸血雌蚊、打断吸血雌蚊、饱血雌蚊和雄蚊头部的表达量Tab. 1 The OR21a expression level in the heads of non-feeding female, feeding-interrupted females, engorged female and male of Ae. albopictus
图4 β-actin实时定量PCR熔解曲线Fig.4 Melting curve of β-actin for real-time PCR
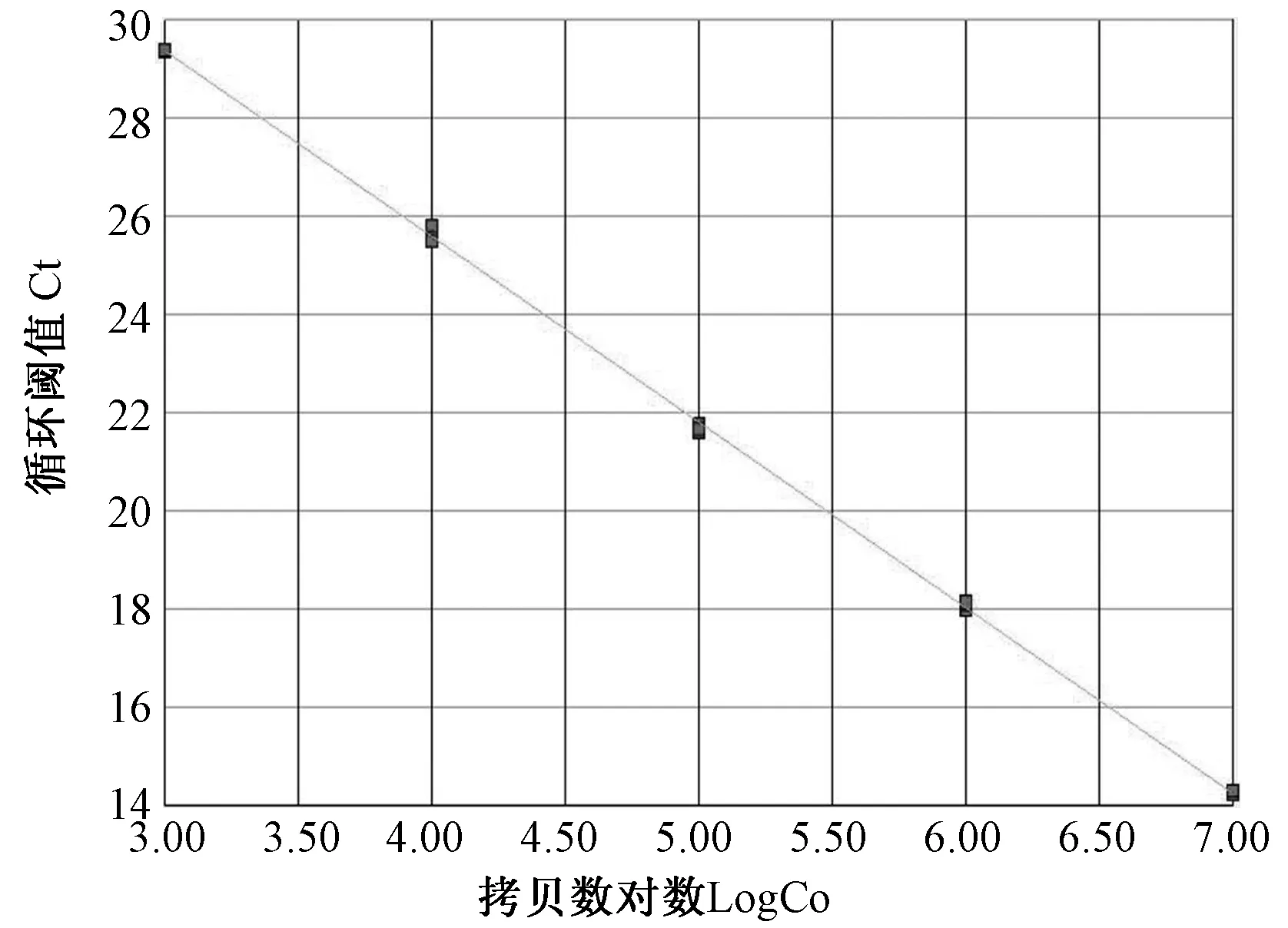
图5 OR21a实时定量PCR的标准曲线制作图Fig. 5 Standard curve of OR21a for real-time PCR

图6 OR21a实时定量PCR熔解曲线Fig. 6 Melting curve of OR21a for real-time PCR

图7 OR21a基因差异表达倍数Fig. 7 The expression ratio of OR21a genes NF: 未吸血雌蚊 Non-feeding female; BIF: 打断吸血雌蚊 Feeding-interrupted female; BF: 饱血雌蚊Engorged female; M: 雄蚊 Male.* 有显著性差异,P<0.05。* Significant differences,P<0.05.
OR21a基因在未吸血雌蚊和打断吸血雌蚊头部表达没有显著性差异,但表达量有下降的趋势;OR21a在未吸血雌蚊和雄蚊头部和未吸血雌蚊和饱血雌蚊头部之间表达都存在显著性差异。其中白纹伊蚊OR21a基因在未吸血雌蚊头部表达量最高,饱血雌蚊头部表达量最低,而在雄蚊头部的表达量介于打断吸血雌蚊和饱血雌蚊之间。
2.3 OR21a基因在蚊虫不同发育阶段的分布
从图8上可以看出,OR21a基因在白纹伊蚊1~2龄幼虫、3~4龄幼虫、蛹、成蚊头部中均有分布。
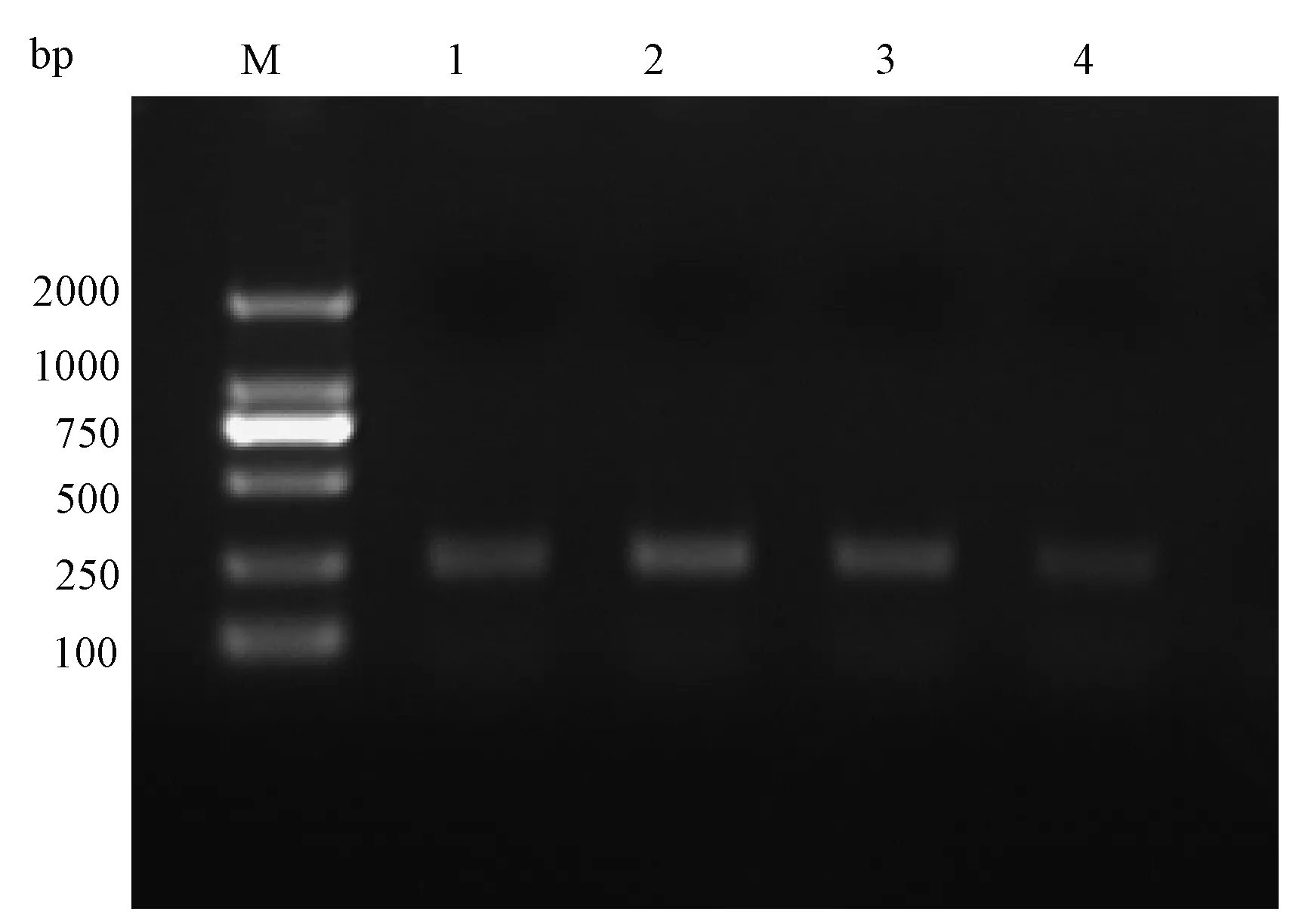
图8 白纹伊蚊OR21a在幼虫、蛹、成蚊头部的PCR结果Fig.8 Electrophoresis of PCR products of OR21a genes in the larvae, pupae, and head of adult femaleM:DL2000 marker;1: Or21a在1-2龄幼虫扩增条带;2:Or21a在3~4龄幼虫扩增条带;3:Or21a在蛹扩增条带;4:Or21a在成蚊头部扩增条带。M: DL2000; 1: OR21a amplification band of first- and second-instar larvae; 2: OR21a amplification band of third- and fourth-instar larvae; 3: OR21a amplification band of pupae; 4: OR21a amplification band of female head.
3 讨论
研究已证实蚊虫主要依靠嗅觉系统来完成定位宿主、吸血、产卵等行为,对蚊虫嗅觉系统的研究已成为发展蚊虫防治方法的新方向,但到目前为止蚊虫嗅觉具体的分子机理还知之甚少。冈比亚按蚊气味受体的功能研究发现,一些气味受体会对相应的气味物质产生较强烈的反应,同时某种气味物质也会引起一种或多种气味受体的反应,可见气味物质和气味受体之间一定存在特殊的信号传递通道来控制蚊虫的行为(Careyetal., 2010)。蚊虫气味受体数量普遍较多(Hilletal., 2002; Bohbotetal., 2007),其中已知致倦库蚊拥有个数最多,为130个(Xuetal., 2013),气味受体与其他嗅觉蛋白相互合作、共同发挥作用,可见蚊虫的嗅觉系统是一个相当敏感、精细且可调节的系统,本研究尝试有针对性地研究与感知CO2紧密相关的、可能在白蚊伊蚊搜寻宿主中起重要作用的气味受体基因,希望为蚊虫嗅觉机理的研究及治奠定基础。
分布结果表明,OR21a在白蚊伊蚊的各个生长阶段(幼虫、蛹及成蚊)均有表达,与果蝇Gr21a和Gr63a的表达方式相同(Jonesetal., 2006),目前对OR21a在蚊虫幼虫及蛹阶段表达所起的作用还需进一步的研究。对果蝇的研究发现,1-乙醇和2,3-丁二酮通过作用于Gr21a和Gr63a受体,抑制神经元对CO2气体的电生理反应,致倦库蚊则不同, 1-乙醇和1-丁醛能抑制神经元的生理反应,而2,3-丁二酮没有抑制作用(Turneretal., 2009)。可见,双翅目昆虫在长期不同的进化过程中,依循适者生存的原则,虽然属于同源基因,但分子调节机制发生了相应的改变。
本研究克隆得到白纹伊蚊嗅觉感受机制中很可能与CO2相关的气味受体基因OR21a,通过定量分析得知白蚊伊蚊OR21a在未吸血雌蚊、雄蚊、打断吸血雌蚊、饱血雌蚊头部的表达量存在差异,这说明蚊虫会根据不同的生理需求调整OR21a的表达量,实验数据表明OR21a在蚊虫头部的表达量从高至低依次为未吸血雌蚊、打断吸血雌蚊、雄蚊和饱血雌蚊,可见蚊虫吸血后OR21a表达量呈降低趋势,有可能由于吸血后蚊虫生理需求发生改变,对搜寻宿主的需求降低,进而蚊虫对环境中CO2气体的敏感度降低。在日后的研究中,可以尝试采用RNAi技术,观察白蚊伊蚊OR21a被部分或全部干扰后蚊虫的行为反应,尤其是对宿主的识别能力,进一步确认OR21a的功能,为防治虫媒病提供新的思路。
猜你喜欢
小学生学习指导(爆笑校园)(2022年9期)2022-09-30
保健医苑(2022年6期)2022-07-08
今日农业(2021年12期)2021-11-28
化工管理(2021年7期)2021-05-13
中国人兽共患病学报(2021年7期)2021-03-28
寄生虫与医学昆虫学报(2021年3期)2021-02-10
文苑(2020年7期)2020-08-12
小学科学(学生版)(2017年11期)2017-12-20
广东第二课堂·小学(2016年9期)2016-05-14
红蜻蜓·低年级(2015年11期)2016-02-02
